Abstract
Coinfections where hosts harbour more than one parasite species are common in nature. Facilitation among parasites enabling them to better exploit host resources is widespread with direct consequences for their life-history. Plant viruses can facilitate their vectors which may increase their transmission, but which can also have positive effects on non-vector species that share the host plant. Here we study facilitation of two (non-vector) species of Tetranychus spider mites, Tetranychus urticae and T. evansi, by a plant virus, tomato spotted wilt virus (TSWV), in coinfections on a host tomato plant, Solanum lycopersicum. We compared the effect of different strains of TSWV on life-history traits of Tetranychus mites on both cut leaves and whole plants of different tomato varieties. TSWV facilitated both species of spider mites on two different tomato varieties on whole plants, with more offspring of both species becoming adults. In contrast, on cut leaves, facilitation of T. urticae was much more variable depending on the viral strain and tomato variety. We attribute this to the non-homogeneous spread of virus throughout the host plant, which may limit the potential for facilitation if it is plant mediated, either via the host immune response or resource based. Our experiments demonstrate that facilitation among parasites can shape life-history traits of non-vector species which may have knock-on effects driving epidemics of plant parasites and pests.
1 Introduction
Plants and animals are host to a diversity of parasites and often harbour coinfections (Dutt et al., 2022; Petney & Andrews, 1998). Coinfections are composed of different strains or species (e.g. of bacteria, protozoa, viruses, helminths, microsporidians) and can have both positive or negative effects on parasite life-history (Balmer et al., 2009; Mauck et al., 2010b; McQueen & McKenzie, 2006; Zilio & Koella, 2020) and epidemics (Norberg et al., 2023; Rodgers & Bolnick, 2023; Susi et al., 2015). Competition among parasites in coinfections is most commonly assumed, and has been well investigated both theoretically and empirically (reviewed in Alizon et al., 2013; Read and Taylor, 2001). For instance, Budischak et al. (2018) show that resource competition with hookworms can decrease Plasmodium culex density more than 2 fold in humans. In contrast, facilitation (positive) among parasites has been comparatively ignored despite being common (Dallas et al., 2019; Telfer et al., 2010; Zélé et al., 2018).
Theoretical studies show that facilitation among parasites can increase within-host growth (Eswarappa et al., 2012), transmission to new hosts (Alizon et al., 2013) and coinfection prevalence (Kamiya et al., 2018). These predictions have been corroborated by experimental studies showing facilitation in both plants and animals (reviewed in Zélé et al., 2018). Facilitation among parasites can occur via multiple mechanisms including the production of public goods aiding the growth of other parasites (Ford et al., 2016) or interactions mediated via the host immune system, such as negative immune crosstalk or immunosuppression preventing effective immune responses (Dagostin et al., 2023; Graham, 2008; Lello et al., 2018). For example, in sheep, suppression of the immune response by the nematode Haemonchus contortus facilitates the survival of the nematode Trichostrongylus colubriformis (Lello et al., 2018). Also, on tomato, coinfection with Tetranychus evansi can suppress plant immune responses increasing the number of eggs laid by Tetranychus urticae (Sarmento, Lemos, Dias, et al., 2011). Thus, facilitation can impact parasite life-history across different host and parasite systems in nature.
Plants are host to diverse parasites – fungi, bacteria, viruses and insects – and often harbour coinfections. Plant viruses notoriously infect a wide range of plant hosts, and the majority are transmitted by arthropod vectors (Nault, 1997). Virus-vector-plant relationships have been examined in a number of systems and it has been shown that plant viruses can alter the behaviour and phenotype of both their vectors and plant hosts (Fang et al., 2013; Mauck et al., 2010a; Pan et al., 2021) in seemingly adaptive ways to increase transmission (Blanc & Michalakis, 2016; Mauck et al., 2012). Virus infection can change plant physiology, by modifying the release of volatiles, visual cues and/or nutritional profile (Eigenbrode et al., 2002; Guo et al., 2019; Mauck et al., 2014; Pan et al., 2021). In turn, vectors are often more attracted to virus infected plants (Eigenbrode et al., 2002; Ingwell et al., 2012), and can have increased fitness following feeding on virus infected plants (Chen et al., 2013; Maluta et al., 2014; Maris et al., 2004) but see (Mauck et al., 2010a, 2010b, 2014). Viruses manipulating plant phenotypes can also impact other non-vector parasites sharing the host plant (Ángeles-López et al., 2018; Belliure et al., 2010; Mauck et al., 2010b, 2015). This includes positive effects of viruses on vector species being hijacked by other non-vector parasites sharing the plant (Ángeles-López et al., 2018; Belliure et al., 2010; Nachappa et al., 2013).
Plant hosts have evolved a suite of counterattack mechanisms involving physiological and immune adaptations against parasites (Glazebrook, 2005; Howe & Jander, 2008; Ross, 1961). The best studied are the jasmonic acid (JA) and salicylic acid (SA) plant immune pathways. Broadly, against necrotrophs and piercing- sucking herbivores, they employ the JA pathway, which can produce proteinase inhibitors (PIs) and polyphenol oxidase (PPO) (Vijayan et al., 1998; Wang & Wu, 2013) while against biotrophic pathogens like viruses, they employ the SA pathway which produces pathogenesis- related-genes and systemic acquired resistance (Klessig & Malamy, 1994; Raskin, 1992). The SA and JA immune pathways are often reciprocally antagonistic meaning it is not always possible to mount an effective immune response against diverse antagonists (Abe et al., 2012; Kunkel & Brooks, 2002; Thaler et al., 2002; Zarate et al., 2007). The beneficial effect of plant viruses on vector and non-vector fitness may occur via negative immune cross talk between the JA and SA pathways (Zhang et al., 2012). Plant viruses can activate the salicylic acid pathway (López-Gresa et al., 2016; Malamy et al., 1990; Singh et al., 2004) which can prevent activation of the JA pathway (Zhang et al., 2012). Another possibility is that facilitation is resource-based; viruses have been shown to increase the production of free amino acids in the plant host which may make it easier to feed upon (Blua et al., 1994; Nachappa et al., 2013; Shrestha et al., 2012).
In this study we investigate facilitation of non-vector plant ectoparasites, the spider mites, Tetranychus evansi and T. urticae, by tomato spotted wilt virus (TSWV) in coinfections. Interactions between Tetranychus spp. and tomato immune responses have been well studied (Kant et al., 2004; Sarmento, Lemos, Bleeker, et al., 2011). Both T. evansi and T. urticae are negatively affected by the JA pathway via the production of proteinase inhibitors (Ament et al., 2004; Ataide et al., 2016). However, strategies in both exist to overcome JA defences. T. evansi can suppress or reduce the induction of the JA pathway (Knegt et al., 2020; Sarmento, Lemos, Bleeker, et al., 2011; Teodoro-Paulo, Alba, et al., 2023). Although T. urticae often induces the JA pathway (Kant et al., 2004; Li et al., 2002; Rioja et al., 2017), it can be induced to relatively low levels (Teodoro-Paulo -preprint; Kant et al., 2008), and some strains or populations are tolerant to elevated JA (Kant et al., 2008).
The first aim of this study was to explore the generality of facilitation of spider mites by TSWV, to better understand whether it occurs for different TSWV strains, different Tetranychus species and on different varieties of tomato plant, as well as the range of life-history traits affected. Previous work has shown that TSWV increases the number of eggs laid by T. urticae on whole plants (Nachappa et al., 2013) and leaf discs (Belliure et al., 2010). However, T. urticae can experience intense competition during the juvenile period (Godinho et al., 2023) meaning that positive effects of TSWV on the number of eggs laid may not scale up to positively affect the number of offspring becoming adult (Belliure et al., 2010). The second aim was to explore whether facilitation of spider mites could be observed on both whole plants and on leaves removed from TSWV infected plants. Studies investigating plant parasite interactions often use leaves that have been removed from the plant for practical reasons (Belliure et al., 2010; Godinho et al., 2023). A recent study compared interactions between spider mites and their host on whole plants versus on leaves that had been cut from the plant showing that results were qualitatively similar (Dias et al., 2022). However, elevated T. urticae fecundity on TSWV infected leaf discs from pepper plants did not necessarily affect adult traits (Belliure et al., 2010). Differences in the degree or occurrence of facilitation on cut leaves versus whole plants may indicate that there is a gradient in virus induced changes in the plant, via negative immune cross talk and/or elevated free amino-acids, meaning facilitation might not be as strong on cut leaves. Indeed, free amino acid levels in TSWV infected plants vary both temporally and in different tissues (Catoni et al., 2009; Selman et al., 1961).
We first describe 2 experiments where we explore facilitation of T. urticae on tomato leaves cut from plants infected with TSWV. We then describe experiments on whole plants in which we show a positive effect of TSWV on T. evansi and T. urticae on 2 different varieties of tomato plant. We find that different strains of TSWV can positively impact T. urticae, that both T. evansi and T. urticae benefit from facilitation and that it occurs on different varieties of tomato plant.
2 Material and methods
2.1 Tomato spotted wilt virus
Tomato spotted wilt virus (TSWV) is a tripartite RNA plant virus from the Tospoviridae family with 2 ambisense RNA strands (Best, 1968; Van den Hurk et al., 1977). It is one of the most menacing plant viruses for agriculturists globally, and destroys ornamental plants, crops, and vegetables (Crosslin et al., 2009; Roselló et al., 1996). The symptoms include stunted growth in plants, leaf and stem necrosis, chlorotic spots and spoiled fruits (Roselló et al., 1996; Sherwood et al., 2003). In nature, TSWV is vectored by thrips, mainly, Frankiniella occidentalis (Ullman et al., 2002; Whitfield et al., 2015), but in our laboratory, we mechanically inoculate the host plants with a virus inoculum prepared by using leaves of an already infected plant. The infected leaf (1 g) is crushed with 4 mL of buffer (Na₂HPO₄.12H₂O 0.03 M + 0.2% DIECA) and 90 mg of charcoal in a pestle and mortar until it becomes a smooth paste. Then we add 75 mg of carborundum as a surface abrasive for the solution to be easily absorbed by the plant cells. This inoculum is rubbed gently with a finger on the surface of the 1st fully expanded leaf to coat the entire surface and rinsed after 20–25 minutes. The virus strains were obtained from the Plant Pathology unit, INRAE, Avignon and were maintained in the laboratory by serial passage of new plants every 14 days on 3-week-old tomato plants (variety Moneymaker). Plants were checked for infection using ELISA test kits by Agdia on a small piece of leaf removed from each plant. A small piece of leaf was also removed from all mock inoculated plants to keep treatments as similar as possible. TSWV infected and mock inoculated plants are maintained at 25 ± 2 °C with a 16: 8 light: dark cycle.
2.2 The spider mites
Tetranychus urticae, is a generalist mite species feeding on >1000 species of agricultural plants (Helle and Sabelis, 1985; Shih et al., 1976) while Tetranychus evansi are specialists on the Solanaceae family (Qureshi et al., 1969). Spider mites are haplo-diploid. Females lay eggs which hatch ~4 days later. The juvenile period includes 1 nymph stage and 2 deutonymph stages with adults emerging 13–14 days later. The populations of both species used in these experiments are outbred populations which originated and were created in Portugal (see Godinho et al. (2020) for more details). Subpopulations of each were transferred to Montpellier in January and November 2022, and have since been maintained on 3 Moneymaker tomato plants in big plastic boxes (dimension 520 mm × 300 mm × 250 mm) with one plant being changed every week. Populations are kept at 25 ± 2 °C on a 16: 8 light: dark cycle. Prior to each experiment, groups of 50 mated females were placed together on a cut tomato leaf to lay eggs to produce equally aged daughters for the experiments.
2.3 Host tomato plants
We used different commercially available varieties of host tomato plants, Solanum lycopersicum, in our experiments; Moneymaker, Saint-Pierre, Olympe and Micro Tom. Moneymaker and Olympe are F1 varieties. All plants were grown in an arthropod free environment at 25 °C ± 3 °C with a 16h:8h light: dark cycle.
2.4 Laboratory experiments
2.4.1 Effect of tomato variety on the interaction between TSWV and T. urticae on isolated tomato leaves removed from the plant
This experiment tested the effect of TSWV on Tetranychus urticae life-history traits on 3 different varieties of tomato, Moneymaker (MM), Saint-Pierre (SP), and Olympe (OL). Eight plants of each variety were infected with the France81 strain of TSWV 3-weeks post sowing. Eight control plants of each variety were also mock infected by applying a mock inocula (buffer with uninfected plant leaves) to the first fully expanded leaf. Infections were confirmed 14 days later by removing a small piece of leaf from infected plants, and testing with TSWV ImmunoStrips (from Agdia company, Paris, France). Small pieces of leaflets were also removed from control plants to control for any effect of removing leaves when testing for infection.
We cut the 2 leaves above the inoculated one and put both into the same transparent, plastic box (250 mm × 180 mm × 75 mm). Both leaves were pressed onto wet cotton wool, and the stem wrapped in wet cotton wool to keep them fresh. We placed adult mated T. urticae females in groups of 5 onto 2 leaflets on the first leaf and 1 leaflet on the second leaf. Note, spider mites were placed on the second leaf a day later. Both leaves from the same plant were placed in a box representing a single plant replicate with a total of 15 T. urticae (Figure 1). A barrier of lanolin and tanglefoot glue mixed in a 1:1 ratio was applied to the stem to isolate the mites on a single leaflet. The number of eggs laid by females were measured on the 4th day after transferring the mites to boxes at which point the mother females were also removed. On the 14th day after transferring spider mites, we counted the total number of adult male and female offspring on the leaflets. The adult count was continued for 2 more days to account for temporal variation in development. We also measured the size of the leaflets by quantifying the area from photos taken on day 12–13 using Image-J. This experiment aimed to have 3 tomato varieties × 2 parasite treatments (1 virus + mock control) × 8 replicates = 48 plants (see Table 1 for actual replicate numbers).
2.4.2 Effect of different TSWV strains on T. urticae fitness on isolated tomato leaves removed from the plant
Here we tested the effect of three different strains of TSWV (France81, LYE1137vir and LYE55) on T. urticae life-history traits on Moneymaker leaves. As in Experiment 1, we infected 8 3-week old plants post sowing and had 4 plants that were mock inoculated as before. In addition we also had 7 clean plants as controls to test whether wounding of the plant during mock inoculation had any effect on T. urticae life-history traits. As before, 2 leaves were taken from each plant and placed on water saturated cotton wool in the same box and 10 female spider mites placed on 1 leaflet of each leaf isolated with lanolin and Tanglefoot glue (Figure 1). We counted the number of eggs 4 days later (and removed the mothers), the number of emerging juveniles on the 8th day, and the number of male and female adult offspring on the 14th day. We quantified the leaflet areas by taking photos and analysing them with Image-J. See Table 1 for final replicate numbers.

Spider mite species, tomato plant varieties and viral strains used in each experiment (as described in Materials and Methods). The final number of replicates per treatment (numerator) over the aim number of replicates (denominator) are also shown. Note that final numbers were lower due to plants testing negative for TSWV.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10001
Pictorial representation of the experiment design on cut leaves and whole plants. A) spider mites transferred onto the cut leaves from the TSWV or mock infected plants used in experiments 1 and 2, while B) spider mites transferred onto the leaves of an intact plant used in experiments 3 and 4. In cut leaves, the spider mites were contained onto the leaflet with glue and tanglefoot as a barrier, while on the whole plants, we made muslin bags to isolate them onto the leaflet.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10001
2.4.3 Effects of TSWV on the number of adults produced by T. urticae and T. evansi on whole plants of different tomato varieties
In this experiment we tested the effect of TSWV on T. urticae and T. evansi life-history traits on whole tomato plants of the Moneymaker and Micro Tom varieties. We infected 16 plants of each variety 3 weeks post sowing with the France81 virus strain and mock inoculated another 16 as described above. After 14 days, we checked whether the plants were positive for virus infection as described above and placed the whole plants in groups of 8 in plastic boxes (dimension 520 mm × 300 mm × 250 mm).
We transferred adult female mites of either T. urticae or T. evansi in groups of 10 onto one leaflet on two different leaves above the inoculated leaf on each plant. Here the mites were isolated onto their respective leaflet using a muslin leaf cage (dimensions 70 mm × 90 mm × 10 mm). These cages were made by gluing muslin into a box shape and attaching a plastic ziplock from a freezer bag as an opening. The different treatments were randomised across boxes following the transfer of mites but mite species were kept in separate boxes due to contamination risks). We counted the number of adult male and female adults using a binocular microscope on the 14th day following removal of the infested leaves. Eggs or other developmental stages prior to day 14 were not measured as it was not possible to remove and replace the muslin bag without disrupting the mites. This experiment had 2 tomato varieties × 2 parasite treatments (1 virus + mock control) × 2 Tetranychus species × 8 replicates (see Table 1 for actual replicate numbers).
2.4.4 Effects of TSWV on female survival and the number of eggs and juveniles produced by T. urticae on whole plants
In a separate experiment, we tested whether TSWV infection influenced female survival and the number of eggs laid by T. urticae on Moneymaker plants. This was to be sure that differences in fitness on TSWV infected plants was not due to differential survival of mothers. We infected six plants with France81 and seven with LYE1137vir TSWV strains as described previously, and had six mock inoculated controls. After 14 days we checked the plants for positive infections and kept positive plants (2/6 for France81 and 7/7 for LYE1137vir) in plastic boxes (520 mm × 300 mm × 250 mm).
We transferred 10 adult female mites onto 1 leaflet each of 2 different leaves (on the 2 leaves above that which had been inoculated), and isolated them with the muslin leaf cages as described above. On the 4th day after transferring the mites, we took 3 plants that had been infected with LYE1137vir, 2 positively infected with France81, and 3 mock infected plants, and removed the leaves to count eggs; also, we counted the number of mother female mites surviving on the leaflet. These plants were culled and the number of juveniles on the 7 remaining plants (3 mock inoculated and 4 infected with LYE1137vir) were counted in the same way on day 8. This experiment had 3 parasite treatments (2 virus + mock control) × 6–7 replicates (see Table 1).
3 Statistical analyses
All analyses were done in R version (4.3.2) using the package glmmTMB. All models were simplified in a stepwise fashion removing non-significant terms. Full models are shown in Tables S1–S3 in the Supplementary materials.
We used Generalized Linear Mixed Models (GLMMs) to test whether the number of eggs laid, the total number of juveniles, total number of adult offspring, total number of adult daughters, and the offspring sex ratio were affected by plant variety, virus infection, mite species, leaflet area, and their interactions where relevant. As per the different experimental plans (Table 1), plant variety, mite species or virus infection were included in the models as fixed factors and leaflet area as a covariate.
GLMMs with a negative binomial distribution were implemented for each experiment for the number of eggs laid, the number of juveniles, adults and adult females to improve model fits as these count variables were overdispersed. Only in the experiment ‘Effects of TSWV on female survival and the number of eggs and juveniles produced by T. urticae on whole plants’ were the number of eggs laid and mother survival, analysed using a GLMM with Poisson error structure and log link function as this error distribution was a better fit. Offspring sex ratio was always analysed with a binomial error structure and logit link function with the number of adult males and the total number of females combined as a response variable using cbind. Post-hoc tests using the multiple comparisons in multcomp function were implemented for main effects with more than 2 levels and the emmeans function for interaction terms when significant. For the experiment ‘Effect of different TSWV strains on T. urticae fitness on isolated tomato leaves removed from the plant’ there was a significant interaction between virus infection and leaf area. To better understand what was driving this interaction we repeated the analysis for virus infected (France81, LYE1137vir and LYE55), control plants and mock infected treatments separately.
Individual plant replicate nested within viral infection was included in all models as a random factor for experiments on cut leaves removed from the plant and the experiment on whole plants measuring the number of eggs laid and survival. In Experiment 3, ‘Effect of tomato variety on facilitation of T. urticae and T. evansi by TSWV on whole plants’, individual plant replicates were nested within box ID, and box position on the shelf as a random factor. Leaflet number was included in some models as an observation level random effect to control for overdispersion (Harrison, 2014) (Tables S1 and S2).
4 Results
4.1.1 Effect of tomato variety on the interaction between TSWV and T. urticae on isolated tomato leaves removed from the plant
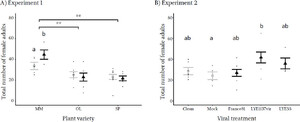
There was no effect of TSWV infection on the number of eggs laid by T. urticae (Table S1). The effect of TSWV infection on the number of adult daughters and total number of offspring depended on plant variety. Specifically, the number of adult daughters and total number of offspring were higher on virus infected plants than on control plants of the Moneymaker variety but not on those of the other varieties (Figure 2a), resulting in significant interactions between variety and virus infection ( = 11.06, p = 0.004 and = 6.88, p = 0.032, respectively, Table S1). There were also significant interactions between leaflet area and virus infection for the number of eggs laid (
= 11.06, p = 0.004 and = 6.88, p = 0.032, respectively, Table S1). There were also significant interactions between leaflet area and virus infection for the number of eggs laid ( = 6.28, p = 0.012), number of adult daughters ( = 8.09, p = 0.004; Figure S1) and number of adult offspring ( = 10.85, p < 0.001) showing a positive relationship among these traits on control, but not virus infected plants.
= 6.28, p = 0.012), number of adult daughters ( = 8.09, p = 0.004; Figure S1) and number of adult offspring ( = 10.85, p < 0.001) showing a positive relationship among these traits on control, but not virus infected plants.
Effect of tomato spotted wilt virus on mean (± standard error) total number of Tetranychus urticae adult female mites in A) Experiment 1, on mock (gray) and TSWV strain France81 infected (black) plants of Moneymaker (MM), Olympe (OL) and Saint-Pierre (SP) tomato varieties. ** represent the significant differences among the plant varieties, while “a” and “b” represent the significant difference between the mock and virus infected on the Moneymaker variety. B) Experiment 2, on mock and clean controls (gray) or plants infected with 3 different TSWV strains (France81, LYE1137vir, LYE55) (black) in Moneymaker plants. All the data points are jittered in black along the respective treatments.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10001
There was a significant effect of tomato variety showing a higher number of eggs laid by T. urticae ( = 11.99, p = 0.003), more adult offspring ( = 35.07, p < 0.001) and more adult daughters ( = 17.41, p < 0.001) on Moneymaker plants than the other two varieties (Figures 2a and S2a–b, Table S1). There was no effect of TSWV infection, tomato variety or leaflet area on the offspring sex ratio (Table S1, Figure S2c).
4.1.2 Effect of different TSWV strains on T. urticae fitness on isolated tomato leaves removed from the plant
There was a significant effect of infection with different viral strains on the number of adult daughters produced by T. urticae ( = 11.34, p = 0.023, Table S2). Specifically, plants infected with the LYE1137vir strain supported higher numbers of adult daughters than mock-infected plants (Figure 2b). The effect of virus infection on offspring sex ratio depended on the area of the leaflet they were feeding on (virus infection*leaflet area = 10.07, p = 0.039), with a more female biased offspring sex ratio on larger leaflets for TSWV infected plants, and a more male biased sex ratio on larger leaflets of clean plants, whereas there was no relationship between leaflet size and sex ratio in mock infected plants (Figure S3). There was no effect of viral strain, leaflet area or their interaction on the number of eggs laid, the number of juveniles or the total number of adults (Figure S4a–d, Table S2).
= 11.34, p = 0.023, Table S2). Specifically, plants infected with the LYE1137vir strain supported higher numbers of adult daughters than mock-infected plants (Figure 2b). The effect of virus infection on offspring sex ratio depended on the area of the leaflet they were feeding on (virus infection*leaflet area = 10.07, p = 0.039), with a more female biased offspring sex ratio on larger leaflets for TSWV infected plants, and a more male biased sex ratio on larger leaflets of clean plants, whereas there was no relationship between leaflet size and sex ratio in mock infected plants (Figure S3). There was no effect of viral strain, leaflet area or their interaction on the number of eggs laid, the number of juveniles or the total number of adults (Figure S4a–d, Table S2).
4.1.3 Effects of TSWV on the number of adults produced by T. urticae and T. evansi on whole plants of different tomato varieties
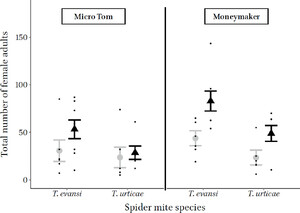
TSWV infection increased the number of offspring becoming adults ( = 16.29, p < 0.001) and the number of adult daughters ( = 14.85, p < 0.001). There were also more adult offspring ( = 4.22, p = 0.039, Fig. S5a) and adult daughters ( = 4.87, p = 0.027, Figure 3) on Moneymaker compared to Micro Tom plants (Table S3). Overall T. evansi had a higher total number of adult offspring ( = 5.38, p = 0.02) and more adult daughters ( = 10.68, p = 0.001) than T. urticae on both plant varieties (see Figure 3, Table S3). There were no significant interactions among TSWV infection, mite species and plant variety on the number of adult offspring or daughters (Table S3).
Effect of tomato spotted wilt virus on mean (± standard error) Tetranychus urticae and Tetranychus evansi number of adult daughters on mock (gray) and TSWV strain France81 infected (black) plants of the Micro Tom and Moneymaker varieties. All the data points are jittered in black along the respective treatments.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10001
The effect of TSWV infection on offspring sex ratio depended on plant variety (virus infection*plant variety; = 7.17, p = 0.007), with a less female biased sex ratio on virus infected plants of the Micro Tom variety, and mite species (virus infection*mite species; = 10.1, p = 0.001) with T. urticae having a less female biased offspring sex ratio on virus infected plants (see Figure S5b).
4.1.4 Effects of TSWV on female survival and the number of eggs and juveniles produced by T. urticae on whole plants
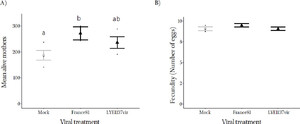
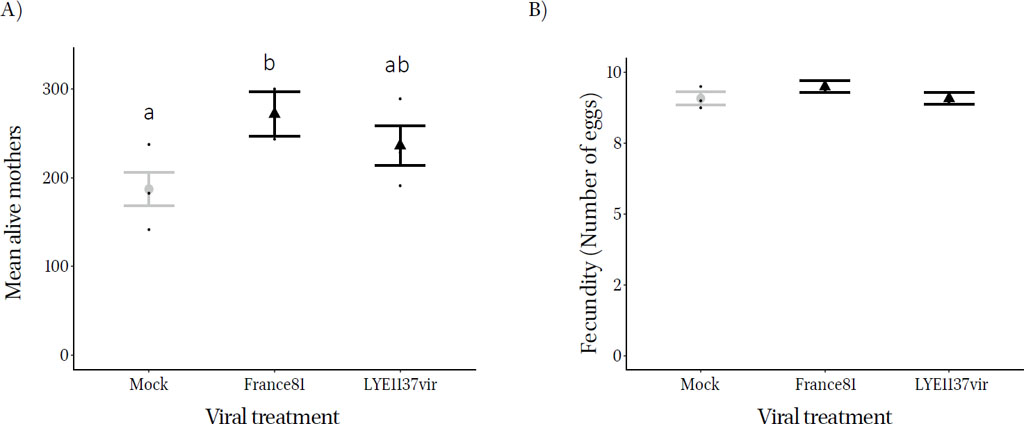
There was a positive effect of TSWV infection on the number of eggs laid by T. urticae ( = 6.35 p = 0.042) with more eggs being laid on France81 infected plants (Figure 4a). Similarly, the number of eggs that hatched, measured as the number of juveniles was higher in the presence of the virus ( = 7.31, p = 0.007; see Figure S6). There was no difference in the survival of mothers on virus infected or control plants ( = 0.06, p = 0.972) (Figure 4b; Table S3) meaning that there was no role of differential survival of mothers in facilitation.
Effect of tomato spotted wilt virus on mean (± standard error) T. urticae A) the number of eggs laid, B) mother survival on Moneymaker whole plants mock infected (gray) or infected with TSWV strains France81 and LYE1137vir (black). All the data points are jittered in black along the respective treatments.
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10001
5 Discussion
Our results confirm the facilitatory effect of tomato spotted wilt virus on both T. urticae and T. evansi on whole plants. This advances on previous studies (Belliure et al., 2010; Nachappa et al., 2013) showing that TSWV has a positive effect on the number of adult daughters and total number of adult offspring in addition to the number of eggs laid. Facilitation was repeatable for both T. evansi and T. urticae on two different tomato varieties on whole plants, Moneymaker and Micro Tom. In contrast, on cut leaves, facilitation was variable across experiments and depended upon the tomato variety. This may be due to the non-homogeneous spread of virus throughout the plant meaning the cut leaves used in the experiment were not always infected with TSWV or an indirect systemic effect mediated via virus infection was not effective.
5.1 Facilitation on whole plants
The mechanism of facilitation of T. urticae and T. evansi by TSWV might be due to different, non-mutually exclusive mechanisms. A number of studies show that the facilitation of arthropods (vectors and non-vectors) by plant virus infection arises via negative immune cross talk between the JA and SA immune pathways (Shi et al., 2014; Zhang et al., 2012). Prior induction of the SA pathway following infection with TSWV (López-Gresa et al., 2016) may prevent upregulation of the JA pathway and production of proteinase inhibitors which disrupt spider mite feeding and digestion of the plant material (Sarmento, Lemos, Bleeker, et al., 2011).
T. evansi have been shown to suppress the JA pathway or induce it to low levels (Knegt et al., 2020; Sarmento, Lemos, Bleeker, et al., 2011; Teodoro-Paulo et al., 2023). Similarly, populations of T. urticae can vary in the extent to which they induce the JA pathway (Kant et al., 2008). Both the T. evansi (Teodoro-Paulo et al., 2023) and T. urticae (Teodoro-Paulo, Fernandes, et al., 2023) populations used in this experiment induced the JA pathway slightly, to levels higher than in clean plants. It has been shown that even very low levels of JA can reduce the number of eggs laid by both species, with reductions being observed at lower levels for T. evansi than T. urticae (Ataide et al., 2016). If mediated via negative immune cross talk (Thaler et al., 2010; Zhang et al., 2012), facilitation may not be observed, or be reduced, for T. evansi lineages (e.g. Sarmento, 2011), or other parasites (Pan et al., 2013) that suppress JA responses.
Another, non-mutually exclusive hypothesis is that facilitation is via TSWV changing the nutritional quality of the plant. Plant viruses, including TSWV, can increase the levels of free-amino acids circulating in cell sap with positive effects on arthropod life-history traits (Ángeles-López et al., 2018; Blua et al., 1994; Nachappa et al., 2013). Thus, increased free-amino acids may be a source of nitrogen or essential amino acids for both Tetranychus species. Finally, the virus may be a source of nutrition for spider mites since they can ingest virus particles when feeding (Orlob, 1968). TSWV has a lipid envelope (Mohamed, 1981) with glycoproteins on the envelope surface (Tas et al., 1977) which could serve as a source of lipids and amino acids. It could be tested whether plant viruses can serve as a source of nutrition for spider mites (or other plant feeding arthropods) by feeding them on an artificial diet (as in Orlob, 1968), but also measuring life-history traits. A recent study found that Halteria ciliates can grow and divide feeding on viruses (DeLong et al., 2023) demonstrating that viruses can be a source of food. Thus, it remains possible that feeding on viruses is a more widespread phenomenon, maybe being common as a food source.
5.2 Variable facilitation on cut leaves
All plants that had been selected for inclusion in our experiments had tested positive for TSWV infection (for plants in virus treatments). However, despite plants being infected with TSWV, facilitation of spider mites was not always observed when measured on cut leaves. This may be for a number of reasons.
One possibility is the non-homogeneous spread of virus throughout the plant, and thus, leaves upon which facilitation was not observed may have been uninfected. Samuel, 1934 described how tobacco mosaic virus in potato plants reaches the roots ~ 4 days following infection, then the apical leaves (~5 days) and then gradually descends to leaves below and ascends from the roots (~10 days) with plants being completely infected 25 days later. Consistent with this we found that apical leaves were always infected 11 days following infection in plants that tested positive for TSWV, but that leaves immediately below were not always infected (see figure A1.1). More generally, following inoculation, many plant viruses spread from the epidermal cells via cell-to-cell movement through the plasmodesmata to the phloem in the sieve elements to be transported to the roots and other sink leaves (Hipper et al., 2013). Some plant viruses eventually spread throughout a plant and are not tissue restricted whereas others remain only in certain tissues (Hipper et al, Agrios, 2005).
The fact that leaves used in our experiments may not always have contained viral particles may have prevented facilitation from occurring. However, we did not think that removing leaves would necessarily prevent facilitation as plant mediated effects were maintained on cut leaves in other experiments with spider mites and other arthropods (Dias et al., 2022; Musser et al., 2002; Sarmento, Lemos, Dias, et al., 2011). For example, induction of plant immune responses on whole plants by T. urticae can later reduce the number of eggs laid by T. evansi on leaves or leaf discs that have been removed from the plant, including on leaves never previously in contact with mites (Dias et al., 2022; Sarmento, Lemos, Dias, et al., 2011). Although the mechanism of facilitation is linked to systemic responses, variation in expression of genes implicated in the JA and SA pathways can be observed locally in different regions of the same leaf (Betsuyaku et al., 2018; Schimmel et al., 2017). Furthermore, levels of free-amino acids may be higher in leaves infected with TSWV. Levels of free amino acids vary through time and in different tissues following TSWV infection (Catoni et al., 2009; Selman et al., 1961). Thus, facilitation of spider mites by TSWV may rely on local interactions. In line with this, it would be interesting to see if the degree of facilitation on whole plants depends on whether the leaf is infected with virus and viral quantity on whole plants. Other systems have found variation in effects of plant parasites/symbionts on arthropod fitness when measuring traits on whole plants versus leaf discs (Jaber and Vidal, 2010).
Another possibility is that the inconsistencies we observe between experiments on cut leaves versus whole plants may also be because the plant varieties compared in each were different. Indeed, on whole plants, the degree of the facilitation of T. urticae on Micro Tom was smaller than that on Moneymaker (Figure 3), although this difference in effect size was not significant (no tomato variety*mite species interaction). However, when comparing the France81 strain on the Moneymaker tomato variety, both factors that were included in all the experiments, there was always facilitation on whole plants, but not on cut leaves.
5.3 Sex allocation in response to virus infection
We find that virus infection can change the offspring sex ratio of spider mites feeding on these plants, but not always in the same direction. On intact Moneymaker plants, infection with TSWV strain France81 reduced the female sex ratio bias of T. urticae offspring, but on cut leaves it did not affect the sex ratio of T. urticae, and in T. evansi feeding on intact plants, this strain increased the female sex ratio. The reasons for these differences in sex allocation, the differential investment into male versus female offspring, for T. urticae on whole plant versus cut leaves, and between the two species, are not clear. One possibility is that the effect of TSWV is related to resource availability. In solitary parasitoids (Charnov et al., 1981) and sawflies (Craig et al., 1992), allocation to female offspring is higher in good resource environments attributed to greater fitness returns from larger daughters and/or benefitting more from good environments. Although female spider mites are larger than males, it is not clear whether relative fitness returns are greater from larger daughters (Macke et al., 2010). Spider mites are haplodiploid, females developing from fertilised eggs and males from unfertilised eggs. Previous work has shown that one way female T. urticae adjust their sex allocation is through allocation of resources to eggs, with larger eggs more likely to encounter sperm, be fertilised and become female (Macke et al., 2010). Further, females can facultatively adjust egg size and sex allocation adaptively in response to environmental cues (Macke et al., 2012). Thus, fitness returns may arise due to the optimal offspring sex ratio which, in spider mites, can result from higher allocation of resources to daughters under certain conditions (Macke et al., 2011, 2014) which may also be related to resources available in different environments. If TSWV improves the plant as a resource for spider mites under certain conditions this may in turn increase sex allocation to the production of daughters.
As females are the dispersing stage it has previously been postulated that T. urticae produce a more female biased offspring sex ratio under poor conditions to escape (Young et al., 1986). Density dependent dispersal has been shown for many species (Dorken and Pannell, 2008; Matthysen, 2005). Recently Chokechaipaisarn and Gardner (2022) theoretically showed that more female biased offspring sex-ratios are favored under density-dependent dispersal in viscous populations, like spider mites. Thus, it is also possible that females may be responding to signals indicating that deterioration of the plant environment is imminent.
5.4 Virus interactions with non-vector species
Virus-vector interactions are indeed embedded within complex communities and better understanding of viral transmission may ultimately depend on knowledge about interactions at broader scales (Crowder et al., 2019; Eigenbrode et al., 2018). Viruses positively impacting the fitness of their vectors is adaptive if it increases transmission to new hosts (Blanc and Michalakis, 2016). However, it is not clear whether simultaneous facilitation of other non-vector parasites or arthropod herbivores sharing a host is costly which may in turn counter any beneficial effects on vectors. We are only aware of one study exploring the tripartite interaction between T. urticae, TSWV and their thrips vector and this study focussed on the positive effects of TSWV and thrips feeding on T. urticae life-history traits (Belliure et al., 2010). In the absence of virus, it has been shown that thrips predate upon Tetranychus species (Agrawal et al., 1999) with a higher proportion of Frankliniella occidentalis thrips larvae becoming adult when feeding on T. urticae eggs but a lower proportion becoming adult when feeding on T. evansi eggs (Ataide et al., 2019). Further, upregulation of the plant immune system by T. turkestani can negatively affect thrips survival in the absence of predation (Agrawal et al., 1999). These studies (as well as others) show that spider mites have the potential to impact thrips life-history via multiple mechanisms, notably, predation, immune mediated effects on the plant (Agrawal et al., 1999; Ataide et al., 2019), spider mite web protecting thrips from predation (Pallini et al., 1998) and possibly competition, but thrips were found to be the superior competitor (Zhi et al., 2006). Thus, overall it would seem that the presence of Tetranychus species positively impacts thrips life-history, and so would not necessarily affect TSWV transmission. Further work is needed to understand the direct effects of spider mites on TSWV growth and the potential for indirect effects via the thrips vector.
Other studies have explored how vector – non-vector interactions may impact viral transmission. One study found that feeding by a non-vector species displaced an aphid vector to the most susceptible region of the plant for Pea enation mosaic virus (PEMV) transmission (Chisholm et al, 2019), and that the presence of these non-vector weevils increased PEMV titre (Chisholm et al., 2018). Su et al. (2016) found that non-vector T. urticae negatively affects whitefly vector life-history traits on tomato hosts both in the presence and absence of Tomato yellow leaf curl virus (TYLCV). However, a more recent study by the same authors found that T. urticae actually increases TYLCV viral titre and also transmission by increasing whitefly feeding rate (Su et al., 2020). These studies suggest that plant viruses can under certain circumstances actually benefit from interactions between their vectors and non-vector species.
Greater knowledge about how virus – vector – non- vector interactions affect virus (or parasite) transmission and vector/non-vector life-history could aid control programs. In plant systems, vectors, non-vectors and viruses/parasites are often all pest species. It may be the case that one of the players disproportionately augments life-history traits and/or spread of others in the system. In such a scenario, targeting that species may have beneficial knock-on effects for the control of other pest species.
5.5 More general evolutionary consequences of facilitation
Facilitation can be an important process shaping the ecology and evolution of both free-living and parasitic species (Bronstein, 2009; McIntire and Fajardo, 2014; Zélé et al., 2018). However, the potential for facilitation to persist may vary across environments and how evolution of facilitation proceeds between different species may depend on the consequences of facilitation for the benefactor (Bronstein, 2009) which may vary across environments. For example, facilitation is often predicted to be more prevalent under stressful conditions when competitive interactions are reduced (He et al., 2013; Hesse et al., 2021). Further, coevolution between the protective bacteria Enterococcus faecalis and Staphyloccucus aureus in coinfection in a nematode host selected against siderophore (iron scavenging molecules that are a public good) production by S. aureus; siderophores facilitate E. faecalis growth with negative effects on S. aureus (Ford et al., 2016). This highlights that facilitation can be selected against when pairwise interactions are at play. It may though be the case that scaling up to multiple interactions in a community may be more likely to permit niche differentiation and/or indirect interactions among players to maintain facilitative interactions.
6 Conclusion
We find that TSWV can facilitate life-history traits of both T. urticae and T. evansi on whole plants. The reciprocal effect of spider mites on TSWV either via the plant or vector remains to be shown. Nevertheless, this work highlights that facilitation among parasites may be an important factor affecting parasite severity and spread. This work shows the importance of garnering a better understanding of parasite life-history within a multiple-parasite/community context.
Notes and Acknowledgements
Data availability
Data is available on the landing page of this article at brill.com/pami under Supplementary Materials and on https://zenodo.org/records/14939003.
Statement of conflict
The authors declare no conflict of interest.
Funding
This work was funded by an ANR grant (EVOLVIR: ANR-20-CE35-0013) to A. B. D.
Acknowledgements
We most sincerely thank Benoit Moury and Marion Szadkowski from Plant Pathology unit, INRAE, Avignon, for providing our laboratory with the TSWV strains, and their helpful feedback about the experiments and virus pathology. We would also like to thank Yannis Michalakis for the insightful discussions about the results, and the project. We would like to thank Giacomo Zilio for helpful comments on a previous version of this manuscript. This is publication number ISEM-2025-028.
Bibliography
Abe, H., Tomitaka, Y., Shimoda, T., Seo, S., Sakurai, T., Kugimiya, S., Tsuda, S., & Kobayashi, M. (2012). Antagonistic plant defense system regulated by phytohormones assists interactions among vector insect, thrips and a Tospovirus. Plant and Cell Physiology, 53(1), 204–212. https://doi.org/10.1093/pcp/pcr173
Agrawal, A. A., Kobayashi, C., & Thaler, J. S. (1999). Influence of prey availability and induced host-plant resistance on omnivory by western flower thrips. Ecology, 80(2), 518–523. https://doi.org/10.1890/0012-9658(1999)080[0518:IOPAAI]2.0.CO;2
Alizon, S., de Roode, J. C., & Michalakis, Y. (2013). Multiple infections and the evolution of virulence. Ecol. Lett., 16(4), 556–567. https://doi.org/10.1111/ele.12076
Ament, K., Kant, M. R., Sabelis, M. W., Haring, M. A., & Schuurink, R. C. (2004). Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato. Plant Physiol., 135(4), 2025–2037. https://doi.org/10.1104/pp.104.048694
Ángeles-López, Y. I., Rivera-Bustamante, R., & Heil, M. (2018). Fatal attraction of non-vector impairs fitness of manipulating plant virus. J. Ecol., 106(1), 391–400. https://doi.org/10.1111/1365-2745.12838
Ataide, L. M. S., Dias, C. R., Schimmel, B. C. J., van Erp, T., Pallini, A., & Kant, M. R. (2019). Food decisions of an omnivorous thrips are independent from the indirect effects of jasmonate-inducible plant defences on prey quality. Scientific Reports, 9(1), 1727. https://doi.org/10.1038/s41598-018-38463-w
Ataide, L. M. S., Pappas, M. L., Schimmel, B. C. J., Lopez-Orenes, A., Alba, J. M., Duarte, M. V. A., Pallini, A., Schuurink, R. C., & Kant, M. R. (2016). Induced plant-defenses suppress herbivore reproduction but also constrain predation of their offspring. Plant Sci., 252, 300–310. https://doi.org/10.1016/j.plantsci.2016.08.004
Balmer, O., Stearns, S. C., Schötzau, A., & Brun, R. (2009). Intraspecific competition between co-infecting parasite strains enhances host survival in African trypanosomes. Ecology, 90(12), 3367–3378. https://doi.org/10.1890/08-2291.1
Belliure, B., Sabelis, M. W., & Janssen, A. (2010). Vector and virus induce plant responses that benefit a non-vector herbivore. Basic and Applied Ecology, 11(2), 162–169. https://doi.org/10.1016/j.baae.2009.09.004
Best, R. J. (1968). Tomato spotted wilt virus. Adv. Virus Res., 13, 65–146. https://doi.org/10.1016/s0065-3527(08)60251-1
Betsuyaku, S., Katou, S., Takebayashi, Y., Sakakibara, H., Nomura, N., & Fukuda, H. (2018). Salicylic acid and jasmonic acid pathways are activated in spatially different domains around the infection site during effector-triggered immunity in Arabidopsis thaliana. Plant and Cell Physiology, 59(1), 8–16. https://doi.org/10.1093/pcp/pcx181
Blanc, S., & Michalakis, Y. (2016). Manipulation of hosts and vectors by plant viruses and impact of the environment. Current Opinion in Insect Science, 16, 36–43. https://doi.org/10.1016/j.cois.2016.05.007
Blua, M. J., Perring, T. M., & Madore, M. A. (1994). Plant virus-induced changes in aphid population development and temporal fluctuations in plant nutrients. J. Chem. Ecol., 20(3), 691–707. https://doi.org/10.1007/BF02059607
Bronstein, J. L. (2009). The evolution of facilitation and mutualism. Journal of Ecology, 97(6), 1160–1170. https://doi.org/10.1111/j.1365-2745.2009.01566.x
Budischak, S. A., Wiria, A. E., Hamid, F., Wammes, L. J., Kaisar, M. M. M., van Lieshout, L., Sartono, E., Supali, T., Yazdanbakhsh, M., & Graham, A. L. (2018). Competing for blood: The ecology of parasite resource competition in human malaria-helminth co-infections. Ecol. Lett., 21(4), 536–545. https://doi.org/10.1111/ele.12919
Catoni, M., Miozzi, L., Fiorilli, V., Lanfranco, L., & Accotto, G. P. (2009). Comparative analysis of expression profiles in shoots and roots of tomato systemically infected by tomato spotted wilt virus reveals organ-specific transcriptional responses. Molecular Plant-Microbe Interactions®, 22(12), 1504–1513. https://doi.org/10.1094/MPMI-22-12-1504
Charnov, E. L., Los-den Hartogh, R. L., Jones, W. T., & van den Assem, J. (1981). Sex ratio evolution in a variable environment. Nature, 289(5793), 27–33. https://doi.org/10.1038/289027a0
Chen, G., Pan, H., Xie, W., Wang, S., Wu, Q., Fang, Y., Shi, X., & Zhang, Y. (2013). Virus infection of a weed increases vector attraction to and vector fitness on the weed. Scientific Reports, 3(1), 2253. https://doi.org/10.1038/srep02253
Chisholm, P. J., Sertsuvalkul, N., Casteel, C. L., & Crowder, D. W. (2018). Reciprocal plant-mediated interactions between a virus and a non-vector herbivore. Ecology, 99(10), 2139–2144. https://doi.org/10.1002/ecy.2449
Chokechaipaisarn, C., & Gardner, A. (2022). Density-dependent dispersal promotes female-biased sex allocation in viscous populations. Biology Letters, 18(8), 20220205. https://doi.org/10.1098/rsbl.2022.0205
Craig, T. P., Price, P. W., & Itami, J. K. (1992). Facultative sex ratio shifts by a herbivorous insect in response to variation in host plant quality. Oecologia, 92(2), 153–161. https://doi.org/10.1007/BF00317357
Crosslin, J. M., Mallik, I., & Gudmestad, N. C. (2009). First report of tomato spotted wilt virus causing potato tuber necrosis in Texas. Plant Dis., 93(8), 845. https://doi.org/10.1094/PDIS-93-8-0845A
Crowder, D. W., Li, J., Borer, E. T., Finke, D. L., Sharon, R., Pattemore, D. E., & Medlock, J. (2019). Species interactions affect the spread of vector-borne plant pathogens independent of transmission mode. Ecology, 100(9), e02782. https://doi.org/10.1002/ecy.2782
Dagostin, F., Vanalli, C., Boag, B., Casagrandi, R., Gatto, M., Mari, L., & Cattadori, I. M. (2023). The enemy of my enemy is my friend: Immune-mediated facilitation contributes to fitness of co-infecting helminths. J. Anim. Ecol., 92(2), 477–491. https://doi.org/10.1111/1365-2656.13863
Dallas, T. A., Laine, A.-L., & Ovaskainen, O. (2019). Detecting parasite associations within multi-species host and parasite communities. Proc. Biol. Sci., 286(1912), 20191109. https://doi.org/10.1098/rspb.2019.1109
DeLong, J. P., Van Etten, J. L., Al-Ameeli, Z., Agarkova, I. V., & Dunigan, D. D. (2023). The consumption of viruses returns energy to food chains. Proceedings of the National Academy of Sciences, 120(1), e2215000120. https://doi.org/10.1073/pnas.2215000120
Dias, C. R., Ataíde, L. M. S., Meijer, T. T., Venzon, M., Pallini, A., & Janssen, A. (2022). Phytophagous mite performance on intact plants and leaf discs with different defence levels. Entomologia Experimentalis et Applicata, 170(11), 941–947. https://doi.org/10.1111/eea.13227
Dorken, M. E., & Pannell, J. R. (2008). Density‐dependent regulation of the sex ratio in an annual plant. The American Naturalist, 171(6), 824–830. https://doi.org/10.1086/587524
Dutt, A., Andrivon, D., & Le May, C. (2022). Multi- infections, competitive interactions, and pathogen coexistence. Plant Pathology, 71(1), 5–22. https://doi.org/10.1111/ppa.13469
Eigenbrode, S. D., Bosque-Pérez, N. A., & Davis, T. S. (2018). Insect-borne plant pathogens and their vectors: ecology, evolution, and complex interactions. Annual Review of Entomology, 63(Volume 63, 2018), 169–191. https://doi.org/10.1146/annurev-ento-020117-043119
Eigenbrode, S. D., Ding, H., Shiel, P., & Berger, P. H. (2002). Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae). Proc. Biol. Sci., 269(1490), 455–460. https://doi.org/10.1098/rspb.2001.1909
Eswarappa, S. M., Estrela, S., & Brown, S. P. (2012). Within-host dynamics of multi-species infections: facilitation, competition and virulence. PLoS One, 7(6), e38730. https://doi.org/10.1371/journal.pone.0038730
Fang, Y., Jiao, X., Xie, W., Wang, S., Wu, Q., Shi, X., Chen, G., Su, Q., Yang, X., Pan, H., & Zhang, Y. (2013). Tomato yellow leaf curl virus alters the host preferences of its vector Bemisia tabaci. Sci. Rep., 3, 2876. https://doi.org/10.1038/srep02876
Ford, S. A., Kao, D., Williams, D., & King, K. C. (2016). Microbe- mediated host defence drives the evolution of reduced pathogen virulence. Nature Communications, 7(1), 13430. https://doi.org/10.1038/ncomms13430
Glazebrook, J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol., 43, 205–227. https://doi.org/10.1146/annurev.phyto.43.040204.135923
Godinho, D. P., Cruz, M. A., Charlery de la Masselière, M., Teodoro-Paulo, J., Eira, C., Fragata, I., Rodrigues, L. R., Zélé, F., & Magalhães, S. (2020). Creating outbred and inbred populations in haplodiploids to measure adaptive responses in the laboratory. Ecol. Evol., 10(14), 7291–7305. https://doi.org/10.1002/ece3.6454
Godinho, D. P., Rodrigues, L. R., Lefèvre, S., Delteil, L., Mira, A. F., Fragata, I. R., Magalhães, S., & Duncan, A. B. (2023). Limited host availability disrupts the genetic correlation between virulence and transmission. Evol Lett, 7(1), 58–66. https://doi.org/10.1093/evlett/qrac008
Graham, A. L. (2008). Ecological rules governing helminth- microparasite coinfection. Proc. Natl. Acad. Sci. U. S. A., 105(2), 566–570. https://doi.org/10.1073/pnas.0707221105
Griffiths, E. C., Pedersen, A. B., Fenton, A., & Petchey, O. L. (2014). Analysis of a summary network of co-infection in humans reveals that parasites interact most via shared resources. Proc. Biol. Sci., 281(1782), 20132286. https://doi.org/10.1098/rspb.2013.2286
Guo, L., Su, Q., Yang, Z., Xie, W., Wang, S., Wu, Q., & Zhang, Y. (2019). Amino acid utilization may explain why Bemisia tabaci Q and B differ in their performance on plants infected by the tomato yellow leaf curl virus. Front. Physiol., 10, 430111. https://doi.org/10.3389/fphys.2019.00489
Harrison, X. A. (2014). Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ, 2, e616. https://doi.org/10.7717/peerj.616
He, Q., Bertness, M. D., & Altieri, A. H. (2013). Global shifts towards positive species interactions with increasing environmental stress. Ecology Letters, 16(5), 695–706. https://doi.org/10.1111/ele.12080
Helle, W., & Sabelis, M. W. (1985). Spider mites: their biology, natural enemies and control (Vol. 1). Elsevier Amsterdam.
Hesse, E., O’Brien, S., Luján, A. M., Sanders, D., Bayer, F., van Veen, E. M., Hodgson, D. J., & Buckling, A. (2021). Stress causes interspecific facilitation within a compost community. Ecology Letters, 24(10), 2169–2177. https://doi.org/10.1111/ele.13847
Hipper, C., Brault, V., Ziegler-Graff, V., & Revers, F. (2013). Viral and cellular factors involved in phloem transport of plant viruses. Frontiers in Plant Science, 4. https://doi.org/10.3389/fpls.2013.00154
Howe, G. A., & Jander, G. (2008). Plant immunity to insect herbivores. Annual Review of Plant Biology, 59 (Volume 59, 2008), 41–66. https://doi.org/10.1146/annurev.arplant.59.032607.092825
Ingwell, L. L., Eigenbrode, S. D., & Bosque-Pérez, N. A. (2012). Plant viruses alter insect behavior to enhance their spread. Sci. Rep., 2, 578. https://doi.org/10.1038/srep00578
Jaber, L. R., & Vidal, S. (2010). Fungal endophyte negative effects on herbivory are enhanced on intact plants and maintained in a subsequent generation. Ecological Entomology, 35(1), 25–36. https://doi.org/10.1111/j.1365-2311.2009.01152.x
Kamiya, T., Mideo, N., & Alizon, S. (2018). Coevolution of virulence and immunosuppression in multiple infections. Journal of Evolutionary Biology, 31(7), 995–1005. https://doi.org/10.1111/jeb.13280
Kant, M. R., Ament, K., Sabelis, M. W., Haring, M. A., & Schuurink, R. C. (2004). Differential timing of spider mite- induced direct and indirect defenses in tomato plants. Plant Physiol., 135(1), 483–495. https://doi.org/10.1104/pp.103.038315
Kant, M. R., Sabelis, M. W., Haring, M. A., & Schuurink, R. C. (2008). Intraspecific variation in a generalist herbivore accounts for differential induction and impact of host plant defences. Proc. Biol. Sci., 275(1633), 443–452. https://doi.org/10.1098/rspb.2007.1277
Klessig, D. F., & Malamy, J. (1994). The salicylic acid signal in plants. Plant Mol. Biol., 26(5), 1439–1458. https://doi.org/10.1007/BF00016484
Knegt, B., Meijer, T. T., Kant, M. R., Kiers, E. T., & Egas, M. (2020). Spider mite populations suppress tomato defenses to varying degrees. Ecol. Evol., 10(10), 4375–4390. https://doi.org/10.1002/ece3.6204
Kunkel, B. N., & Brooks, D. M. (2002). Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol., 5(4), 325–331. https://doi.org/10.1016/s1369-5266(02)00275-3
Lello, J., McClure, S. J., Tyrrell, K., & Viney, M. E. (2018). Predicting the effects of parasite co-infection across species boundaries. Proceedings of the Royal Society B: Biological Sciences. https://doi.org/10.1098/rspb.2017.2610
Li, C., Williams, M. M., Loh, Y.-T., Lee, G. I., & Howe, G. A. (2002). Resistance of cultivated tomato to cell content-feeding herbivores is regulated by the octadecanoid-signaling pathway. Plant Physiol., 130(1), 494–503. https://doi.org/10.1104/pp.005314
López-Gresa, M. P., Lisón, P., Yenush, L., Conejero, V., Rodrigo, I., & Bellés, J. M. (2016). Salicylic acid is involved in the basal resistance of tomato plants to citrus exocortis viroid and tomato spotted wilt virus. PLOS ONE, 11(11), e0166938. https://doi.org/10.1371/journal.pone.0166938
Macke, E., Magalhães, S., Bach, F., & Olivieri, I. (2011). Experimental evolution of reduced sex ratio adjustment under local mate competition. Science, 334(1), 1127–1129. https://doi.org/10.1126/science.1212177
Macke, E., Magalhães, S., Bach, F., & Olivieri, I. (2012). Sex-ratio adjustment in response to local mate competition is achieved through an alteration of egg size in a haplodiploid spider mite. Proceedings of the Royal Society B: Biological Sciences, 279(1747), 4634–4642. https://doi.org/10.1098/rspb.2012.1598
Macke, E., Magalhães, S., Khan, H. D.-T., Luciano, A., Frantz, A., Facon, B., & Olivieri, I. (2010). Sex allocation in haplodiploids is mediated by egg size: evidence in the spider mite Tetranychus urticae Koch. Proceedings of the Royal Society B: Biological Sciences, 278(1708), 1054–1063. https://doi.org/10.1098/rspb.2010.1706
Macke, E., Olivieri, I., & Magalhães, S. (2014). Local mate competition mediates sexual conflict over sex ratio in a haplodiploid spider mite. Current Biology, 24(23), 2850–2854. https://doi.org/10.1016/j.cub.2014.10.040
Malamy, J., Carr, J. P., Klessig, D. F., & Raskin, I. (1990). Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science, 250(4983), 1002– 1004. https://doi.org/10.1126/science.250.4983.1002
Maluta, N. K. P., Garzo, E., Moreno, A., Lopes, J. R. S., & Fereres, A. (2014). Tomato yellow leaf curl virus benefits population growth of the Q biotype of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Neotrop. Entomol., 43(4), 385–392. https://doi.org/10.1007/s13744-014-0223-z
Maris, P. C., Joosten, N. N., Goldbach, R. W., & Peters, D. (2004). Tomato spotted wilt virus infection improves host suitability for its vector Frankliniella occidentalis. Phytopathology, 94(7), 706–711. https://doi.org/10.1094/PHYTO.2004.94.7.706
Matthysen, E. (2005). Density-dependent dispersal in birds and mammals. Ecography, 28(3), 403–416. https://doi.org/10.1111/j.0906-7590.2005.04073.x
Mauck, K., Bosque-Pérez, N. A., Eigenbrode, S. D., De Moraes, C. M., & Mescher, M. C. (2012). Transmission mechanisms shape pathogen effects on host–vector interactions: Evidence from plant viruses. Funct. Ecol., 26(5), 1162–1175. https://doi.org/10.1111/j.1365-2435.2012.02026.x
Mauck, K. E., De Moraes, C. M., & Mescher, M. C. (2010a). Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proceedings of the National Academy of Sciences, 107(8), 3600–3605. https://doi.org/10.1073/pnas.0907191107
Mauck, K. E., De Moraes, C. M., & Mescher, M. C. (2010b). Effects of cucumber mosaic virus infection on vector and non-vector herbivores of squash. Commun. Integr. Biol., 3(6), 579–582. https://doi.org/10.4161/cib.3.6.13094
Mauck, K. E., De Moraes, C. M., & Mescher, M. C. (2014). Biochemical and physiological mechanisms underlying effects of cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant Cell Environ., 37(6), 1427–1439. https://doi.org/10.1111/pce.12249
Mauck, K. E., Smyers, E., De Moraes, C. M., & Mescher, M. C. (2015). Virus infection influences host plant interactions with non-vector herbivores and predators. Funct. Ecol., 29(5), 662–673. https://doi.org/10.1111/1365-2435.12371
McIntire, E. J. B., & Fajardo, A. (2014). Facilitation as a ubiquitous driver of biodiversity. New Phytol., 201(2), 403–416. https://doi.org/10.1111/nph.12478
McQueen, P. G., & McKenzie, F. E. (2006). Competition for red blood cells can enhance Plasmodium vivax parasitemia in mixed-species malaria infections. Am. J. Trop. Med. Hyg., 75(1), 112–125. https://www.ncbi.nlm.nih.gov/pubmed/16837717
Mohamed, N. A. (1981). Isolation and characterization of subviral structures from tomato spotted wilt virus. Journal of General Virology, 53(2), 197–206. https://doi.org/10.1099/0022-1317-53-2-197
Musser, R. O., Hum-Musser, S. M., Eichenseer, H., Peiffer, M., Ervin, G., Murphy, J. B., & Felton, G. W. (2002). Caterpillar saliva beats plant defences. Nature, 416(6881), 599–600. https://doi.org/10.1038/416599a
Nachappa, P., Margolies, D. C., Nechols, J. R., Whitfield, A. E., & Rotenberg, D. (2013). Tomato spotted wilt virus benefits a non-vector arthropod, Tetranychus urticae, by modulating different plant responses in tomato. PLoS One, 8(9), e75909. https://doi.org/10.1371/journal.pone.0075909
Nault, L. R. (1997). Arthropod transmission of plant viruses: a new synthesis. Annals of the Entomological Society of America, 90(5), 521–541. https://doi.org/10.1093/aesa/90.5.521
Norberg, A., Susi, H., Sallinen, S., Baran, P., Clark, N. J., & Laine, A.-L. (2023). Direct and indirect viral associations predict coexistence in wild plant virus communities. Current Biology, 33(9), 1665–1676.e4. https://doi.org/10.1016/j.cub.2023.03.022
Orlob, G. B. (1968). Relationships between Tetranychus urticae Koch and some plant viruses. Virology, 35(1), 121–133. https://doi.org/10.1016/0042-6822(68)90312-7
Pallini, Janssen, & Sabelis. (1998). Predators induce interspecific herbivore competition for food in refuge space. Ecology Letters, 1(3), 171–177. https://doi.org/10.1046/j.1461-0248.1998.00019.x
Pan, H., Chen, G., Li, F., Wu, Q., Wang, S., Xie, W., Liu, B., Xu, B., & Zhang, Y. (2013). Tomato spotted wilt virus infection reduces the fitness of a nonvector herbivore on pepper. J. Econ. Entomol., 106(2), 924–928. https://doi.org/10.1603/ec12365
Pan, L.-L., Miao, H., Wang, Q., Walling, L. L., & Liu, S.-S. (2021). Virus-induced phytohormone dynamics and their effects on plant–insect interactions. New Phytol., 230(4), 1305–1320. https://doi.org/10.1111/nph.17261
Petney, T. N., & Andrews, R. H. (1998). Multiparasite communities in animals and humans: Frequency, structure and pathogenic significance. Int. J. Parasitol., 28(3), 377–393. https://doi.org/10.1016/s0020-7519(97)00189-6
Qureshi, A. H., Oatman, E. R., & Fleschner, C. A. (1969). Biology of the spider mite, Tetranychus evansi1,2. Ann. Entomol. Soc. Am., 62(4), 898–903. https://doi.org/10.1093/aesa/62.4.898
Ramiro, R. S., Pollitt, L. C., Mideo, N., & Reece, S. E. (2016). Facilitation through altered resource availability in a mixed-species rodent malaria infection. Ecol. Lett., 19(9), 1041–1050. https://doi.org/10.1111/ele.12639
Raskin, I. (1992). Role of salicylic acid in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 43(1), 439–463. https://doi.org/10.1146/annurev.pp.43.060192.002255
Read, A. F., & Taylor, L. H. (2001). The ecology of genetically diverse infections. Science, 292(5519), 1099–1102. https://doi.org/10.1126/science.1059410
Rioja, C., Zhurov, V., Bruinsma, K., Grbic, M., & Grbic, V. (2017). Plant-herbivore interactions: a case of an extreme generalist, the two-spotted spider mite Tetranychus urticae. Mol. Plant. Microbe. Interact., 30(12), 935–945. https://doi.org/10.1094/MPMI-07-17-0168-CR
Rodgers, M. L., & Bolnick, D. I. (2023). Opening a can of worms: a test of the co-infection facilitation hypothesis. Oecologia, 1–9. https://doi.org/10.1007/s00442-023-05409-7
Roselló, S., Díez, M. J., & Nuez, F. (1996). Viral diseases causing the greatest economic losses to the tomato crop. I. The tomato spotted wilt virus – a review. Scientia Horticulturae, 67(3), 117–150. https://doi.org/10.1016/S0304-4238(96)00946-6
Ross, A. F. (1961). Systemic acquired resistance induced by localized virus infections in plants. Virology, 14(3), 340–358. https://doi.org/10.1016/0042-6822(61)90319-1
Samuel, G. (1934). The movement of tobacco mosaic virus within the plant. Annals of Applied Biology, 21(1), 90–111. https://doi.org/10.1111/j.1744-7348.1934.tb06891.x
Sarmento, R. A., Lemos, F., Bleeker, P. M., Schuurink, R. C., Pallini, A., Oliveira, M. G. A., Lima, E. R., Kant, M., Sabelis, M. W., & Janssen, A. (2011). A herbivore that manipulates plant defence. Ecol. Lett., 14(3), 229–236. https://doi.org/10.1111/j.1461-0248.2010.01575.x
Sarmento, R. A., Lemos, F., Dias, C. R., Kikuchi, W. T., Rodrigues, J. C. P., Pallini, A., Sabelis, M. W., & Janssen, A. (2011). A herbivorous mite down-regulates plant defence and produces web to exclude competitors. PLoS One, 6(8), e23757. https://doi.org/10.1371/journal.pone.0023757
Schimmel, B. C. J., Ataide, L. M. S., & Kant, M. R. (2017). Spatiotemporal heterogeneity of tomato induced defense responses affects spider mite performance and behavior. Plant Signaling & Behavior, 12(10), e1370526. https://doi.org/10.1080/15592324.2017.1370526
Selman, I. W., Brierley, M. R., Pegg, G. F., & Hill, T. A. (1961). Changes in the free amino acids and amides in tomato plants inoculated with tomato spotted wilt virus. Annals of Applied Biology, 49(4), 601–615. https://doi.org/10.1111/j.1744-7348.1961.tb03657.x
Sherwood, J. L., German, T. L., Moyer, J. W., & Ullman, D. E. (2003). Tomato spotted wilt. Plant Health Instr. https://doi.org/10.1094/phi-i-2003-0613-02
Shi, X., Pan, H., Zhang, H., Jiao, X., Xie, W., Wu, Q., Wang, S., Fang, Y., Chen, G., Zhou, X., & Zhang, Y. (2014). Bemisia tabaci Q carrying tomato yellow leaf curl virus strongly suppresses host plant defenses. Sci. Rep., 4, 5230. https://doi.org/10.1038/srep05230
Shih, C.-I. T., Poe, S. L., & Cromroy, H. L. (1976). Biology, life table, and intrinsic rate of increase of Tetranychus urticae1, 3. Ann. Entomol. Soc. Am., 69(2), 362–364. https://doi.org/10.1093/aesa/69.2.362
Shrestha, A., Srinivasan, R., Riley, D. G., & Culbreath, A. K. (2012). Direct and indirect effects of a thrips-transmitted Tospovirus on the preference and fitness of its vector, Frankliniella fusca. Entomologia Experimentalis et Applicata, 145(3), 260–271. https://doi.org/10.1111/eea.12011
Singh, D. P., Moore, C. A., Gilliland, A., & Carr, J. P. (2004). Activation of multiple antiviral defence mechanisms by salicylic acid. Mol. Plant Pathol., 5(1), 57–63. https://doi.org/10.1111/j.1364-3703.2004.00203.x
Su, Q., Mescher, M., Wang, S., Chen, G., Xie, W., Wu, Q., Wang, W., & Zhang, Y.-J. (2016). Tomato yellow leaf curl virus differentially influences plant defense responses to a vector and a non-vector herbivore [Dataset].
Su, Q., Yang, F., Yao, Q., Peng, Z., Tong, H., Wang, S., Wu, Q., & Zhang, Y.-J. (2020). A non-vector herbivore indirectly increases the transmission of a vector-borne virus by reducing plant chemical defenses. Functional Ecology, 34. https://doi.org/10.1111/1365-2435.13535
Susi, H., Barrès, B., Vale, P. F., & Laine, A.-L. (2015). Co-infection alters population dynamics of infectious disease. Nature Communications, 6(1), 5975. https://doi.org/10.1038/ncomms6975
Tas, P. W. L., Boerjan, M. L., & Peters, D. (1977). The structural proteins of tomato spotted wilt virus. Journal of General Virology, 36(2), 267–279. https://doi.org/10.1099/0022-1317-36-2-267
Telfer, S., Lambin, X., Birtles, R., Beldomenico, P., Burthe, S., Paterson, S., & Begon, M. (2010). Species interactions in a parasite community drive infection risk in a wildlife population. Science, 330(6001), 243–246. https://doi.org/10.1126/science.1190333
Teodoro-Paulo, J., Alba, J. M., Charlesworth, S., Kant, M. R., Magalhães, S., & Duncan, A. B. (2023). Intraspecific variation for host immune activation by the spider mite. R Soc Open Sci, 10(6), 230525. https://doi.org/10.1098/rsos.230525
Teodoro-Paulo, J., Fernandes, C., Dong, L., Magalhães, S., Duncan, A. B., Alba, J. M., & Kant, M. R. (2023). Tomato defence suppression is a common trait in Tetranychus urticae collected from field-grown tomatoes (2023020505). Preprints. https://doi.org/10.20944/preprints202302.0505.v1
Thaler, J. S., Agrawal, A. A., & Halitschke, R. (2010). Salicylate-mediated interactions between pathogens and herbivores. Ecology, 91(4), 1075–1082. https://doi.org/10.1890/08-2347.1
Thaler, J. S., Fidantsef, A. L., & Bostock, R. M. (2002). Antagonism between jasmonate- and salicylate-mediated induced plant resistance: effects of concentration and timing of elicitation on defense-related proteins, herbivore, and pathogen performance in tomato. Journal of Chemical Ecology, 28(6), 1131–1159. https://doi.org/10.1023/A:1016225515936
Ullman, D. E., Meideros, R., Campbell, L. R., Whitfield, A. E., Sherwood, J. L., & German, T. L. (2002). Thrips as vectors of tospoviruses. In Advances in botanical research (Vol. 36, pp. 113–140). Academic Press. https://doi.org/10.1016/S0065-2296(02)36061-0
Van den Hurk, J., Tas, P. W. L., & Peters, D. (1977). The ribonucleic acid of tomato spotted wilt virus. J. Gen. Virol., 36(1), 81–91. https://doi.org/10.1099/0022-1317-36-1-81
Vijayan, P., Shockey, J., Lévesque, C. A., Cook, R. J., & Browse, J. (1998). A role for jasmonate in pathogen defense of Arabidopsis. Proceedings of the National Academy of Sciences, 95(12), 7209–7214. https://doi.org/10.1073/pnas.95.12.7209
Wang, L., & Wu, J. (2013). The essential role of jasmonic acid in plant–herbivore interactions – using the wild tobacco Nicotiana attenuata as a model. Journal of Genetics and Genomics, 40(12), 597–606. https://doi.org/10.1016/j.jgg.2013.10.001
Whitfield, A. E., Falk, B. W., & Rotenberg, D. (2015). Insect vector-mediated transmission of plant viruses. Virology, 479–480, 278–289. https://doi.org/10.1016/j.virol.2015.03.026
Young, S. S. Y., Wrensch, D. L., & Kongchuensin, M. (1986). Control of sex ratio by female spider mites. Entomologia Experimentalis et Applicata, 40(1), 53–60. https://doi.org/10.1111/j.1570-7458.1986.tb02155.x
Zarate, S. I., Kempema, L. A., & Walling, L. L. (2007). Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiology, 143(2), 866–875. https://doi.org/10.1104/pp.106.090035
Zélé, F., Magalhães, S., Kéfi, S., & Duncan, A. B. (2018). Ecology and evolution of facilitation among symbionts. Nat. Commun., 9(1), 4869. https://doi.org/10.1038/s41467-018-06779-w
Zhang, T., Luan, J.-B., Qi, J.-F., Huang, C.-J., Li, M., Zhou, X.-P., & Liu, S.-S. (2012). Begomovirus-whitefly mutualism is achieved through repression of plant defences by a virus pathogenicity factor. Mol. Ecol., 21(5), 1294–1304. https://doi.org/10.1111/j.1365-294X.2012.05457.x
Zhi, J., Margolies, D. C., Nechols, J. R., & Boyer Jr, J. E. (2006). Host-plant-mediated interaction between populations of a true omnivore and its herbivorous prey. Entomologia Experimentalis et Applicata, 121(1), 59–66. https://doi.org/10.1111/j.1570-8703.2006.00456.x
Zilio, G., & Koella, J. C. (2020). Sequential co-infections drive parasite competition and the outcome of infection. Journal of Animal Ecology, 89(10), 2367–2377. https://doi.org/10.1111/1365-2656.13302



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}