Abstract
Microbial communities in the plant rhizosphere – the soil region closely associated with plant roots – play critical roles in shaping plant growth, defence, fitness, and ecosystem processes. Inoculation of plants with specific rhizobacteria strains has shown promising potential for increasing crop yields. Rhizobacteria can also induce plant defences, resulting in reduced insect growth and reproduction, and can manipulate plant biochemistry to alter insect host-choice and recruit natural enemies of the insects. We present a meta-analysis examining the impact of rhizobacteria inoculation of plants on insect fitness and behaviour. Our findings indicate that rhizobacteria inoculation generally reduces herbivore fitness and affects host choice behaviours. However, effect sizes varied significantly depending on type of herbivore (chewing versus sucking), plant host, and rhizobacterial species. Bacillus spp. showed stronger effects than the commonly studied Pseudomonas spp. Rhizobacteria notably reduced traits such as host choice, leaf consumption, survival, and reproduction of chewing herbivores, while primarily impacting sucking herbivores by reducing reproduction. Single-strain inoculants tended to perform better, especially for sucking herbivores, suggesting potential strain incompatibility issues with multi-strain inoculants. Furthermore, field trials showed less impact on insect fitness reduction compared to experiments under controlled conditions, possibly due to soil diversity and environmental factors affecting inoculant persistence. Despite very limited experimental data, studies observed that inoculation with rhizobacteria can attract parasitoid wasps and predators to the plants, as an additional indirect benefit, but with variable results across the few studies. These results underscore the need for considering broader environmental interactions when developing effective rhizobacteria-based pest management strategies. Understanding specific and generalist rhizosphere interactions can aid in developing synthetic microbial communities with broad protective functions across various plants and environments.
1 Introduction
The rhizosphere is the narrow region of soil directly influenced by plant roots, hosting a dynamic and diverse microbial community with up to 1011 microbial cells per gram root and more than 30,000 prokaryotic species (Berendsen et al., 2012). This zone plays a vital role in sustaining plant health by fostering interactions between roots and beneficial microorganisms. Plants influence the rhizosphere microbiome by releasing root exudates and altering soil conditions, selectively recruiting distinct microbial communities tailored to a plant’s unique physiological and biochemical traits (Berendsen et al., 2012; Trivedi et al., 2020). Recruitment of rhizosphere bacteria can occur over short-time frames, enabling a plant to efficiently respond to changes in the environment, e.g. from drought or nutrient stress (Bakker et al., 2013). In agriculture, applying specific rhizobacterial strains to crops is promoted as a biostimulant strategy to enhance nutrient uptake and support plant growth, offering an alternative to synthetic fertilizers (Trivedi et al., 2020). Recent syntheses examining the effect of rhizobacterial inoculation on plant responses to drought (Rubin et al., 2017; Zhao et al., 2022), temperature (Zhang et al., 2023), salt (Pan et al., 2019), phosphate (De Zutter et al., 2022) and across different experimental factors (da Silva et al., 2024; Li et al., 2022; Zeffa et al., 2020) have demonstrated the strong and broad impact that different rhizobacteria can have on plant fitness and yield.
Plants can also respond to herbivory through recruitment of different rhizobacteria communities, that enhance plant defences responses, thereby decreasing insect reproduction and population growth rates (Pineda et al., 2010). Even before herbivory, plants can be primed to respond more quickly upon insect feeding through application of various plant hormones but also inoculation with defence-inducing rhizobacteria (Pieterse et al., 2013, 2014; Selosse et al., 2014). Priming of a plant can involve structural changes in the histones that fold DNA, leading to a quicker initiation of RNA transcription and faster synthesis of defence-related compounds. Application of defence inducing rhizobacteria, as single or combination inoculants, to crop plants can reduce the survival and fecundity of attacking insect pests (bioprotectants) and is a potential alternative to chemical pesticides. However, the mechanisms underlying this vary across plant and herbivore species, with variable induction of defences by different rhizobacteria. In addition to priming and induction of chemical defences, rhizobacteria may also induce changes in secondary metabolites including volatile emissions that are known to attract natural enemies of herbivores to infested plants (Bouwmeester et al., 2025). The main groups of plant-beneficial rhizobacteria studied thus far include Bacillus spp., Pseudomonas spp., Azospirillum spp. and various N-fixing rhizobia (Trivedi et al., 2020). Molecular mechanisms underlying the induction of plant defences against pathogens and pests have been most widely studied in B. subtilis (Blake et al., 2021), and P. simiae WCS417r (Pieterse et al., 2021); however, the taxonomic diversity of rhizobacteria studied is increasing over time.
Plant responses to chewing insects, e.g. caterpillars, differ from responses to sucking insects, e.g. aphids or whitefly (Schuman and Baldwin, 2016; Wu and Baldwin, 2010). The complex interplay between plant responses and the type of herbivory suggests that the effectiveness of rhizobacteria is context-dependent (Blubaugh et al., 2018; Friman et al., 2021). Chewing insects often trigger robust wound-response pathways, activating jasmonic acid signalling, whereas sucking insects primarily induce salicylic acid pathways, leading to different plant defence mechanisms (Walling, 2000). This divergence in defence strategies implies that rhizobacteria-mediated protection may be specific to the type of herbivore involved, highlighting the importance of understanding these relationships to optimize biological control strategies.
Here we present a meta-analysis to synthesise published data measuring effects of experimental bacterial inoculation of plants on insect behaviour and fitness, comparing chewing and sucking herbivorous insects. In addition to defence induction, microbial inoculation and herbivory can also trigger changes in plant volatiles that can recruit natural enemies of the herbivores. While Herbivore-Induced Plant Volatiles (HIPVs) in plants are well-known to attract natural enemies (Turlings and Erb, 2018), the consequences of microbial-induced plant volatiles on herbivore or natural enemy behaviour is relatively unstudied (Bouwmeester et al., 2025). Therefore, we also examined parasitoid wasp and predator behaviour in response to bacterial-inoculation of plants. This meta-analysis aims to describe general effects of experimental microbial inoculation of plants on insect behaviour and fitness, with a focus on rhizosphere bacteria effects on chewing and sucking insects, and their natural enemies.
Our main hypotheses are:
The effect of rhizobacteria inoculation on insect responses varies across insect feeding groups (chewing herbivores, sucking herbivores, natural enemies of herbivores).
The effect of rhizobacteria inoculation on herbivore insect responses varies across main bacterial genus, insect family and host plant family.
The type of inoculation (single or community) will impact insect responses.
The experimental environment (controlled pot experiments vs field trials) will alter the strength of response of the insect.
2 Materials and methods
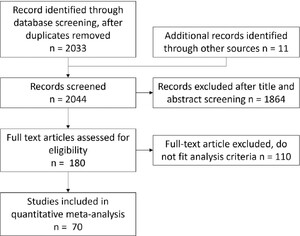
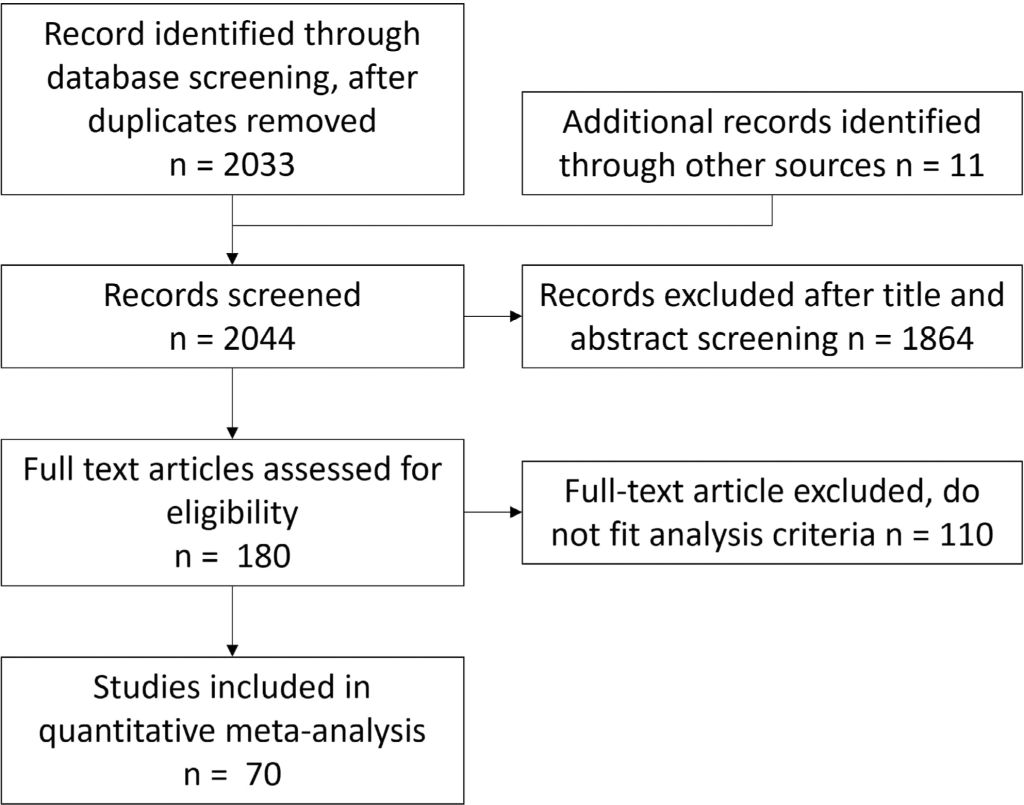
We used two broad and inclusive search approaches to maximise identification of papers for inclusion. The first search was compiled in ISI Web of Science up to September 2023 using the following keywords combinations: (Rhizo* AND aphid*), (Rhizo* AND Whitefly*), (Rhizo* AND cater*), (Rhizo* AND lepido*), (Rhizo* AND herbivor*), resulting in 428 papers. We used separate terms for aphid, whitefly, caterpillars, and lepidoptera as these are well-known species studied in this area, and the term herbivore was used to encompass a wider range. The second search used three sub-searches using terms 1. (‘Insect*’ AND (‘volatile*’ OR ‘terpen*’ OR ‘sesquiterp*’ OR ‘floral volatile*’ OR ‘leaf volatile*’) AND (‘microb*’ OR ‘bacteri*’ OR ‘rhizo*’ OR ‘fungi*’) AND (‘behav*’ OR ‘attract*’ OR ‘repel*’ OR ‘pref*’ OR ‘oviposit*’ OR ‘feed*’), 2. (‘Insect*’ OR ‘herbiv*’ OR ‘behav*’ OR ‘parsit*’) AND (‘volatile*’ OR ‘terpen*’ OR ‘sesquiterp*’ OR ‘floral volatile*’ OR ‘leaf volatile*’) AND (‘microb*’ OR ‘bacteri*’ OR ‘fungi*’) AND (‘plant*’) AND (‘behav*’ OR ‘attract*’ OR ‘repel*’ OR ‘pref*’), 3. (‘Insect*’ OR ‘herbiv*’ OR ‘behave*’ OR ‘parsit*’) AND (‘volatile*’ OR ‘terpen*’ OR ‘sesquiterp*’ OR ‘floral volatile*’ OR ‘leaf volatile*’ OR ‘secondary metabol*’) AND (‘microb*’ OR ‘bacteri*’ OR ‘fungi*’) AND (‘behav*’ OR ‘attract*’ OR ‘repel*’ OR ‘pref*’) resulting in 1615 papers. Following assessment of title and abstract, we identified 180 potential papers of interest, from which 70 provided suitable data for inclusion in the meta-analysis (Figure 1).

Summary of selection process for final papers for inclusion in the meta-analysis, as based on PRISMA guidelines
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
We screened the studies using the following inclusion criteria: (1) data must come from experimental or field studies that tested the bacteria effect on insects comparing a bacteria-treated with control untreated plants; (2) data are presented to enable extraction of mean values, a measure of variance, and sample size; (3) measured variables are related to insect behaviour (host choice or consumption rate) or fitness (insect survival, development time, body size, fecundity, or lifespan). We extracted the mean, variance, and sample size across the treated and untreated groups. We also extracted experimental information on bacteria inoculum (single strain or multi-strain community), microbial genus/species (multi-genus noted for mixed-genera community inoculations), insect feeding type (chewing herbivore, sucking herbivore, parasitoid wasps or predator), insect family/species, plant family/species, and study environment (field, greenhouse, or growth chamber). Following in-depth screening of 180 papers, 44 papers were removed due to not containing primary data, 66 removed due to lack of informative data across treatments and/or controls, resulting in a final data set of 344 data points extracted from 70 papers published between 1997 and 2023 fulfilling all the inclusion criteria.
In studies with multiple genotypes, strains, or varieties, data were pooled within species due to lack of replication to examine specific effects. We selected only data from the ‘wildtype’ microbial strain where data were presented also on lab-generated mutants. For controlled environment time-series experiments we chose the time that related best to the other studies in the dataset; for example, the number of offspring produced by aphids is commonly measured at day 10 or 14. For field data, we averaged across multiple sampling time points, as this best represented the data across the included studies.
2.1 Data analysis
The meta-analysis was performed in RStudio version 2024.04.2 (RStudio Team, 2024) using R version 4.3.0 (R Core Team, 2024) using the package metaphor (Viechtbauer, 2010). Effect sizes were calculated using the standardized mean difference with an unbiased estimate of the sampling variances (SMDH, giving Hedges’ g). This measurement compares the treated plants (inoculated with the bacteria and exposed to the herbivore insect) to the control plants (that were just exposed to the herbivore insect). A priori power analyses (medium effect size d=0.5) were calculated in R following Quintana and Tiebel (2019) for the main insect variables within the analysed data subset. For the power analysis, we used the number of effect sizes from the extracted data (data points), the average number of replicates used to calculate each effect size (control vs treated plant), against a predicted effect size of d = 0.5 (medium) and low heterogeneity (h = 0.33).
The effect of the inoculated bacteria on the insect traits was analysed with a meta-analytic linear mixed effect model (rma.mv). Study was included as a random effect to account for multiple data points across insect, plant, and bacteria species within each study. In each insect subset (sucking and chewing herbivores), insect response was treated as a fixed effect moderator (levels: host choice, feeding, survival, development, body size, fecundity, lifespan) also separated by main rhizobacteria genera (levels: Bacillus, Pseudomonas, Other) using a nested analysis approach. Rhizobacteria were analysed at these levels due to lack of replication at species or strain level, the ‘other’ category includes all non-Bacillus and non-Pseudomonas strains. Additional models were used to determine the effect across different plant and insect families, nesting rhizobacteria group within family for the model analysis, as well as across experimental manipulations (controlled pot experiments (growth chamber and greenhouse) vs field trials). Importantly, results are presented when there were at least three data points from three independent studies for the comparisons, unless noted. We evaluated publication bias in the dataset by assessing the funnel-plot asymmetry and Eggers test (Supplementary Data S1). For the data visualisation, figures show both the mean effect size and 95% confidence intervals obtained from the linear mixed effect models output, as well as the individual-level data as a bubble plot with sizes relative to the precision of the estimate (1/SE), where larger bubbles show a higher certainty of the data. k corresponds to the number of effect sizes, from n number of studies.
3 Results
3.1 Effects of rhizobacteria inoculation of plants on herbivores and their natural enemies
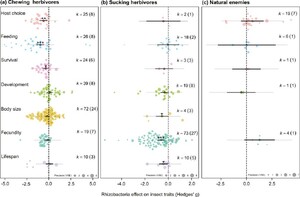
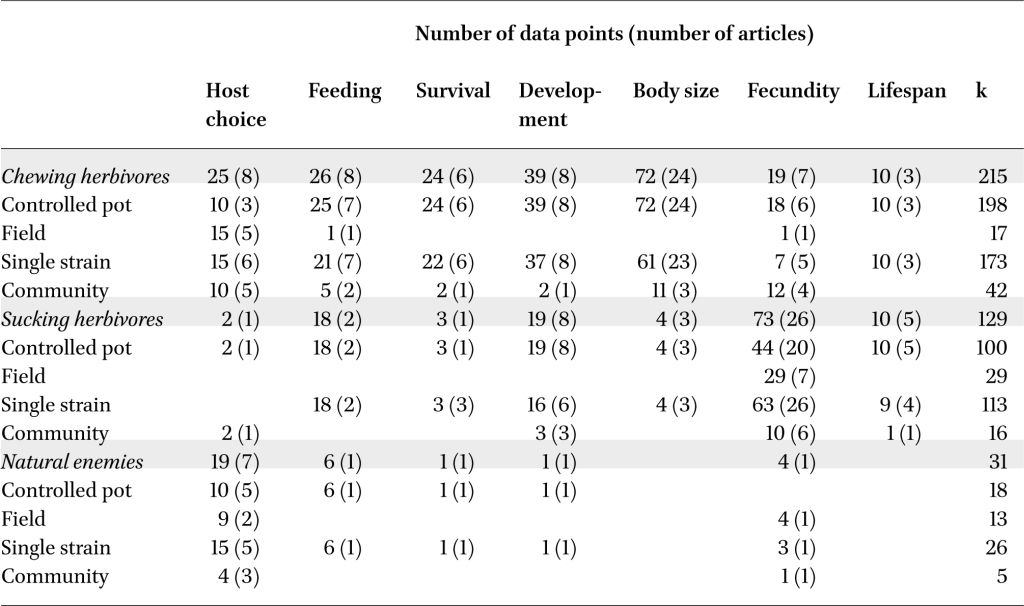
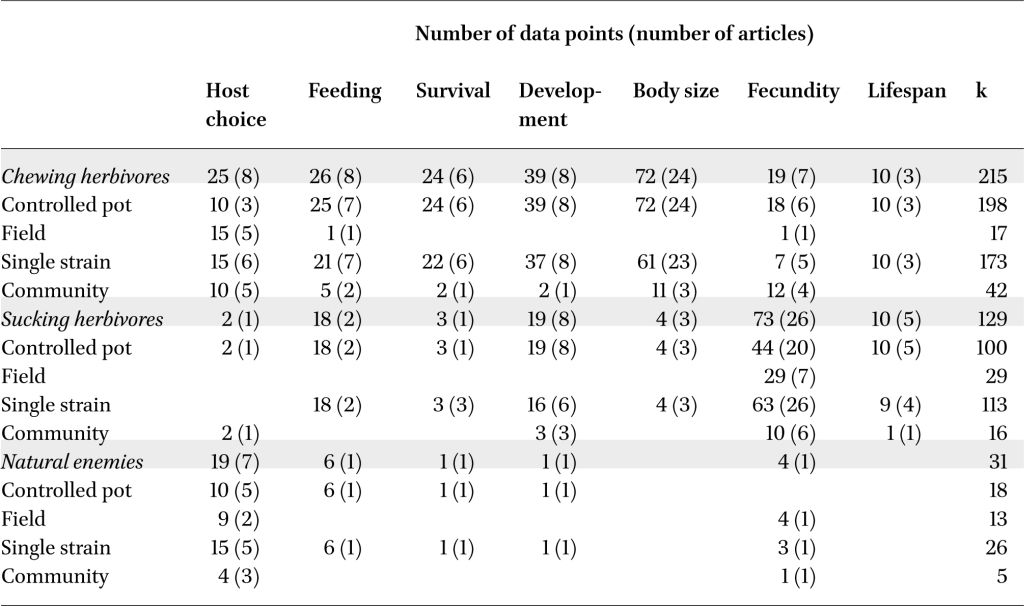
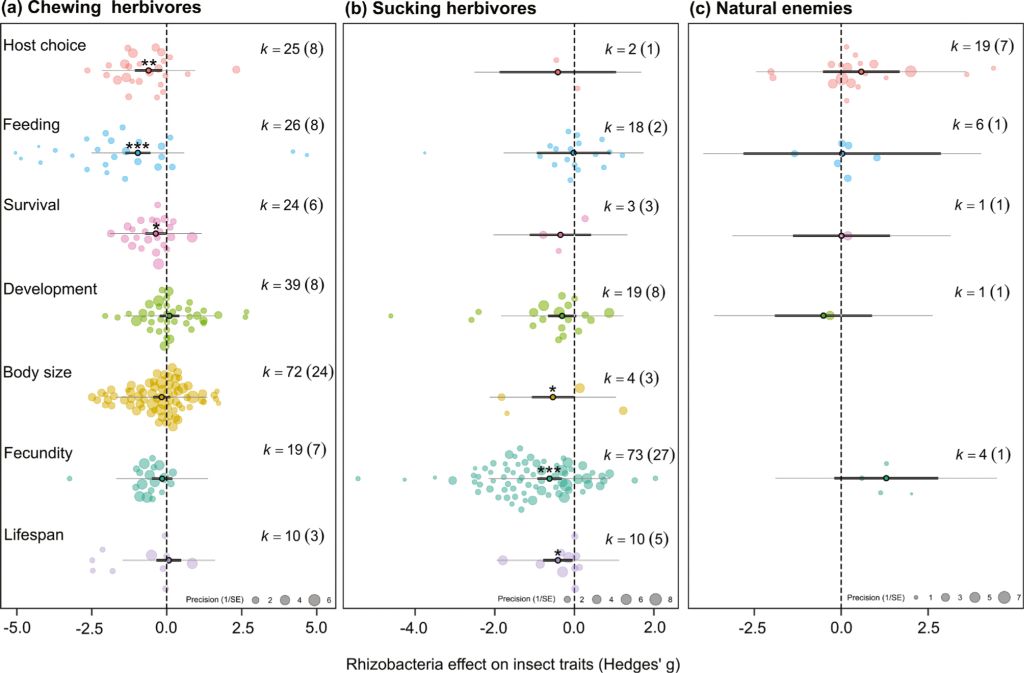
The herbivore final dataset contained 344 data points from 66 papers, representing 215 data points (38 papers) from chewing herbivores and 129 data points (31 papers) from sucking herbivores, with data on all responses identified (Table 1). Across all measured response traits in all herbivore species, rhizobacteria inoculation of plants had a negative effect on insect herbivores (hedges g = –0.417, P<0.001, k=344, n=66). Yet, the strength and significance of these effects varied between chewing (hedges g = –0.563, P<0.001, k=215, n=38) and sucking herbivores (hedges g = –0.242, P=0.209, k=129, n=31), as well as across the different response traits measured (test of moderators QM=59.3, df=14, P<0.001; Figure 2A,B). A large residual heterogeneity in this model (QM=1416.3, df=330, P<0.001) indicated the need to include additional factors in the model (rhizobacteria/plant/insect species or system information) and therefore in following sections we examined chewing and sucking herbivore datasets separately.
Summary of data points included in meta-analyses, separated by seven behavioural and fitness traits commonly measured, exploring the distribution of data points among experimental factorsa
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
We also extracted 31 data points from eight papers with information on rhizobacterial effects on insect natural enemies (predators and parasitoid, examining variables including host choice, feeding behaviour, and fecundity; Table 1, Figure 2C). While there were no general significant effects, likely due to the lack of data (hedges g = 0.416, P=0.385, k=31, n=8), there is a potential pattern of natural enemy preference to plants inoculated with non-Bacillus or non-Pseudomonas rhizobacteria (‘Other’ rhizobacteria: hedges g = 1.61, P=0.050, k=4, n=3) with no effect for Bacillus (hedges g = 0.232, P=0.810, k=11, n=2) or Pseudomonas inoculants (hedges g = –0.649, P=0.513, k=4, n=2). The other rhizobacteria grouping included plants inoculated with Bradyrhizobium japonicum, Delftia acidovorans, and various multi-genus consortia.
Effect of rhizobacterial inoculation across response traits measured for (A) chewing herbivores, (B) sucking herbivores and (C) natural enemies (parasitoid wasps and predators). Orchard plots showing effect size mean, thick bars – 95% confidence intervals, thin bars – 95% prediction intervals, and individual effect sizes scaled by their precision (1/SE). k corresponds to the number of effect sizes; the number of studies is shown in brackets. Effects are considered significant if the 95% CI does not overlap with zero.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
3.2 Effect of rhizobacteria inoculation on chewing insects
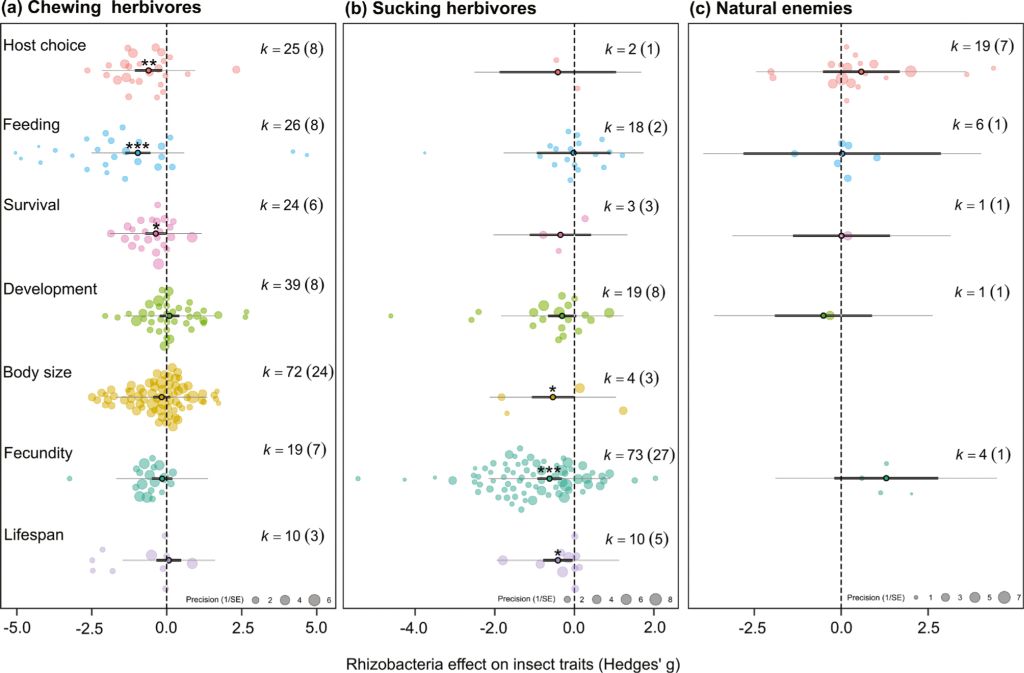
The chewing insects belonged to six different families (Lepidoptera k=178, Chrysomelidae k=19, Anthomyiidae k=15, Cecidomyiidae k=1, Gryllotalpidae k=1, Sciaridae k=1) and the Lepidoptera represented nine genera, including Spodoptera sp. (k=48 data points, from n=13 papers) and Plutella xylostella (k=67 data points, from n=4 papers); with a tendency for a small number of papers to examine a broad range of rhizobacteria leading to a loss of independence in these effect estimates.
Effects on chewing insect behaviour were observed across the range of bacterial inoculants leading to overall significant reductions in the number of insects landing on an inoculated host plant (host choice: hedges g = –0.637, P=0.006, k=25, n=8) with subsequent reduction in leaf consumption (feeding: hedges g = –0.965, P<0.001, k=26, n=8) (Figure 2A, 3A). There was also a significant negative effect on chewing herbivore survival (hedges g = –0.367, P=0.041, k=24, n=6; Figure 2a), but this was driven by two studies using several strains of Bacillus inoculants (hedges g = –0.602, P=0.028, k=11, n=2; Figure 3A). No significant effect was observed for studies using Pseudomonas inoculants (hedges g = –0.131, P=0.656, k=8, n=3; Figure 3A) or ‘Other’ inoculants (hedges g = –0.150, P=0.586, k=5, n=2; Figure 3A). Despite body size being the most measured variable, it showed no strong significant reduction when averaged across all rhizobacteria inoculants (hedges g = –0.170, P=0.248, k=72, n=24; Figure 2A). However, when grouped by rhizobacteria genus, Bacillus inoculations of host plants significantly reduced chewing herbivore body size (hedges g = –0.487, P=0.032, k=24, n=6), while Pseudomonas and ‘Other’ inoculations (representing nine genera) did not (Pseudomonas: hedges g = –0.051, P=0.820, k=39, n=13; Other: hedges g = –0.062, P=0.804, k=9, n=6; Figure 3A). Insect development time was positively affected by Bacillus inoculation of the host plant, representing shorter development time, but only from two studies (hedges g = 0.561, P=0.026, k=19, n=2).
Effect of rhizobacterial inoculation on chewing herbivores, separated by Bacillus spp. inoculants (red), Pseudomonas spp. inoculants (blue), and other rhizobacteria (grey) across (A) response traits (B) main insect families, and within Lepidoptera and (C) host plant family. Effect size mean and 95% confidence intervals from the meta-analysis linear mixed effect model outputs, individual data shown as bubble plots with sizes relative to the precision of the estimate (1/SE), where larger bubbles show a higher certainty of the data. Number of data points (k) shown in brackets.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
Of the six chewing herbivore families studied, rhizobacteria inoculation significantly reduced chewing herbivore fitness within Chrysomelidae (hedges g = –0.584, P=0.030, k=19, n=8) and Lepidoptera (hedges g = –0.382, P=0.008, k=178, n=26) across a diverse set of microbial inoculants. Chrysomelidae leaf beetles were negatively affected by non-Bacillus bacterial inoculants of their host plants (Other: hedges g = –0.844, P=0.023, k=5, n=4; Pseudomonas: hedges g = –0.628, P=0.098, k=5, n=2; Bacillus: hedges g = –0.433, P=0.218, k=9, n=4; Figure 3b). Whereas, within Lepidoptera the strongest effects were from Bacillus inoculants (Spodoptera: hedges g = –0.933, P=0.021, k=26, n=3; Other Lepidoptera: hedges g = –1.00, P=0.089, k=20, n=3; Figure 3B). There were no significant effects on Plutella Lepidopterans owing to the broad variation in observed outcomes, despite the high number of data points, these are collected from only a few independent studies (Bacillus: hedges g = –0.459, P=0.212, k=46, n=3; Other: hedges g = –0.629, P=0.121, k=20, n=1; Figure 3B).
Chewing insects were most often studied on Brassicaceae (120/215) or Poaceae (73/215) plants with both Chrysomelidae and Lepidoptera studied within both plant families. Other plant families studied include Malvaceae (9/215) Cucurbitaceae (5/215), Fabaceae (4/215), Solanaceae (3/215) and Geraniaceae (1/215). Bacillus inoculants used in Brassicaceae and Poaceae systems had negative effects on insects (Poaceae: hedges g = –0.818, P=0.006, k=40, n=5; Brassicaceae: hedges g = –0.663, P=0.051, k=51, n=4; Figure 3C). Other microbial inoculants (including Alcaligenes, Enterobacter, and Kluyvera) also showed significant effects on chewing insects but data were collected from a single paper and thus robustness is low (hedges g = 0.826, P=0.030, k=20, n=1; Figure 3C). Pseudomonas inoculants had no overall effect within Brassicaceae (hedges g = –0.161, P=0.405, k=49, n=12; Figure 2C) or Poaceae (hedges g = 0.152, P=0.738, k=5, n=3; Figure 3C).
3.3 Effect of rhizobacteria inoculation on sucking herbivores
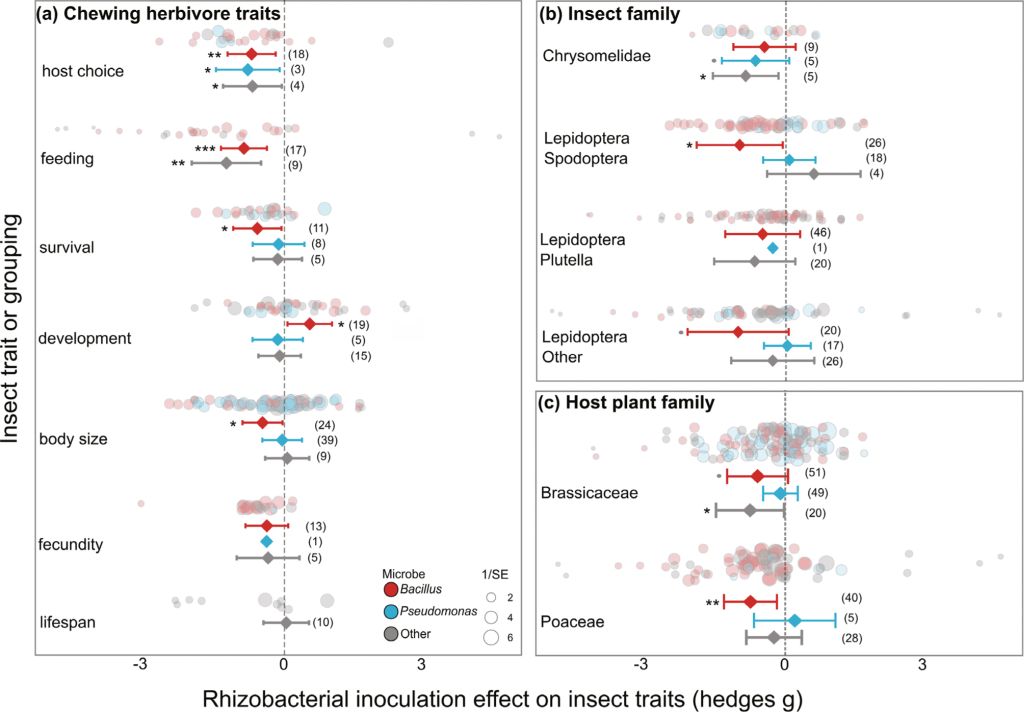
The data were dominated by studies on Aphididae (86/129 data points from 8 species of aphid); these included Brevicoryne brassicae (n=23), Acyrthosiphon pisum (n=18), Myzus persicae (n=14), Rhopalosiphum padi (n=13), Lipaphis erysimi (n=18), Aphis glycines (n=5), undescribed aphids (n=4), Aphis gossypii (n=3), and Sitobion avenae (n=2). The remaining data were from Tetranychidae spider-mites (n=28 on Tetranychus urticae), Aleyrodidae whitefly (n=7 from Bemisia tabaci and n=4 from Aleyrodes proletella), Triozidae psyllids (n=3, from Bactericera cockerelli), and n=1 from the Cicadellidae leafhopper Amrasca biguttula.
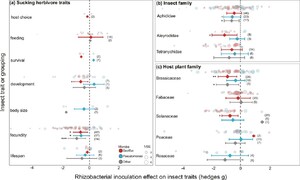
Most studies measured sucking herbivore fecundity (number of offspring produced) (73/129) which was significantly reduced by rhizobacteria inoculation of the host plants (hedges g = –0.624, P<0.001, k=73, n=26; Figure 2B). Averaged across all inoculants, we also found significant effects on sucking herbivore lifespan (hedges g = –0.413, P=0.030, k=10, n=5), and body size (hedges g = –0.539, P=0.046, k=4, n=3). Similar to the chewing herbivores, the majority of experiments on sucking herbivores used either Bacillus (n=71/129) or Pseudomonas strains (n=33/129). We found substantial variation in the number of studies and strength of effects across these different inoculants (Figure 4A). Insect fecundity was significantly reduced by Bacillus (hedges g = –0.565, P=0.036, k=42, n=12), Pseudomonas spp. (hedges g = –0.559, P=0.033, k=17, n=9), and other inoculant species (hedges g = –0.922, P<0.001, k=14, n=10). A potential negative impact on development time to adult (hedges g = –0.308, P=0.090, k=19, n=8) was driven by a significant negative effect on Bacillus inoculated plants (hedges g = –0.726, P=0.011, k=7, n=3) countered by potential positive effects on Pseudomonas inoculated plants (hedges g = –0.544, P=0.065, k=7, n=4).
Effect of rhizobacterial inoculation on sucking herbivores, separated by Bacillus spp. inoculants (red), Pseudomonas spp. inoculants (blue), and other rhizobacteria (grey) across (A) response traits (B) main insect families, and (C) host plant family. Effect size mean and 95% confidence intervals from the meta-analysis linear mixed effect model outputs, individual data shown as bubble plots with sizes relative to the precision of the estimate (1/SE), where larger bubbles show a higher certainty of the data. Number of data points (k) shown in brackets.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
The three insect families with sufficient data for analysis were all negatively impacted by rhizobacterial inoculation of the host plants (Aphididae: hedges g = –0.469, P=0.005, k=86, n=24; Aleyrodidae: hedges g = –0.767, P=0.027, k=11, n=5; Tetranychidae: hedges g = –0.607, P=0.059, k=28, n=3). In particular, Aleyrodidae insects (whitefly) were significantly impacted by Bacillus inoculants (hedges g = –1.086, P=0.016, k=8, n=3; Figure 4B), while Aphididae (aphids) were most negatively affected by non-Bacillus inoculants (‘Other’: hedges g = –0.589, P=0.027, k=17, n=10; Pseudomonas: hedges g = –0.514, P=0.097, k=23, n=8; Figure 4B). For aphid studies, the other inoculants included Rhizobium sp, Bradyrhizobium sp, Sinorhizobium sp, and an Acidovorax sp.
Sucking insects were studied across seven different plant families, Brassicaceae (39/129), Fabaceae (28/129), Solanaceae (26/129), Poaceae (20/129), Rosaceae (12/129), Malvaceae (3/129), Cucurbitaceae (1/129) (Figure 4C, excluding Malvaceae and Cucurbitaceae due to low data). We found significant negative effects of rhizobacterial inoculation on sucking insects feeding on plants within Brassicaceae (hedges g = –0.755, P=0.001, k=39, n=10), driven by effects of Bacillus spp. (hedges g = –0.931, P=0.006, k=26, n=4) and other inoculants (hedges g = –0.933, P=0.022, k=3, n=2), but not Pseudomonas (hedges g = –0.355, P=0.277, k=10, n=4) (Figure 4C). Bacillus spp. inoculants also significantly impacted sucking insects on Solanaceae plants (hedges g = –0.753, P=0.011, k=20, n=5; Figure 4C).
3.4 Effect of rhizobacteria community inoculations (multi-Bacillus and multi-genus inoculants)
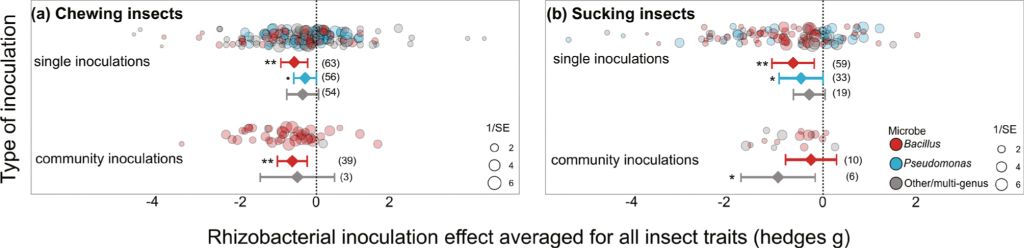
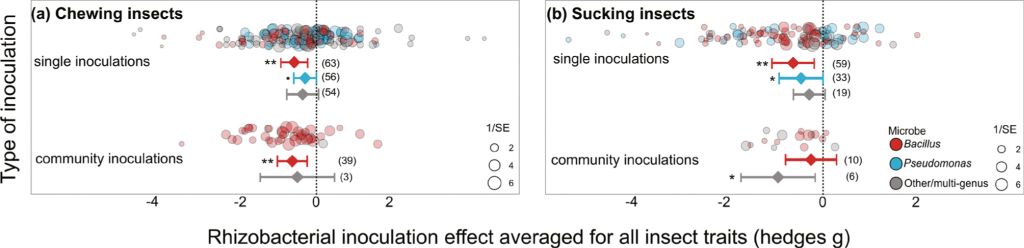
Chewing herbivores were slightly more negatively affected when feeding on plants inoculated with multiple rhizobacteria (hedges g = –0.383, P=0.043, k=42, n=9) than single inoculants (hedges g = –0.273, P=0.051, k=173, n=34) (community vs single: QM= 5.04, df=2, P=0.081). This difference was driven by the majority of community inoculations using multiple Bacillus strains (hedges g = –0.653, P=0.002, k=39, n=7) rather than multi-genus communities, for which there was insufficient information for a robust analysis (hedges g = –0.511, P=0.330, k=3, n=2). The effect size of multi-strain Bacillus inoculants on chewing herbivores was comparable to single-strain Bacillus inoculations (hedges g = –0.598, P=0.002, k=63, n=10; Figure 5A). Single-strain inoculants from Pseudomonas had only minimal overall effect on chewing herbivores (hedges g = –0.296, P=0.064, k=56, n=16; Figure 5A), and no multi-strain Pseudomonas communities were tested.
Effect of rhizobacterial inoculation using single and community inoculations for (A) chewing or (B) sucking herbivores. Effect size mean and 95% confidence intervals from the meta-analysis linear mixed effect model outputs, individual data shown as bubble plots with sizes relative to the precision of the estimate (1/SE), where larger bubbles show a higher certainty of the data. Number of data points (k) shown in brackets.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
In contrast, sucking insects were more negatively affected by single strain inoculations (hedges g = –0.552, P=0.003, k=113, n=28) than the less-studied community inoculations (hedges g = –0.348, P=0.094, k=16, n=8). Community inoculations were dominated by those using multi-strain Bacillus inoculants that, surprisingly, had no significant effect on insect fitness (hedges g = –0.236, P=0.393, k=10, n=5) despite single-strain Bacillus inoculants having a strong effect (hedges g = –0.624, P=0.007, k=59, n=10). Of particular note, multi-genus inoculants were able to reduce sucking insect fitness yet examined across very few studies (hedges g = –1.016, P=0.010, k=6, n=3). Multi-genus inoculants contained various combinations of Bradyrhizobium spp., Azospirillium spp., Delftia spp., Azotobacter spp., as well as Bacillus spp. and Pseudomonas spp.
3.5 Effect of controlled pot experiments compared to field trials
For chewing herbivores, only the host-choice of insects to Bacillus-inoculated plants could be analysed to compare field choice and controlled pot experiments since other variables were only measured in controlled pot experiments. Similarly, for sucking insects only insect population sizes were measured (fecundity) across both environments, with multiple bacterial inoculants (minimum of three data points per microbe grouping per environment). We found that for these variables that the significant effect of rhizobacteria in controlled pot experiments (chewing: hedges g = –1.244, P=0.023, k=9, n=3; sucking: hedges g = –0.748, P=0.004, k=44, n=20) was lost in field trials (chewing: hedges g = –0.646, P=0.235, k=9, n=5; sucking: hedges g = –0.449, P=0.256, k=29, n=8) (Figure 6).
Effect of rhizobacterial inoculation comparing controlled pot and field trials for (A) chewing herbivore using host-choice and Bacillus sp. only due to lack of data for others, and (B) sucking herbivore fecundity combining all microbes studied. Effect size mean and 95% confidence intervals from the meta-analysis linear mixed effect model outputs, individual data shown as bubble plots with sizes relative to the precision of the estimate (1/SE), where larger bubbles show a higher certainty of the data. Number of data points (k) shown in brackets.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10003
4 Discussion
Overall, inoculation of host plants with rhizobacteria reduces insect fitness and alters behaviour to further reduce colonisation (host choice) and leaf consumption (feeding). We also observed potential for rhizobacteria inoculation to increase host choice of natural enemies of herbivore insects, as an additional indirect benefit via increased top-down biocontrol. This supports continued efforts to understand how rhizobacteria boost plant resistance to herbivores and a need to explore indirect effects via recruitment of natural enemies in greater detail.
Variable effects across chewing and sucking herbivores were observed with rhizobacterial inoculation of host plants more strongly reducing chewing insect behaviours (host choice and feeding) than fitness traits, while sucking herbivores were primarily impacted through reduced reproduction (fecundity) with effects also for reduced body sizes and life span. This is likely a reflection of the feeding behaviour. Sucking insects feed on plant sap using modified tube-like mouthparts causing less mechanical damage to plant tissues than chewing herbivores (Walling, 2000). This mode of feeding by sucking insects means they may by-pass some of the plant’s immediate defences triggered by tissue damage, but may be more susceptible to other induced chemical defences following sap-ingestion that impact their ability to reproduce (Züst and Agrawal, 2016). In contrast, chewing insects disrupt plant tissues more extensively, triggering induction of rhizobacteria-primed broad-spectrum plant defences. These may be more immediate and thus influence host-choice and feeding rate before any detectable effect on growth or reproductive output.
Several studies showed that rhizobacteria of plants can attract natural enemies, through increased host choice towards inoculated plants. Indeed, while overall the meta-analysis result was non-significant, the subset of ‘other’ rhizobacteria, excluding Bacillus and Pseudomonas strains but including single and multi-genus combinations (Benítez et al., 2017; Gadhave and Gange, 2022; Godschalx et al., 2023; Pulido et al., 2019), showed a significant effect. The broad variation and low number of studies indicates a need to further study these interactions. Rhizobacteria-mediated changes in plant volatiles (Bouwmeester et al., 2025) that attract natural enemies of herbivores would provide a valuable indirect biocontrol effect in addition to direct effects on herbivore fitness.
There were strong differences in herbivores responses across the rhizobacteria groups, with Bacillus spp. (Blake et al., 2021) inoculants providing stronger general effects than the commonly studied Pseudomonas spp. (Pieterse et al., 2021). We grouped all other rhizobacteria species since alone they had insufficient data, but they also significantly impacted insects across various traits. These effects were not restricted to specific insect or plant families also highlighting the generalisability of rhizobacterial inoculation for plant protection (Biere and Bennett, 2013; Gruden et al., 2020). While most chewing herbivores studied were Lepidopterans, we observed significant behaviour and fitness effects of rhizobacteria on Chrysomelidae leaf beetles and variation among Lepidopteran sub-families. Across host plants, Bacillus inoculations were most effective against chewing herbivores for both Brassicaceae and Poaceae plants with limited effect of Pseudomonas inoculants. Aphids were the most studied sucking herbivores, most affected by Pseudomonas spp. and ‘other’ rhizobacteria, many of which belonged to nitrogen-fixing rhizobia species. This may indicate an interplay between plants and microbes that can modulate nutrient availability and induce plant defences, i.e. as biostimulants (plant-growth-promoting rhizobacteria) and bioprotectants (priming and induction of defence pathways).
Single-strain inoculants generally performed better than multi-strain or community inoculations (stronger reduction of insect traits). This may reflect strain incompatibility rather than loss of strain efficacy, since many studies assemble communities based on taxonomic diversity rather than functional compatibility (Song et al., 2020). Interestingly, for the sucking herbivores, single-strain Bacillus inoculants performed better than multi-strain Bacillus inoculants, indicating potential strain incompatibility could affect outcomes (Díaz et al., 2023). The reduced abundance of Bacillus strains in the rhizosphere over time following inoculation (Moshe et al., 2024) may also impact variable effectiveness of a multi-strain community, particularly if they use similar resources that diminish over time. This may also explain the increased effectiveness of multi-genus inoculants for reducing sucking insect fitness on inoculated plants, as observed in the meta-analysis results. The potential of developing diverse community inoculants (also termed SynComs, synthetic communities (Bai et al., 2015; Lebeis et al., 2015) to provide a broad array of benefits is high, and as such is a keen focus of current plant microbiome research (Fagorzi et al., 2023; He et al., 2024; Marin et al., 2021; Pascale et al., 2020; Poudel et al., 2023; Shayanthan et al., 2022; Wang et al., 2024). Strain and functional compatibility will be key to enable SynComs to persist and deliver benefits to the host plants over time (De Souza et al., 2020; Hayashi et al., 2024; O’Banion et al., 2020; Song et al., 2020).
The majority of experimental work is conducted in environmentally-controlled conditions, often using sterilise soil in sterilised pots and watered with sterilised water. This is a good approach to uncover the effect of a focal inoculated strain, but it has been difficult to translate to the field which is messy and diverse. Our results showed that while the overall effects were significant for pot-based experiments but not for the field trials, there were several field trials reporting highly significant outcomes. A limitation of this data is that it was restricted to host choice (only on Bacillus spp. inoculated plants) for chewing herbivores, and fecundity (averaged across all microbes, but mostly Bacillus spp. or rhizobia) for sucking herbivores. Nevertheless, our results suggest that the increased native microbiota of field soils and environmental variability could act to reduce the ability of inoculants to colonise and persist among plant roots leading to reduced effectiveness compared to controlled pots experiments (Frew et al., 2022; Gruden et al., 2020; He et al., 2024).
In conclusion, there is great potential for using rhizobacteria to increase plant resistance to insect pests but these meta-analysis results show that we need to consider the broader interacting environment when developing effective inoculants (Shayanthan et al., 2022; Song et al., 2020). This will involve understanding which rhizobacteria are important for specific plants, against specific insects or under specific environments, but also those that provide broad benefits across multiple ecological systems. With this knowledge, we can develop microbial inoculants that contain multiple beneficial species that synergistically interact to benefit the plant with broad acting functions across multiple plants, crops and environments.
Supplementary material
Supplementary material can be found online at https://doi.org/10.6084/m9.figshare.30111244
Supplementary Data S1. Meta-analysis data.
Acknowledgements
This work was supported by a BBSRC (UKRI) David Phillips Fellowship BB/S010556/1 to SEZ, BBSRC NLD studentship to MP, and German Research Council, Deutsche Forschungsgemeinschaft (DFG; project number 397565003) funding to SZ and OSM, and a BBSRC (UKRI) European Partnerships grant to SZ (BB/W018578/1).
Conflict of interest
The authors have declared no conflict of interest.
References
Bai, Y., Müller, D.B., Srinivas, G., Garrido-Oter, R., Potthoff, E., Rott, M., Dombrowski, N., Münch, P.C., Spaepen, S. and Remus-Emsermann, M., 2015. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 528: 364–369.
Bakker, P.A., Berendsen, R.L., Doornbos, R.F., Wintermans, P.C. and Pieterse, C.M., 2013. The rhizosphere revisited: root microbiomics. Frontiers in Plant Science 4: 165.
Benítez, E., Paredes, D., Rodríguez, E., Aldana, D., González, M., Nogales, R., Campos, M. and Moreno, B., 2017. Bottom-up effects on herbivore-induced plant defences: a case study based on compositional patterns of rhizosphere microbial communities. Scientific Reports 7: 6251.
Berendsen, R.L., Pieterse, C.M. and Bakker, P.A., 2012. The rhizosphere microbiome and plant health. Trends in Plant Science 17: 478–486. https://doi.org/10.1016/j.tplants.2012.04.001
Biere, A. and Bennett, A.E., 2013. Three-way interactions between plants, microbes and insects. Functional Ecology 27: 567–573. https://doi.org/10.1111/1365-2435.12100
Blake, C., Christensen, M.N. and Kovács, Á.T., 2021. Molecular aspects of plant growth promotion and protection by Bacillus subtilis. Molecular Plant-Microbe Interactions 34: 15–25. https://doi.org/10.1094/MPMI-08-20-0225-CR
Blubaugh, C.K., Carpenter-Boggs, L., Reganold, J.P., Schaeffer, R.N. and Snyder, W.E., 2018. Bacteria and competing herbivores weaken top-down and bottom-up aphid suppression. Frontiers in Plant Science 9: 1239. https://doi.org/10.3389/fpls.2018.01239
Bouwmeester, H., Dong, L., Wippel, K., Hofland, T. and Smilde, A., 2025. The chemical interaction between plants and the rhizosphere microbiome. Trends in Plant Science. https://doi.org/10.1016/j.tplants.2025.06.001
Da Silva, F.B., Barbosa, J.Z., Tiecher, T., Borin, J.B.M., Treichel, B. and de Sá, E.L.S., 2024. The species-dependent effect of PGPR co-inoculation in legume plants: A global meta-analysis. Rhizosphere 30: 100869.
De Souza, R.S.C., Armanhi, J.S.L. and Arruda, P., 2020. From microbiome to traits: designing synthetic microbial communities for improved crop resiliency. Frontiers in Plant Science 11: 1179.
De Zutter, N., Ameye, M., Bekaert, B., Verwaeren, J., De Gelder, L. and Audenaert, K., 2022. Uncovering new insights and misconceptions on the effectiveness of phosphate solubilizing rhizobacteria in plants: a meta-analysis. Frontiers in Plant Science 13: 858804.
Díaz, P.R., Merlo, F., Carrozzi, L., Valverde, C., Creus, C.M. and Maroniche, G.A., 2023. Lettuce growth improvement by Azospirillum argentinense and fluorescent Pseudomonas co-inoculation depends on strain compatibility. Applied Soil Ecology 189: 104969. https://doi.org/10.1016/j.apsoil.2023.104969
Fagorzi, C., Passeri, I., Cangioli, L., Vaccaro, F. and Mengoni, A., 2023. When biodiversity preservation meets biotechnology: The challenge of developing synthetic microbiota for resilient sustainable crop production. Journal of Sustainable Agriculture and Environment 2: 5–15. https://doi.org/10.1002/sae2.12038
Frew, A., Antunes, P.M., Cameron, D.D., Hartley, S.E., Johnson, S.N., Rillig, M.C. and Bennett, A.E., 2022. Plant herbivore protection by arbuscular mycorrhizas: a role for fungal diversity? New Phytologist 233: 1022–1031.
Friman, J., Pineda, A., van Loon, J.J. and Dicke, M., 2021. Bidirectional plant‐mediated interactions between rhizobacteria and shoot‐feeding herbivorous insects: a community ecology perspective. Ecological Entomology 46: 1–10.
Gadhave, K.R. and Gange, A.C., 2022. Soil-dwelling Bacillus spp. affects aphid infestation of calabrese and natural enemy responses in a context-specific manner. Agricultural and Forest Entomology 24: 618–625. 10.1111/afe.12507
Godschalx, A.L., Diethelm, A.C., Kautz, S. and Ballhorn, D.J., 2023. Nitrogen-fixing rhizobia affect multitrophic interactions in the field. Journal of Insect Behavior 36: 168–179.
Gruden, K., Lidoy, J., Petek, M., Podpečan, V., Flors, V., Papadopoulou, K.K., Pappas, M.L., Martinez-Medina, A., Bejarano, E., Biere, A. and Pozo, M.J., 2020. Ménage à trois: unraveling the mechanisms regulating plant – microbe – arthropod interactions. Trends in Plant Science 25: 1215–1226. https://doi.org/10.1016/j.tplants.2020.07.008
Hayashi, I., Fujita, H. and Toju, H., 2024. Deterministic and stochastic processes generating alternative states of microbiomes. ISME Communications 4: ycae007. 10.1093/ismeco/ycae007
He, X., Wang, D., Jiang, Y., Li, M., Delgado-Baquerizo, M., McLaughlin, C., Marcon, C., Guo, L., Baer, M., Moya, Y.A.T., von Wiren, N., Deichmann, M., Schaaf, G., Piepho, H.P., Yang, Z., Yang, J., Yim, B., Smalla, K., Goormachtig, S., de Vries, F.T., Huging, H., Baer, M., Sawers, R.J.H., Reif, J.C., Hochholdinger, F., Chen, X. and Yu, P., 2024. Heritable microbiome variation is correlated with source environment in locally adapted maize varieties. Nature Plants 10: 598–617. https://doi.org/10.1038/s41477-024-01654-7
Lebeis, S.L., Paredes, S.H., Lundberg, D.S., Breakfield, N., Gehring, J., McDonald, M., Malfatti, S., Glavina del Rio, T., Jones, C.D. and Tringe, S.G., 2015. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 349: 860–864.
Li, J., Wang, J., Liu, H., Macdonald, C.A. and Singh, B.K., 2022. Application of microbial inoculants significantly enhances crop productivity: a meta‐analysis of studies from 2010 to 2020. Journal of Sustainable Agriculture and Environment 1: 216–225.
Marin, O., Gonzalez, B. and Poupin, M.J., 2021. From microbial dynamics to functionality in the rhizosphere: a systematic review of the opportunities with synthetic microbial communities. Frontiers in Plant Science 12: 650609. https://doi.org/10.3389/fpls.2021.650609
Moshe, M., Frenkel, O., Sela, N., Davidovich, C., Amutuhaire, H., Banin, E. and Cytryn, E., 2024. Persistence and microbiome modification in Rhizoctonia solani-inoculated rhizosphere following amendment with a Bacillus biocontrol agent. Phytobiomes Journal 8: 611–620. 10.1094/pbiomes-01-24-0006-r
O’Banion, B.S., O’Neal, L., Alexandre, G. and Lebeis, S.L., 2020. Bridging the gap between single-strain and community-level plant-microbe chemical interactions. Molecular Plant-Microbe Interactions 33: 124–134.
Pan, J., Peng, F., Xue, X., You, Q., Zhang, W., Wang, T. and Huang, C., 2019. The growth promotion of two salt-tolerant plant groups with PGPR inoculation: a meta-analysis. Sustainability 11: 378.
Pascale, A., Proietti, S., Pantelides, I.S. and Stringlis, I.A., 2020. Modulation of the root microbiome by plant molecules: the basis for targeted disease suppression and plant growth promotion. Frontiers in Plant Science 10. https://doi.org/10.3389/fpls.2019.01741
Pieterse, C.M., Berendsen, R.L., de Jonge, R., Stringlis, I.A., Van Dijken, A.J., Van Pelt, J.A., Van Wees, S., Yu, K., Zamioudis, C. and Bakker, P.A., 2021. Pseudomonas simiae WCS417: star track of a model beneficial rhizobacterium. Plant and Soil 461: 245–263. https://doi.org/10.1007/s11104-020-04786-9
Pieterse, C.M., Poelman, E.H., Van Wees, S.C. and Dicke, M., 2013. Induced plant responses to microbes and insects. Frontiers in Plant Science 4: 475.
Pieterse, C.M.J., Zamioudis, C., Berendsen, R.L., Weller, D.M., Van Wees, S.C.M. and Bakker, P., 2014. Induced systemic resistance by beneficial microbes. Annual Review of Phytopathology 52: 347–375. 10.1146/annurev-phyto-082712-102340
Pineda, A., Zheng, S.-J., van Loon, J.J.A., Pieterse, C.M.J. and Dicke, M., 2010. Helping plants to deal with insects: the role of beneficial soil-borne microbes. Trends in Plant Science 15: 507–514. 10.1016/j.tplants.2010.05.007
Poudel, R., Jumpponen, A., Kennelly, M.M., Rivard, C., Gomez-Montano, L. and Garrett, K.A., 2023. Integration of phenotypes in microbiome networks for designing synthetic communities: a study of mycobiomes in the grafted tomato system. Applied and Environmental Microbiology 89: e01843–01822. doi:10.1128/aem.01843–22
Pulido, H., Mauck, K.E., De Moraes, C.M. and Mescher, M.C., 2019. Combined effects of mutualistic rhizobacteria counteract virus-induced suppression of indirect plant defences in soya bean. Proceedings of the Royal Society B – Biological Sciences 286: 20190211. https://doi.org/10.1098/rspb.2019.0211
Quintana, D. and Tiebel, J., 2019. An R script and excel file to calculate statistical power for your meta-analysis. Available at: https://osf.io/5c7uz/.10.17605/OSF.IO/5C7UZ
R Core Team, 2024. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
RStudio Team, 2024. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA, USA.
Rubin, R.L., Van Groenigen, K.J. and Hungate, B.A., 2017. Plant growth promoting rhizobacteria are more effective under drought: a meta-analysis. Plant and Soil 416: 309–323.
Schuman, M.C. and Baldwin, I.T., 2016. The layers of plant responses to insect herbivores. Annual Review of Entomology 61: 373–394.
Selosse, M.-A., Bessis, A. and Pozo, M.J., 2014. Microbial priming of plant and animal immunity: symbionts as developmental signals. Trends in Microbiology 22: 607–613.
Shayanthan, A., Ordoñez, P.A.C. and Oresnik, I.J., 2022. The role of synthetic microbial communities (syncom) in sustainable agriculture. Frontiers in Agronomy 4: 58. https://doi.org/10.3389/fagro.2022.896307
Song, C., Zhu, F., Carrion, V.J. and Cordovez, V., 2020. Beyond plant microbiome composition: exploiting microbial functions and plant traits via integrated approaches. Frontiers in Bioengineering and Biotechnology 8: 896. https://doi.org/10.3389/fbioe.2020.00896
Trivedi, P., Leach, J.E., Tringe, S.G., Sa, T. and Singh, B.K., 2020. Plant-microbiome interactions: from community assembly to plant health. Nature Reviews Microbiology 18: 607–621. https://doi.org/10.1038/s41579-020-0412-1
Turlings, T.C. and Erb, M., 2018. Tritrophic interactions mediated by herbivore-induced plant volatiles: mechanisms, ecological relevance, and application potential. Annual Review of Entomology 63: 433–452.
Viechtbauer, W., 2010. Conducting meta-analyses in R with the metafor package. Journal of Statistical Software 36: 1–48.
Walling, L.L., 2000. The myriad plant responses to herbivores. Journal of Plant Growth Regulation 19: 195–216.
Wang, M., Ge, A.H., Ma, X., Wang, X., Xie, Q., Wang, L., Song, X., Jiang, M., Yang, W., Murray, J.D., Wang, Y., Liu, H., Cao, X. and Wang, E., 2024. Dynamic root microbiome sustains soybean productivity under unbalanced fertilization. Nature Communications 15: 1668. https://doi.org/10.1038/s41467-024-45925-5
Wu, J. and Baldwin, I.T., 2010. New insights into plant responses to the attack from insect herbivores. Annual Review of Genetics 44: 1–24.
Zeffa, D.M., Fantin, L.H., Koltun, A., de Oliveira, A.L., Nunes, M.P., Canteri, M.G. and Gonçalves, L.S., 2020. Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: a meta-analysis of studies from 1987 to 2018. PeerJ 8: e7905.
Zhang, Z., Li, Y., Williams, R.A., Chen, Y., Peng, R., Liu, X., Qi, Y. and Wang, Z., 2023. Responses of soil respiration and its sensitivities to temperature and precipitation: A meta-analysis. Ecological Informatics 75: 102057.
Zhao, X., Tian, P., Sun, Z., Liu, S., Wang, Q. and Zeng, Z., 2022. Rhizosphere effects on soil organic carbon processes in terrestrial ecosystems: A meta-analysis. Geoderma 412: 115739.
Züst, T. and Agrawal, A.A., 2016. Mechanisms and evolution of plant resistance to aphids. Nature Plants 2: 15206. https://doi.org/10.1038/nplants.2015.206





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}