Abstract
Foliar insects inevitably ingest microorganisms residing both on the surface and within the internal tissues of the leaves they consume. Among these, epiphytes â microbes inhabiting the leaf surface â are the first to interact with the insects. While some studies have examined the impact of specific epiphytic bacterial strains on insect performance, the role of the entire epiphytic community remains largely unexplored. In this study, we manipulated the presence/absence of the epiphytic community on Datura inoxia leaves and evaluated its effect on the performance of its specialist herbivorous insect Lema daturaphila. Removing the entire epiphytic community affected larval survival, but this effect varied over time. A significant difference was observed only in December, when larvae feeding on leaves without epiphytes experienced a reduced risk of mortality. We also found that larvae feeding on leaves without epiphytes consumed more foliar tissue but were less efficient compared to those eating control leaves. In addition, larvae growing in December were lighter, consumed less, developed more slowly, but showed higher feeding efficiency and adult mass compared to February. Overall, our findings suggest that the epiphytic community associated with Datura inoxia increases the feeding efficiency of its specialist insect, Lema daturaphila.
1 Introduction
Plants and their folivorous insects are constantly, and inevitably, interchanging microorganisms (Felton and Tumlison, 2008; Mason, 2020; Mayoral-Peña et al., 2020). While consuming the foliar tissue, larvae are exposed to a plethora of microorganisms associated with both the surface (epiphytes) and the internal tissues of the leaves (endophytes) (Carvalho and Castillo, 2018; Lindow and Brandl, 2003). While the microorganisms of the epiphytic and endophytic communities belong to different taxonomic groups, bacteria is the most abundant one (Dastogeer et al., 2020). Recent studies have shown that epiphytes are involved not only in the growth and development of their host plants (Francis et al., 2020; Goswami et al., 2021), but can also act as a protective barrier by negatively affecting pathogens and herbivorous insects (Goswami et al., 2021; Mason, 2020; Peters et al., 2017; Poveda, 2021). In particular, epiphytes can influence insect feeding preferences (Peters et al., 2017) by contributing to the production of secondary metabolites that enhance plant defences (Poveda, 2021). Despite their ecological significance and potential influence on plantâinsect interactions, the possible effects of the entire epiphytic community on insect performance are still poorly understood.
Changes in environmental conditions play a critical role in shaping the diversity, composition, and ecological interactions of microorganisms in the phyllosphere â the above ground portions of plants â affecting both bacterial and fungal microbiota (Cordier et al., 2012; Huang et al., 2023; Peñuelas et al., 2012). Indeed, the phyllosphere represents a harsh and unpredictable environment, with fluctuations in moisture, UV and temperature levels along with limited resource availability that hinder the growth of most microbial organisms (Koskella, 2020). In particular, studies show that rising temperatures reduce the abundance of beneficial foliar bacteria while promoting the proliferation of pathogenic ones (Huang et al., 2023). Thermal fluctuations also influence metabolic compound production and stomatal activity, which, in turn, impact microbial colonization (Aydogan et al., 2020; Huang et al., 2023) and alter the phylogenetic structure of microbial communities (Aydogan et al., 2020). Further research is needed to determine the extent to which temperature-driven changes influence phyllosphere dynamics and, ultimately, plantâinsect interactions (Aydogan et al., 2020; Huang et al., 2023).
The composition and structure of an insectâs gut microbiome are strongly influenced by the phyllosphere (Guidolin and Consoli, 2017; Mogouong et al., 2021). As microorganisms pass through the food bolus, they modify the resident intestinal microbiota (Douglas, 2015; Itoh et al., 2020; Mason, 2020). In particular, it has been suggested that epiphytes may directly impact leaf tissue consumption by playing a functional role in digestion or detoxification (Mogouong et al., 2021). Several herbivorous insects, including Aphis fabae (Pons et al., 2019), Sitobion miscanthi (Li et al., 2018), Macrosteles quadripunctulatus (Gonella et al., 2015), Empoasca vitis (Gonella et al., 2015), Trichoplusia ni (Leite-Mondin et al., 2021), and Lymantria dispar (Broderick et al., 2004), are known to acquire microorganisms during phytophagy. This microbial exchange fosters interactions between the gut microbiota and the phyllosphere, ultimately influencing insect feeding capacity. Notably, variations in the gut microbiota composition can persist across generations (Freitak et al., 2009a; Leite-Mondin et al., 2021). Whether epiphytes directly or indirectly affect insect performance remains an open question requiring further study.
Different studies report that bacteria present in the diet of phytophagous insects can significantly affect their development, physiology, and pathogen susceptibility, with effects varying by bacterial species. Some studies report a negative impact on larval survival and developmental time, though no significant effect on pupal mass in the cabbage looper T. ni (Freitak et al., 2009b). Similarly, other research found negative effects on larval mass and growth (Shikano et al., 2015) in T. ni and reduced offspring viability in Malacosoma californicum (Olson et al., 2017). In contrast, in Sitobion miscanthi, bacterial presence positively influenced offspring production and increased adult mass (Qian et al., 2018). In a similar way, Acinetobacter appears to play a beneficial role by facilitating the detoxification of toxic compounds in Lymantria dispar (Mason et al., 2014; Shikano et al., 2015; Olson et al., 2017). Nevertheless, all these effects were found when using individual bacterial strains, and the combined effect of the different epiphytic microbes might be very different from those found for each particular strain (Peters et al., 2017). Because epiphytes belonging to different taxonomic groups (bacteria, algae, viruses, fungi) interact among themselves before and after consumption, evaluating the effect of the entire community provides complementary but essential information about how microorganisms inhabiting the phyllosphere might shape plantâinsect interactions (Agler et al., 2016). To test whether the entire epiphytic community affects insect performance, we manipulated its presence-absence on the leaves of Datura inoxia and used these leaves to feed larvae of the specialist insect Lema daturaphila.
2 Material and methods
2.1 Study system
Datura inoxia Mill (Solanaceae) is an herbaceous ruderal biennial plant that grows on the outskirts of Queretaro city in Central Mexico, where it is often eaten by its specialist insect Lema daturaphila Kogan and Goeden (Coleoptera: Chrysomelidae) (Kariñho-Betancourt, 2019; MartÃnez y DÃaz de Salas et al., 2017). In a previous study, we showed that the epiphytic bacterial community associated with the leaves of D. inoxia is more diverse compared to those bacterial communities associated with other tissues within the plant (Mayoral-Peña et al., 2022). Thus, the larvae of L. daturaphila are constantly exposed and eating a myriad of microorganisms. In addition, functional inference analyses suggest that some epiphytic bacteria might be involved in the biosynthesis and degradation of secondary metabolites (Mayoral-Peña et al., 2022) with possible consequences upon insect consumption.
2.2 Experimental design
To test the possible effects of the whole community of microorganisms inhabiting the surface of the leaves (known as the phylloplane) on insect performance, we manipulated the presence/absence of this microbial community following a complete factorial design. Starting in late August of 2021, sets of around 100 seeds were germinated in the greenhouse every month until December. When the seedlings from each set fully expanded their first leaf, they were transplanted individually in 4L pots filled with commercial potting soil. All plants were fertilised at 30 and 60 days after transplant with Peters (1g/L containing a balanced combination of nitrogen, phosphorus, potassium, magnesium, and essential micronutrients), and were kept inside the greenhouse until their fourth leaf was fully expanded (approximately two months after transplant, right after being fertilised for a second time) to prevent any kind of damage or consumption. After this period, the plants were moved outside the greenhouse and placed near naturally growing D. inoxia on the university campus to facilitate the natural acquisition of epiphytes from the environment. The plants stayed outside for approximately four weeks until their eighth leaf was fully expanded. At the same time when the first set of seedlings were growing in the greenhouse, around 150 eggs, larvae and adults of L. daturaphila were collected from the locality of El Marqués (20â °37â²28.1â³N, 100â °22â²07.2â³W) to start a colony in the laboratory under controlled conditions (25â °C and a 12:12 photoperiod) (Espinosa and Fornoni, 2006).
Given that D. inoxia is a biennial plant and that some larvae and adults can be found in the field late into the winter, we were curious to evaluate how the effects of our treatment might vary over time due to changes in environmental conditions. Thus, the experiment began in November 2021 and lasted for four months, using a total of 255 plants. From each plant, two consecutive leaves were selected, cut using a sterile blade, and placed on ice until processed in the laboratory. In most cases, the seventh and eighth leaves (counted from the bottom up) were chosen. However, in ten plants, the ninth and tenth leaves were selected instead, as the previous leaves showed signs of damage. Each plant was sampled only once. Once in the laboratory, under a laminar flux cabinet, one of the leaves was placed in a sterile Petri dish and left untreated (control treatment) while the other one was treated with a series of washes to surface-sterile it (no epiphytes treatment). Overall, the serial washes were as follows: one wash using an epiphyte buffer (KHâPOâ, KâHPOâ, triton) (1 min), four washes with sterile water (2 min each), and a final mechanical cleaning using a sterile gauze (modified from Desgarennes et al., 2014). From the last wash with sterile water, 100
During the experiment the following variables were measured: survival (days until death), larval mass (g), foliar consumption (cm²), efficiency of foliar consumption (g/cm²), developmental time (days from hatching to emergence) and adult mass (Garrido et al., 2012). Because previous studies performed with the same beetle show that larval mass at eclosion is the same among larvae (Espinosa and Fornoni, 2006; Garrido et al., 2012; Garrido et al., 2022), we measured larval mass only at day seven using a digital scale (Mettler Toledo, PL303). To estimate foliar consumption, photographs of the leaves were taken after they were removed from the Petri dish or when the larvae pupated and were analysed using the Fiji distribution of ImageJ (Rueden et al., 2017) and total area consumed per larva was then calculated. Efficiency of foliar consumption was estimated as the ratio between larva mass and foliar consumption. Adult mass was measured using the digital scale.
2.3 Statistical analysis
The statistical model for all the variables included treatment (with or without epiphytes), block (the months when the experiment was performed) and their interaction as sources of variation. Larval survival was analysed with a Proportional Hazards mixed model using the coxph function in the survival package (Lin and Zelterman, 2002). Due to variation in mating and oviposition patterns within the insect colony over the course of the experiment, which affected the total number of replicates per month, only samples from December and February were included in the analyses. The analysis for larval survival was done with 248 replicates: 130 for December (65 per treatment: control and without epiphytes) and 118 for February (59 per treatment). For subsequent analyses, replicates with missing values were excluded â for example, in some cases, larval consumption could not be accurately measured because larvae consumed entire leaves before photographs were taken, preventing measurement. All the other variables were analysed with two-way ANOVAs with type III sums of squares to accommodate the unbalanced design, using the Anova function in the car package (Fox and Weisberg, 2019). Sample sizes for the ANOVAs were 58 in December (30 control and 28 without epiphytes) and 46 in February (26 control and 20 without epiphytes). To investigate potential trade-offs among the measured life-history traits (excluding survival), correlation coefficients were calculated separately for each block (December and February). Statistical significance was assessed using a Bonferroni correction with the psych package (Revelle, 2017). All the analyses and plots were done using R (R Core Team, 2024). The data is available at the following link: https://doi.org/10.5281/zenodo.15485949 and tab Supplementary Materials on the landing page of this article at brill.com/pami.
3 Results
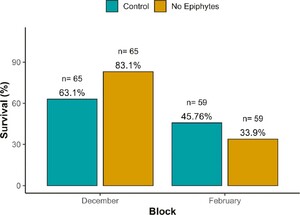
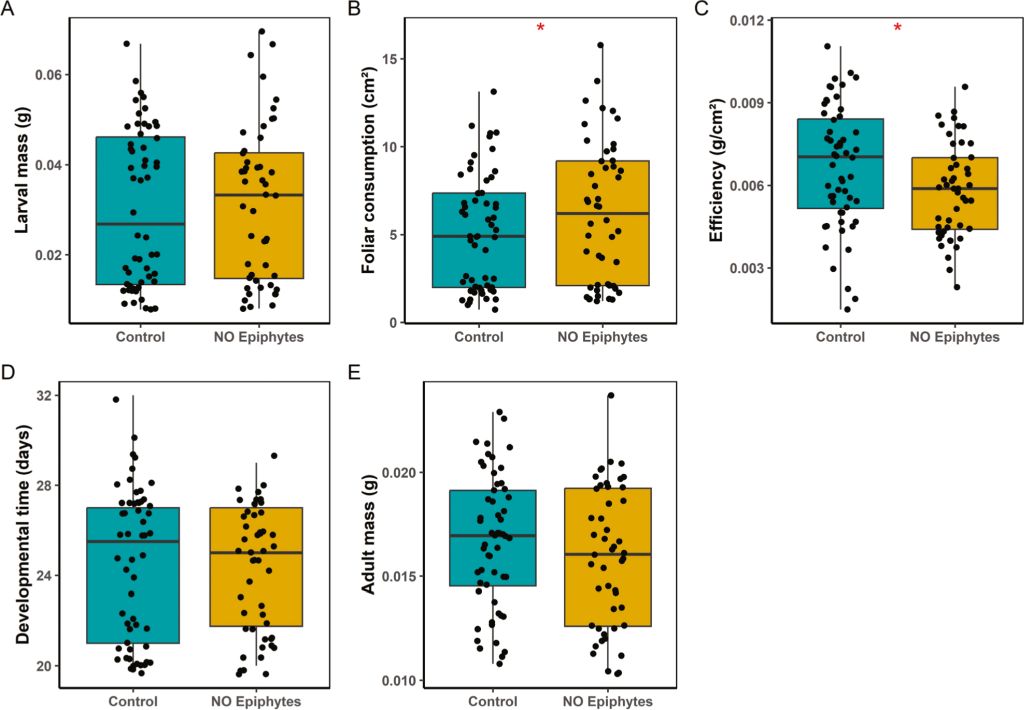
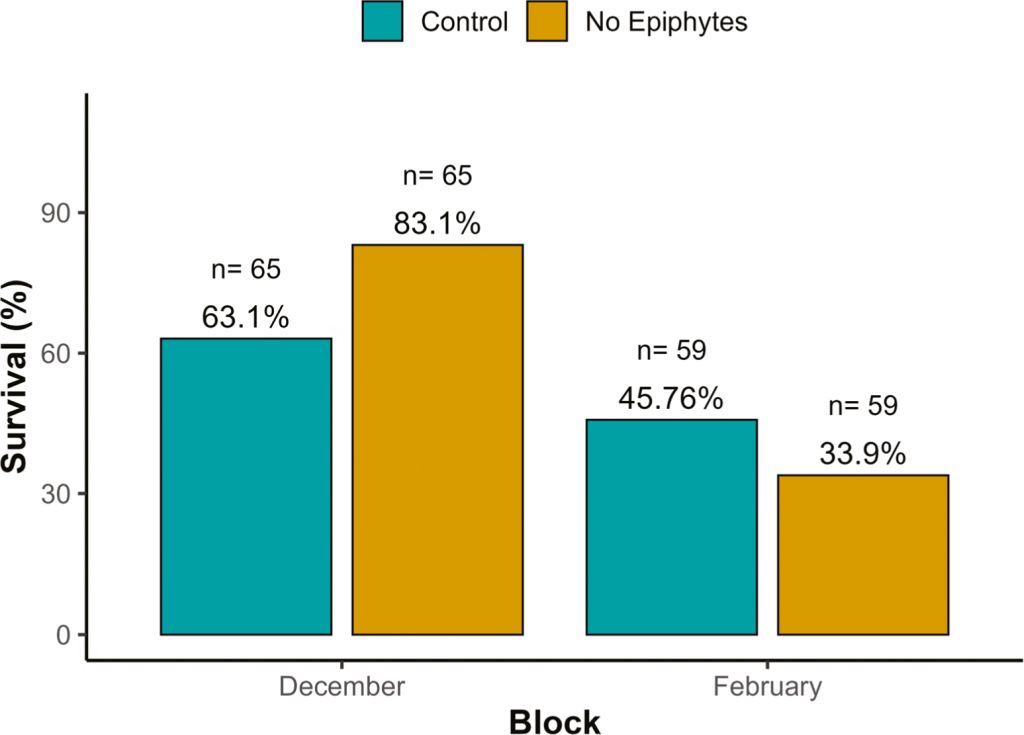
For larval survival, we detected a significant effect of the treatment (P = 0.0088; Table S1) and of the interaction between treatment and block (P = 0.0032; Table S1), indicating that the effect of our treatment varied over time. In December, larvae fed with leaves without epiphytes experienced a lower risk of mortality compared to the control group (Hazard Ratio (HR) = 0.39; 95% CI: 0.19â0.79, P = 0.0088). Correspondingly, survival was higher in the no-epiphyte treatment (83.1%) than in the control (63.0%; Figure 1), representing a 1.32-fold increase. In February, this trend reversed slightly: survival was lower for larvae fed with leaves without epiphytes (33.9%) compared to the control group (45.8%; Figure 1). Treatment also affected foliar consumption (P = 0.028; Table S1) and efficiency (P = 0.023; Table S1). When eating leaves without epiphytes, larvae increased their foliar consumption by 21% (Figure 2B) but their efficiency of foliar consumption was 13% less (Figure 2C) compared to larvae eating control leaves. All the other variables were not affected by the treatment (Table S1).

Survival percentages by treatment across blocks (December and February)
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10002
Effect of consuming leaves with the full epiphytic community (control) versus leaves with no epiphytes on: (A) Larval mass, (B) Foliar consumption, (C) Efficiency of foliar consumption, (D) Developmental time, and (E) Adult mass. * indicates P < 0.01
Citation: Journal of Plant-Arthropod-Microbe Interactions 2, 1-2 (2025) ; 10.3920/29501679-bja10002
We also found a significant effect of block (i.e. month) on larval mass (P < 0.0001; Table S1), consumption (P < 0.0001; Table S1), developmental time (P < 0.0001; Table S1) and adult mass (P < 0.0001; Table S1). The monthly mean temperatures exhibited variability, with recorded values of 14.5â °C in December, and 17â °C in February. Larval mass showed a tendency to increase as the months passed, from 0.019 ± 0.0014 g (mean ± SE) in December to more than double its value in February (Table S2). Foliar consumption also increased, reaching its highest value in February (8.75 ± 0.37 cm²) compared to December (3.34 ± 0.32 cm²). In contrast, feeding efficiency was 1.17 times higher in December than in February. Larvae completed their development faster with the passing months, with the shortest developmental time recorded in February (21.3 ± 0.2 days) compared to the one in December (26.8 ± 0.18 days). Adult mass was 1.29 times greater in December compared to the mass reached by adults in February. No significant interactions between treatment and block were detected for growth- or consumption-related parameters.
Finally, after Bonferroni correction, we detected significant correlations among traits. In December, larval mass was positively correlated with foliar consumption (r = 0.75; P < 0.0001), while efficiency was negatively correlated with consumption (r = â0.62; P < 0.0001). In February, larval and adult mass were positively correlated (r = 0.58; P < 0.0001), and the negative relationship between consumption and efficiency persisted (r = â0.76; P < 0.0001; Table S3).
4 Discussion and conclusion
We found that the presence of the epiphytic community impacted larval performance. Epiphytes enhanced the efficiency of foliar consumption and adult mass, while the absence of the whole epiphytic community resulted in an increase in foliar consumption. On the other hand, larvae feeding on leaves with the complete endophyte community had longer development times and lower survival than larvae that consumed leaves with no epiphytes during one of the colder months (December). These results highlight the potential role of epiphytes not only on insect performance, but in shaping the ecology and evolution of plantâinsect interactions.
Our findings show that L. daturaphila survival is not determined solely by the presence or absence of epiphytes, but by the interaction between our treatment and environmental changes. For example, in December, when mean temperature was 14.5â °C, larvae feeding on epiphyte-bearing leaves had lower survival, but in February, when temperature was higher, survival declined across both treatments, and a reverse trend was observed. However, this trend was not significant, suggesting that warmer conditions may buffer or alter the effect of our treatments. Previous studies have shown that in T. ni and M. californicum, the presence of epiphytes had minimal impact under favorable conditions but became detrimental under stress, such as food scarcity (Olson et al., 2017; Shikano et al., 2015). In our study, several abiotic factors associated with seasonal variation â primarily temperature, UV radiation, and light availability â may have influenced this interaction. These results suggest that the role of epiphytes is more complex than previously assumed and highlight the need for further research under varying environmental conditions.
Interestingly, we found no significant difference in larval mass (measured at day seven) between treatments. One possible explanation is that larvae may compensate for the absence of epiphytes by increasing foliar consumption, a common strategy among herbivores facing suboptimal nutritional conditions (Clancy et al., 2014; Yang and Joern, 1994). Although the underlying mechanisms remain unclear, previous research suggests this compensation may stem from reduced efficiency in polysaccharide degradation, a process often mediated by gut microbiota (Schmidt and Engel, 2021). Since carbohydrate metabolism plays a key role in weight gain â particularly important for pupal development in holometabolous insects (Le Gall and Behmer, 2014) â inefficient carbohydrate processing may drive increased food intake. Additionally, epiphytic microbes may aid in the breakdown of plant secondary metabolites (Mason et al., 2014), a function commonly attributed to the gut microbiota (Douglas, 2015; Jing et al., 2020). Some studies even suggest that epiphytes contribute to glycoside degradation, potentially enhancing larval growth (Mason et al., 2014). However, these hypotheses remain speculative in our system, and further research is needed to clarify whether microbial interactions primarily influence nutrient acquisition, carbohydrate metabolism, detoxification of plant compounds, or other physiological processes.
In addition, we found that when eating leaves without epiphytes larvae were less efficient in converting leaf biomass into caterpillar biomass. Although we do not have data to shed light on a possible mechanism, previous studies suggest that phyllosphere microbes can influence insect gut microbiota (Aluja et al., 2021; Leite-Mondin et al., 2021; Mogouong et al., 2021), potentially enhancing digestion and leaf tissue breakdown (Guidolin and Cônsoli, 2017; Qian et al., 2018). One possible mechanism involves the production of antimicrobial compounds by epiphytes, which may shape gut microbial communities by promoting beneficial strains (Coyte et al., 2015; Schmidt and Engel, 2021). Functional analyses of D. inoxia epiphytes (unpublished data) suggest they produce antibiotics such as ansamycins and tetracyclines, supporting this idea. Alternatively, epiphytes may directly aid in foliar digestion through enzymes involved in nutrient breakdown and detoxification (Gupta and Nair, 2020; Engel and Moran, 2013). In their absence, these functions may be less efficiently carried out by the insect alone. Overall, the improved feeding efficiency likely results from interactions between epiphytes, gut microbes, and host enzymes (Newell and Douglas, 2014; Jing et al., 2020). Further research is needed to clarify the specific microbial and physiological mechanisms involved.
All variables measured, except survival and efficiency, showed differences across months (block factor in the analyses; Table S1). From December through February mean temperature rose from 14.5â °C to 17â °C and larval mass, consumption and adult mass increased accordingly. These patterns align with evidence that temperature shapes insect performance, potentially by altering microbiota composition and interactions (Liu et al., 2021; Sun et al., 2022). Notably, when growing in December at 14.5â °C, epiphytes appeared to prolong insect development. Microbial symbionts are known to enhance thermal tolerance in insects (Fahim et al., 2020; Jaramillo and Castañeda, 2021), and since cold slows development (Cui et al., 2018; Huang et al., 2018), epiphytes might help mitigate this delay. However, when growing in February at 17â °C, these differences disappeared. These results suggest temperature or associated factors such as daylength may reshape hostâmicrobiota interactions, thereby influencing insect development. It has been suggested that thermal variation affects microbial diversity, structure, and interactions within the phyllosphere (Cordier et al., 2012; Huang et al., 2023; Peñuelas et al., 2012), potentially reducing beneficial taxa and enhancing pathogens (Aydogan et al., 2020; Huang et al., 2023). Such shifts could explain the observed developmental patterns. Nevertheless, further research is needed to clarify how temperature drives changes in microbial communities and its posterior effect on plantâinsect interactions.
In December, foliar consumption and larval mass were positively correlated, but this relationship was not observed in February. Conversely, larval and adult mass were positively correlated in February, whereas no significant association was found in December. These results suggest that environmental variation, particularly temperature, plays a key role in shaping life-history traits. The absence of a correlation between larval and adult mass in December may be explained by the fact that, under cooler conditions, larvae weighed at day seven were likely still far from pupation and continued growing for more days thereafter. To gain a more comprehensive understanding of trait associations, future studies should consider separating total developmental time into larval and pupal stages to better understand these relationships. Surprisingly, we did not detect any correlation between developmental time and any other variable. For example, a well-known trade-off between adult mass and development (Stearns, 1989; Roff, 2000; Zera and Harshman, 2001) was not detected in either month. Although no significant differences were found in larval or adult mass between treatments, future research should explore whether the presence or absence of epiphytes in the diet influences female sex ratio or fecundity. Adult mass is a key fitness trait, as larger females tend to produce more and larger eggs (Berger et al., 2008; Calvo and Molina, 2005; Sturm, 2016), and larger males often have a mating advantage (Hanks, 1996; Agosta, 2010).
Overall, our study offers a first step toward understanding how the whole epiphytic community affects the performance of a specialist insect. Several questions remain open for future research. For example, characterizing the functional profiles of bacterial and fungal communities on Datura inoxia leaves, followed by isolating and manipulating specific strains, could help clarify their roles. Also, investigating how these microbes influence insect performance, through nutrient assimilation, immune modulation, or detoxification, would shed light on underlying mechanisms. Finally, tracking adult insects to assess potential effects on reproduction could reveal longer-term fitness impacts.
Acknowledgements
The authors declare no conflict of interest. We thank the National Council for Humanities, Science, and Technology (CONAHCYT) for the support provided through the doctoral scholarship to ZM-P, as well as the funding granted by the FONFIVE-UAQ-2024 project (FNB-2024-04) to EG. We also express our gratitude to Dr. Juan Fornoni and three anonymous reviewers for their comments and feedback, which enriched this work.
References
Agler, M. T., Ruhe, J., Kroll, S., Morhenn, C., Kim, S. T., Weigel, D., Kemen, E. M., 2016. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biology 14(1): 1â31, https://doi.org/10.1371/journal.pbio.1002352
Agosta, S. J., 2010. Male body size and mating success and their relation to larval host plant history in the moth Rothschildia lebeau in Costa Rican dry forest. Biotropica 42(2): 201â207, https://doi.org/10.1111/j.1744-7429.2009.00556.x
Aluja, M., Zamora-Briseño, J. A., Pérez-Brocal, V., Altúzar- Molina, A., Guillén, L. ⦠Alonso-Sánchez, A. G., Moya, A., (2021). Metagenomic survey of the highly polyphagous Anastrepha ludens developing in ancestral and exotic hosts reveals the lack of a stable microbiota in larvae and the strong Influence of metamorphosis on adult gut microbiota. Frontiers in Microbiology, 12: 685937, https://doi.org/10.3389/fmicb.2021.685937
Beran, F., Köllner, T. G., Gershenzon, J., Tholl, D., 2019. Chemical convergence between plants and insects: biosynthetic origins and functions of common secondary metabolites. New Phytologist 223: 52â67, https://doi.org/10.1111/nph.15718
Berger, D., Walters, R., Gotthard, K., 2008. What limits insect fecundity? Body size- and temperature-dependent egg maturation and oviposition in a butterfly. Functional Ecology 22: 523â529, https://doi.org/10.1111/j.1365-2435.2008.01392.x
Broderick, N. A., Raffa, K. F., Goodman, R. M., Handelsman, J., 2004. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Applied and Environmental Microbiology 70: 293â300, https://doi.org/10.1128/AEM.70.1.293
Calvo, D., Molina, J. M., (2005). Fecundityâbody size relationship and other reproductive aspects of Streblote panda (Lepidoptera: Lasiocampidae). Annals of the Entomological Society of America 98: 191â196, https://doi.org/10.1603/0013-8746(2005)098[0191:FSRAOR]2.0.CO;2
Carvalho, S. D., Castillo, J. A., 2018. Influence of light on plantâ phyllosphere interaction. Frontiers in Plant Science 9: 1â16, https://doi.org/10.3389/fpls.2018.01482
Clancy, K. M. and Price, P. W., 1987. Rapid herbivore growth enhances enemy attack: sublethal plant defenses remain a paradox. Ecology 68: 733â737, http://www.jstor.org/stable/1938479
Cordier, T., Robin, C., Capdevielle, X., Fabreguettes, O., Desprez-Loustau, M.-L., Vacher, C., 2012. The composition of phyllosphere fungal assemblages of European beech (Fagus sylvatica) varies significantly along an elevation gradient. New Phytologist 196: 510â519, DOI:10.1111/j.1469-8137.2012.04284.x
Coyte, K. Z., Schluter, J., Foster, K. R., 2015. The ecology of the microbiome: Networks, competition, and stability. Science 350: 663â666, https://doi.org/10.1126/science.aad2602
Cui, J., Zhu, S., Bi, R., Xu, W., Gao, Y., Shi, S., 2018. Sampling and biostatistics effect of temperature on the development, survival, and fecundity of Heliothis viriplaca (Lepidoptera: Noctuidae). Journal of Economic Entomology 111: 1940â1946. https://doi.org/10.1093/jee/toy151
Dastogeer, K. M. G., Tumpa, F. H., Sultana, A., Akter, M. A., Chakraborty, A., 2020. Plant microbiome â an account of the factors that shape community composition and diversity. Current Plant Biology 23: 100161, https://doi.org/10.1016/j.cpb.2020.100161
Desgarennes, D., Garrido, E., Torres-Gomez, M. J., Peña-Cabriales, J. J., Partida-Martinez, L. P., 2014. Diazotrophic potential among bacterial communities associated with wild and cultivated Agave species. FEMS Microbiology Ecology 90: 844â857, https://doi.org/10.1111/1574-6941.12438
Douglas, A. E., 2015. Multiorganismal insects: Diversity and function of resident microorganisms. Annual Review of Entomology 60: 17â34, https://doi.org/10.1146/annurev-ento-010814-020822
Engel, P., Moran, N. A., 2013. The gut microbiota of insects â diversity in structure and function. FEMS Microbiology Reviews 37: 699â735, https://doi.org/10.1111/1574-6976.12025
Espinosa, E. G., Fornoni, J., 2006. Host tolerance does not impose selection on natural enemies. New Phytologist 170: 609â614, https://doi.org/10.1111/j.1469-8137.2006.01681.x
Fahim, M. R., Wang, Y., Cai, Z., Bai, S., Yao, Z., Awan, A., Zhang, Z., Zheng, W., Zhuang, H., 2020. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog 16: e1008441, https://doi.org/10.1371/journal.ppat.1008441
Felton, G. W., Tumlinson, J. H., 2008. Plantâinsect dialogs: complex interactions at the plantâinsect interface. Current Opinion in Plant Biology 11: 457â463, https://doi.org/10.1016/j.pbi.2008.07.001
Fox, J., Weisberg, S. 2019. An R Companion to Applied Regression, Third edition. Sage, Thousand Oaks CA. <https://www.john-fox.ca/Companion/>
Francis, F., Jacquemyn, H., Delvigne, F., Lievens, B., 2020. From diverse origins to specific targets: Role of microorganisms in indirect pest biological control. Insects 11: 1â14, https://doi.org/10.3390/insects11080533
Freitak, D., Heckel, D. G., Vogel, H., 2009a. Dietary-dependent trans-generational immune priming in an insect herbivore. Proceedings of the Royal Society B: Biological Sciences 276: 2617â2624, https://doi.org/10.1098/rspb.2009.0323
Freitak, D., Wheat, C. W., Heckel, D. G., Vogel, H., 2007b. Immune system responses and fitness costs associated with consumption of bacteria in larvae of Trichoplusia ni. BMC Biology 5: 56, https://doi.org/10.1186/1741-7007-5-56
Garrido, E., Andraca-go, G., Fornoni, J., 2012. Local adaptation: simultaneously considering herbivores and their host plants. New Phytologist, 445â453. https://doi.org/10.1111/j.1469-8137.2011.03923.x
Garrido, E., Boege, K., DomÃnguez, C. A., Fornoni, J., 2022. Priming by insects: differential effects of sympatric and allopatric priming upon plant performance and tolerance to herbivory. Plants, 11(24): 3567. https://doi.org/10.3390/plants11243567
Gonella, E., Pajoro, M., Marzorati, M., Crotti, E., Mandrioli, M., Pontini, M., Bulgari, D., Negri, I., Sacchi, L., Chouaia, B., Daffonchio, D., Alma, A., 2015. Plant-mediated interspecific horizontal transmission of an intracellular symbiont in insects. Scientific Reports 5: 15811, https://doi.org/10.1038/srep15811
Goswami, S., Goel, N., Majumdar, R. S., 2021. Phylloplane microbes impact host physiology: a review. Journal of Plant Protection Research 61: 213â221, https://doi.org/10.24425/jppr.2021.137949
Guidolin, A. S., Cônsoli, F. L., 2017. Symbiont diversity of Aphis (Toxoptera) citricidus (Hemiptera: Aphididae) as Influenced by host plants. Microbial Ecology 73: 201â210, https://doi.org/10.1007/s00248-016-0892-8
Gupta, A., Nair, S., 2020. Dynamics of insectâmicrobiome interaction influence host and microbial symbiont. Frontiers in Microbiology 11: 1357, https://doi.org/10.3389/fmicb.2020.01357
Hanks, L. M., 1996. Body size influences mating success of the eucalyptus longhorned borer (coleoptera: cerambycidae). Journal of Insect Behavior 9: 369â382, https://doi.org/10.1007/BF02214016
Huang, W., Long, C., Lam, E., 2018. Roles of plant-associated microbiota in traditional herbal medicine. Trends in Plant Science 23: 559â562, https://doi.org/10.1016/j.tplants.2018.05.003
Itoh, H., Tago, K., Hayatsu, M., Kikuchi, Y., 2018. Detoxifying symbiosis: microbe-mediated detoxification of phytotoxins and pesticides in insects. Natural Product Reports 35: 434â454, https://doi.org/10.1039/C7NP00051K
Jaramillo, A., Castañeda, L. E., 2021. Gut microbiota of Drosophila subobscura contributes to its heat tolerance and is sensitive to transient thermal stress. Frontiers in Microbiology 12: 654108, https://doi.org/10.3389/fmicb.2021.654108
Jing, T. Z., Qi, F. H., Wang, Z. Y., 2020. Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome 8: 1â20, https://doi.org/10.1186/s40168-020-00823-y
Kariñho-Betancourt, E., Agrawal, A. A., Halitschke, R., Núñez- Farfán, J., 2015., Phylogenetic correlations among chemical and physical plant defenses change with ontogeny. New Phytologist 206: 796â806, https://doi.org/10.1111/nph.13300
Koskella, B., 2020. The phyllosphere. Current Biology 30(19): R1143âR1146, https://doi.org/10.1016/j.cub.2020.07.058
Leite-Mondin, M., DiLegge, M. J., Manter, D. K., Weir, T. L., Silva-Filho, M. C., Vivanco, J. M., 2021. The gut microbiota composition of Trichoplusia ni is altered by diet and may influence its polyphagous behavior. Scientific Reports 11: 1â16, https://doi.org/10.1038/s41598-021-85057-0
Li, Q., Fan, J., Sun, J., Wang, M. Q., Chen, J., 2018. Plant-mediated horizontal transmission of Hamiltonella defensa in the wheat aphid Sitobion miscanthi. Journal of Agricultural and Food Chemistry 66: 13367â13377, https://doi.org/10.1021/acs.jafc.8b04828
Lin, H., Zelterman, D., 2002. Modeling survival data: extending the cox model. Technometrics 44: 85â86, https://doi.org/10.1198/tech.2002.s656
Lindow, S. E., Brandl, M. T., 2003. Microbiology of the phyllosphere. Applied and Environmental Microbiology 69: 1875â1883, https://doi.org/10.1128/AEM.69.4.1875-1883.2003
Liu, J., Wang, C., Desneux, N., Lu, Y., 2021. Impact of temperature on survival rate, fecundity, and feeding behaviour of two aphids, Aphis gossypii and Acyrthosiphon gossypii, when reared on cotton. Insects 12: 565, https://doi.org/10.3390/insects12060565
MartÃnez y DÃaz de Salas, M., Sandoval, Luis Hernández Pantoja Hernández, Y., Gómez Sánchez, M., Bárcenas Luna, R., Cabrera Luna, A., 2017. GuÃa ilustrada de la flora del Valle de Querétaro. Universidad Autónoma de Querétaro, Editorial Universitaria, Querétaro, 250 pp. https://biblioteca.semarnat.gob.mx/janium/Documentos/Ciga/libros2018/CD003341.pdf
Mason, C. J., 2020. Complex relationships at the intersection of insect gut microbiomes and plant defenses. Journal of Chemical Ecology 46: 793â807, https://doi.org/10.1007/s10886-020-01187-1
Mason, C. J., Rubert-nason, K. F., Lindroth, R. L., Raffa, K. F., 2014. Aspen defense chemicals Influence midgut bacterial community composition of gypsy moth. Journal of chemicals influence 41: 75â84, https://doi.org/10.1007/s10886-014-0530-1
Mayoral-Peña, Z., Ãlvarez-MartÃnez, R., Fornoni, J., Garrido, E., 2020. The extended Microbiota: How microbes shape plant-insect interactions. In: Nuñez-Farfán and P. J. Valverde (eds.). Springer, pp. 123â145. https://doi.org/10.1007/978-3-030-46012-9_7
Mayoral-peña, Z., Lázaro-Vidal, V., Fornoni, J., Ãlvarez- MartÃnez, R., Garrido, E., 2022. Studying plantâinsect Interactions through the analyses of the diversity, composition, and functional inference of their bacteriomes. Microorganisms 11: 40, https://doi.org/10.3390/microorganisms11010040
Miyashita, A., Hirai, Y., Sekimizu, K., Kaito, C., 2015. Antibiotic- producing bacteria from stag beetle mycangia. Drug Discoveries and Therapeutics 9: 33â37, https://doi.org/10.5582/ddt.2015.01000
Mogouong, J., Constant, P., Legendre, P., Guertin, C., 2021. The phyllosphere microbiome of host trees contributes more than leaf phytochemicals to variation in the Agrilus planipennis gut microbiome structure. Scientific Reports 11: 15911, https://doi.org/10.1038/s41598-021-95146-9
Newell, P. D., Douglas, A. E., 2014. Interspecies interactions determine the impact of the gut microbiota on nutrient allocation in Drosophila melanogaster. Applied and Environmental Microbiology 80: 788â796, https://doi.org/10.1128/AEM.02742-13
Olson, G. L., Myers, J. H., Hemerik, L., Cory, J. S., 2017. Phylloplane bacteria increase the negative impact of food limitation on insect fitness. Ecological Entomology 42: 411â421, https://doi.org/10.1111/een.12399
Peters, B., Türke, M., Junker, R. R., 2017. Epiphytic bacteria on lettuce affect the feeding behavior of an invasive pest slug. Acta Agrobotanica 70: 1â11, https://doi.org/10.5586/aa.1708
Peñuelas, J., Rico, L., Ogaya, R., Jump, A. S., Terradas, J., 2012. Summer season and long-term drought increase the richness of bacteria and fungi in the foliar phyllosphere of Quercus ilex in a mixed Mediterranean forest. Plant Biology 14:565â575, DOI:10.1111/j.1438-8677.2011.00532.x
Pons, I., Renoz, F., Noël, C., Hance, T., 2019. Circulation of the cultivable symbiont serratia symbiotica in aphids is mediated by plants. Frontiers in Microbiology 10: 764, https://doi.org/10.3389/fmicb.2019.00764
Poveda, J., 2021., Beneficial effects of microbial volatile organic compounds (MVOCs) in plants. Applied Soil Ecology 168: 104118, https://doi.org/10.1016/j.apsoil.2021.104118
Qian, L., Jia, F., Jingxuan, S., Manqun, W., Julian, C., 2018. Effect of the Secondary symbiont Hamiltonella defensa on fitness and relative abundance of Buchnera aphidicola of wheat aphid, Sitobion miscanthi. Frontiers in Microbiology 9: 582, https://doi.org/10.3389/fmicb.2018.00582
Revelle, W., 2017., psych: Procedures for psychological, psychometric, and personality research. R Package Version 1.0â95. Available at: https://personality-project.org/r/psych/
Roff, D. A., 2000. Trade-offs between growth and reproduction: An analysis of the quantitative genetic evidence. Journal of Evolutionary Biology 13: 434â445, https://doi.org/10.1046/j.1420-9101.2000.00186.x
Rueden, C. T., Schindelin, J., Hiner, M. C., Dezonia, B. E., Walter, A. E., Arena, E. T., Eliceiri, K. W., 2017. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinformatics 18: 1â26, https://doi.org/10.1186/s12859-017-1934-z
Salem, H., Bauer, E., Strauss, A. S., Vogel, H., Marz, M., Kaltenpoth, M., 2014. Vitamin supplementation by gut symbionts ensures metabolic homeostasis in an insect host. Proceedings of the Royal Society B: Biological Sciences 281: 20141838 https://doi.org/10.1098/rspb.2014.1838
Schmidt, K., Engel, P., 2021. Mechanisms underlying gut microbiota-host interactions in insects. Journal of Experimental Biology 224: 1â16, https://doi.org/10.1242/jeb.207696
Shikano, I., Olson, G. L., Cory, J. S., 2015. Impact of non- pathogenic bacteria on insect disease resistance: Importance of ecological context. Ecological Entomology 40: 620â628, https://doi.org/10.1111/een.12235
Stearns, S. C., 1989. Trade-offs in life-history evolution. Functional Ecology 3: 259â268, https://doi.org/10.2307/2389364
Sturm, R., 2016. Relationship between body size and reproductive capacity in females of the black field cricket (Orthoptera, Gryllidae). Linzer Biol Beiträge 48: 1823â1835.
Sun, J., Tan, X., Li, Q., Francis, F., Chen, J., 2022. Effects of different temperatures on the development and reproduction of Sitobion miscanthi from six different regions in China 10: 794455, https://doi.org/10.3389/fevo.2022.794495
Tsuchida, T., Koga, R., Fukatsu, T., 2004. Host plant specialization governed by facultative symbiont. Science 303: 1989, https://doi.org/10.1126/science.1094611
Wang, Y., Zhu, J., Fang, J., Shen, L., Ma, S., Zhao, Z., Yu, W., Jiang, W., 2020. Diversity, composition and functional inference of gut microbiota in indian cabbage white Pieris canidia (Lepidoptera: Pieridae). Life 10: 254, https://doi.org/10.3390/life10110254
Yang, Y., Joern, A., 1994. Gut size changes in relation to variable food quality and body size in grasshoppers. Functional Ecology 8: 36, https://doi.org/10.2307/2390109
Zakharov, I. A., 2015. Intracellular symbionts as a factor of insect evolution. Biology Bulletin Reviews 5: 99â108, https://doi.org/10.1134/s2079086415020115
Zera, A. J., Harshman, L. G., 2001. The physiology of life history trade-offs in animals. Annual Review of Ecology and Systematics 32: 95â126, https://doi.org/10.1146/annurev.ecolsys.32.081501.114006

{kind=link}
{kind=link}
{kind=link}
{kind=link}