Abstract
Global population growth in both humans and animals is contributing to a shortage of proteins and fats derived from traditional sources such as meat and fish. As a result, alternative food sources such as insects are gaining increased attention. Edible insects are considered an excellent source of nutrients as they contain high-quality proteins, fats and vitamins. In Europe, entomophagy is relatively new and therefore concerns about its safety, potential beneficial health effects and nutritional value need to be considered. In this work, scientific literature was systematically searched using PubMed, Web of Science and Scopus, resulting in 51 studies that examined the effects of edible insects as dietary ingredients on human and mammalian health. Findings suggest that edible insects such as black soldier fly larvae (H. illucens), silkworm (B. mori), yellow mealworm (T. molitor) and field cricket (G. bimaculatus) contain a wide range of bioactive peptides with potential antioxidant, antihypertensive, antimicrobial, immunomodulatory and anti-inflammatory properties. However, entomophagy also entails certain risks, such as allergic reactions and potential contamination with harmful substances that may pose health concerns. A clear understanding of these benefits and risks is crucial for both consumers and industry stakeholders to make informed decisions, guide future research, regulations, and the responsible integration of edible insects into a sustainable food system.
1 Introduction
In recent decades, growing human and animal populations have raised concerns about potential shortages of proteins and fats derived from meat and fish (Jian et al., 2022; Li and Wu, 2023). In human nutrition, animal products are important primary food sources (Lu et al., 2022), whilst for animals, animal by-products (ABPs) – such as bones and meat not intended for human consumption – serve as the primary, highly nutritious source of proteins and fats (Thompson, 2008). Global population growth has increased the demand for meat, stimulating livestock production and consequently, the generation of larger quantities of ABPs during slaughter and processing. These ABPs are vital for feed formulations (Thompson, 2008). However, rising demand for animal proteins also raises concerns about potential shortages and increasing costs affecting both human and animal nutrition. In many industrialised countries, meat consumption has been declining due to health concerns, and, in this context, shifts towards plant-based diets have been observed (Godfray et al., 2018). This trend may reduce the local availability of ABPs, intensifying competition between the human food and pet food sectors for animal-derived proteins. Moreover, livestock production, especially beef, requires significantly more land and emits almost 50 times more greenhouse gas emissions than plant-based alternatives (Cusack et al., 2021). Consequently, the adoption of plant-based proteins in favour of a decline in meat consumption is considered a sustainable environmental strategy, which in turn may further limit ABP supply. Despite these shifts, the capacity of traditional protein sources is unlikely to meet the global demand. Limited agricultural land and adverse climatic conditions make expanding conventional livestock production unsustainable, and therefore, fats and proteins need to be derived from other sources (Jafarzadeh et al., 2024).

Overview of insect species authorised for inclusion in feed for farm animals in Europe. Images adapted from Buy Silkworms for Reptiles: Nutritious Live Food, n.d.; Cricket (Gryllodes sigillatus), BugGuide.Net, n.d.; Gryllus assimilis, BugGuide.Net, n.d.
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Today, many insect species are commonly consumed by various ethnic groups in Africa, Asia and Latin America. However, in Western Europe and the United States, the concept of edible insects remains relatively new (Reed et al., 2021). Concerns regarding their safety, nutritional quality and overall suitability for human consumption persist (Ramos-Elorduy, 2009). In the past, insects were excluded from both human and animal diets due to potential health risks such as the transmission of prion-related diseases (Belluco et al., 2018; German Federal Institute for Risk Assessment et al., 2019). Since May 2017, the EU has authorised processed insect proteins in animal feed (Figure 1). Currently, several insect species are permitted for the use in aquaculture, poultry and pig feed, as well as in pet food (Regulation142/2011, 2011; Regulation 1069/2009, 2009; Regulation 2021/1925, 2021; Thrastardottir et al., 2021). In the United States, the regulatory framework is more limited; black soldier fly (H. illucens) larvae are approved for dog food and aquaculture, while insect-based products intended for human consumption require Generally Recognised as Safe (GRAS) status (Lähteenmäki-Uutela et al., 2021).
Overview of the literature search terms used in this systematic review
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Edible insects are considered an excellent source of nutrients as they contain proteins including essential amino acids, fats including fatty acids, and important micronutrients such as iron, calcium, riboflavin and vitamin B12 (Kouřimská and Adámková, 2016). Moreover, numerous studies have shown that the bioactive compounds in insects may provide health benefits for both humans and animals, such as antioxidant, antihypertensive, anti-inflammatory, and antimicrobial effects (Dai et al., 2013; Jantzen da Silva Lucas et al., 2020; Zielińska et al., 2017). However, potential risks – such as contamination with mycotoxins, bacteria, and chemical residues – may arise from the insects’ environment or feed, the latter typically consisting of organic waste streams to enhance sustainability (De Paepe et al., 2019; Poma et al., 2017). Moreover, edible insects may trigger allergic reactions in some individuals and animals (Premrov Bajuk et al., 2021; Verhoeckx et al., 2014). Finally, uncertainties remain regarding the potential health effects and safety of using insects as a novel, sustainable food and feed source. Therefore, this systematic review aims to provide a comprehensive and balanced overview of the potential health benefits and risks associated with insect consumption in humans and mammals, to better inform consumers, industry stakeholders and policymakers. The focus is primarily on health outcomes and safety considerations, while an in-depth evaluation of nutritional composition and the adequacy of insect proteins to meet complete dietary requirements is beyond the scope of this review.
2 Experimental section
Search strategy and study selection
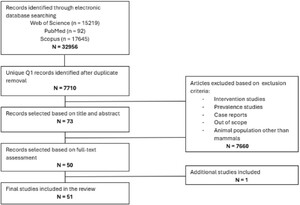
A comprehensive search of English academic literature was conducted across three electronic databases, i.e. PubMed, Web of Science and Scopus. The search focused on articles published between January 2013 and May 2024 in Q1-ranked journals according to the Scimago Journal Rank quartiles (Scimago Journal and Country Rank, n.d.). This inclusion criterium was applied as a methodological quality filter, based on the assumption that Q1-ranked journals are generally associated with more stringent peer-review processes, higher reporting standards and greater methodological rigour. Although it is acknowledged that journal ranking does not guarantee individual study quality and may exclude some relevant studies, it was employed to enhance the overall methodological accuracy. Table 1 presents the core search terms; whereas complete database-specific search strings, including applied filters and syntax, are available in supplementary material (Tables S1–S3 in the Supplementary material). After removing duplicates, the remaining articles were screened for relevance. Selection of relevant studies was conducted in two phases, i.e. an initial screening based on title and abstract, followed by a full-text review of eligible articles for final inclusion. All articles were hand searched for human and animal studies assessing the health-promoting effects and/or risks of consuming edible insects. The screening process was carried out independently by two reviewers (Camila Baptista da Silva and Tiffany De Troyer), and articles were selected based on predefined inclusion and exclusion criteria (Table 2). The study selection followed the PRISMA flowchart (Figure 2), and the study protocol was registered in the PROSPERO registry (CRD42024550089).
Inclusion and exclusion criteria used in this systematic review
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
PRISMA flow chart of the systematic review conducted in this work.
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Data extraction and quality assessment
After the screening process, relevant data such as sample size, study population characteristics (species, age and sex), biological matrix, and major findings were extracted from the full-text articles. The quality of the studies was assessed using the Systematic Review Centre for Laboratory Animal Experimentation (SYRCLE) risk of bias tool or the Newcastle-Ottawa Scale (Gierisch et al., 2014; Hooijmans et al., 2014). More specifically, for case-control studies and cohort studies, the Newcastle-Ottawa Scale was applied, evaluating three key domains: selection (0–4 points), comparability (0–2 points), and outcome assessment (0–3 points). Mammalian studies were assessed using SYRCLE’s risk of bias tool, covering four domains, i.e. selection (0–3 points), detection (0–2 points), performance (0–2 points), and other biases (0–3 points). A higher total score for both the SYRCLE risk of bias tool and the Newcastle–Ottawa Scale indicates better study quality. Studies with a quality score lower than 6 were excluded from this review. Common sources of bias identified during the assessment included inadequate randomisation (selection bias), lack of blinding of investigators or participants (performance and detection bias) and incomplete or selective outcome reporting (reporting bias). These biases can substantially influence study validity, e.g. poor randomisation may lead to systematic differences between groups, while lack of blinding can introduce measurement errors or subjective interpretations of outcomes. Similarly, selective reporting may overestimate treatment effects. Recognising these biases was essential for ensuring that only studies with sufficient methodological accuracy were included in this review. Quality assessment was performed independently by two reviewers (Camila Baptista da Silva and Tiffany De Troyer). In case of disagreement, a third reviewer (Lieselot Hemeryck) was consulted. More detailed information regarding quality assessment is provided as supplementary information (Tables S4 and S5 in the Supplementary material).
Literature screening and study characteristics
The initial search yielded 32 956 articles. After removing duplicates and screening for relevance, 54 full-text articles met the inclusion criteria. Three studies were excluded due to low methodological quality, as assessed by the SYRCLE’s Risk of Bias tool and the Newcastle-Ottawa Scale, leaving 51 studies for further analysis. Of these, two involved healthy human participants, while the remaining 49 were conducted in other mammals. Specifically, healthy populations were studied in dogs (n = 3), cats (n = 3), and pigs (n = 18). In contrast, all rodent studies, in mice (n = 10) and rats (n = 15), except for one, were conducted in disease models. Blood was the most frequently analysed biological matrix (n = 42), followed by faeces (n = 15), urine (n = 4), caecal content (n = 2), colon digesta (n = 2), brain tissue (n = 1) and foetal tissue (n = 1).
The most studied insect species were the black soldier fly (H. illucens, n = 21), silkworm (B. mori, n = 12), and yellow mealworm (T. molitor, n = 9). Other insects investigated included two-spotted cricket (G. bimaculatus, n = 3), house cricket (A. domesticus, n = 2), lesser mealworm (A. diaperinus, n = 3), housefly (M. domestica, n = 2), superworm (Z. morio, n = 2), Japanese rhinoceros beetle (A. dichotoma, n = 2), rose chafer beetle (C. pilifera, n = 1), Japanese burrowing cricket (V. aspersus, n = 1), rice grasshopper (O. c. sinuosa, n = 1), speckled cockroach (N. cinerea, n = 1), Madagascar hissing cockroach (G. portentosa, n = 1), white-spotted flower chafer (P. brevitarsis, n = 1), tropical house cricket (G. sigillatus, n = 1) and two-lined velvet hawkmoth (C. bilineata, n = 1). One study investigated the health effects of insect tea – a beverage derived from the excrement of insects, mainly the repugnant azeta moth (H. repugnalis), which feeds on the leaves of Ilex kudingcha – using a mouse model. Most studies were conducted with whole meal (n = 35), followed by hydrolysates (n = 5), defatted fractions (n = 3), oil fractions (n = 2), protein extract (n = 2), sericin water extract (n = 2), ethanol extract (n = 1), chitooligosaccharides (n = 1) and tea polyphenol (n = 1). More detailed information on the included studies is presented in Table S6 in the Supplementary material.
3 Reported health-promoting effects
Gut microbiota modulation and gut health
Summary of included studies investigating the effects of insect-based diets on gut microbiota composition and fermentation end-products across mammalian species
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Seventeen studies reported outcomes related to microbiota modulation (Table 3). The majority were performed in pigs (n = 10), followed by rodents (n = 3), cats (n = 2), dogs (n = 1), and humans (n = 1). Most studies used H. illucens (n = 12), though other species such as T. molitor (n = 2), G. bimaculatus (n = 2), A. diaperinus (n = 1), A. domesticus (n = 1), B. mori (n = 1), N. cinerea (n = 1), G. portentosa (n = 1), and Z. morio (n = 1) were also investigated.
In healthy humans, Stull et al. (2018) found increased Bifidobacterium animalis and decreased Lactobacillus reuteri following a two week-intervention with 25 g/day of G. sigillatus powder. SCFA levels declined (acetate, propionate), and microbial diversity remained unaltered. However, the uncontrolled background diet likely limited conclusions.
In healthy pigs, H. illucens larvae supplementation at moderate levels (1–12%) increased Lactobacillus and Bifidobacterium genera (Jin et al., 2021; Phaengphairee et al., 2023; Yu et al., 2019; Yu et al., 2020), and elevated SCFA concentrations, particularly butyrate, though effects on branched-chain fatty acids (BCFA) were inconsistent, with increases reported by Yu et al. (2019) and decreases by Malla et al. (2024). These findings were often accompanied by reduced Escherichia coli, biogenic amines, phenols, and indoles. However, other trials showed no significant differences in microbial profiles or diversity (Håkenåsen et al., 2021; Malla et al., 2024; Spranghers et al., 2018; Zacharis et al., 2023), or an increase in potentially harmful bacteria like Staphylococcus and Chlamydia genera (Biasato et al., 2020). Liu et al. (2023) observed that a 25% H. illucens larvae inclusion yielded higher SCFA and more favourable microbiota (e.g. Cyanobacteria, Campilobacteriota, Megasphaera) than higher levels (up to 100% replacement of soybean meal), which negatively impacted piglet performance and microbial profiles (e.g. Staphylococcus), possibly due to limited chitin-degrading capacity.
In healthy cats, Bosch et al. (2024) reported that 37.5% H. illucens larvae inclusion increased SCFA, BCFA and biogenic amine (e.g. putrescine, cadaverine, histamine) production, and fermentative bacteria, such as Bifidobacterium and Megasphaera, while alpha diversity was reduced. Interestingly, the H. illucens larvae diet had a lower protein content and lower apparent digestibility than the control diet, suggesting that more undigested protein may have reached the colon, thereby increasing proteolytic fermentation and associated biogenic amine production. In contrast, Reilly et al. (2022), using a 4% inclusion of insect meals from N. cinerea, G. portentosa, and Z. morio larvae in retorted cat food, found no significant differences in microbial diversity, faecal metabolites (ammonia, phenols, indoles, SCFA, and BCFA) or digestibility compared to a chicken-based control, though some minor changes in taxa (e.g. Bacteroidaceae) were noted. In dogs, Areerat et al. (2023) tested diets containing 10–20% A. domesticus or 7–14% B. mori pupae and found no substantial shifts in major phyla or diversity. Some increases in Lactobacillus and Corynebacterium were observed, particularly with B. mori, potentially due to the presence of chitin in the tested diets.
In rodents, H. illucens larvae oil inclusion (6.8%) improved microbial α- and β-diversity and reduced Clostridiaceae in obese rats (Marschall et al., 2024). In contrast, no major microbial shifts were reported in healthy rats fed A. diaperinus (Lanng et al., 2021). Feeding colitis-induced mice H. illucens larvae that had been previously fermented with Lactobacillus plantarum improved beneficial intestinal bacteria, including Bacteroides caccae and Akkermansia muciniphila (Son et al., 2023). However, as the study did not include a non-fermented H. illucens as control, it remains unclear if these observations were a result of the fermentation process, H. illucens itself or their combination.
Despite promising findings (improvement in fermentative bacteria and their related end-products), reported changes in SCFA, BCFA and biogenic amines production remain inconsistent across species, study designs, and insect types, while ammonia levels were mostly unaffected. These heterogeneous fermentation outcomes can be explained by the complex interactions between insect-derived polysaccharides and the gut microbiota, which have indeed been reported. Chitin and its derivative, chitosan, are resistant to mammalian digestive enzymes and therefore reach the colon largely intact (Tabata et al., 2018), where they can act as fermentable substrates for specific microbial taxa. Chitin and chitosan fermentation may directly contribute to SCFA production or indirectly promote cross-feeding interactions that favour butyrate- and propionate-producing bacteria. In turn, microbially derived metabolites such as SCFA and BCFA can influence host responses, including modulation of intestinal barrier function (Ferri et al., 2024).
Lipid and glucose metabolism
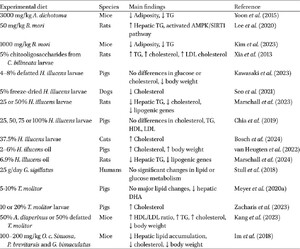
Sixteen studies addressed lipid and/or glucose metabolism, of which most were conducted in rodents (n = 8), followed by pigs (n = 5), cats (n = 1), dogs (n = 1), and humans (n = 1). The majority used H. illucens (n = 7), though other species such as T. molitor (n = 3), G. bimaculatus (n = 2), B. mori (n = 2), and others (n = 6) were also tested.
In humans, Stull et al. (2018) conducted a 14-day crossover study with 25 g/day cricket powder but found no significant changes in lipid or glucose metabolism. The short duration and lack of dietary control were noted as limitations.
In pigs, most studies report no significant alterations in lipid profiles. For instance, Meyer et al. (2020a) observed no changes in hepatic or plasma lipids in pigs fed 5–10% T. molitor meal, although a reduction in phospholipid desaturation and hepatic docosahexaenoic acid (DHA) content was noted. Chia et al. (2019) also found no differences in cholesterol, triglycerides, HDL, or LDL with up to 100% fish meal replacement by H. illucens larvae meal. Similarly, Kawasaki et al. (2023) observed no differences in glucose or cholesterol in piglets receiving up to 8% defatted H. illucens larvae, although a decrease in body weight was reported. In contrast, Zacharis et al. (2023) reported increased serum cholesterol in pigs fed T. molitor reared on a substrate partially enriched with aromatic plant residues, but not in pigs fed the same species reared on a standard substrate, highlighting that rearing substrate may significantly influence metabolic outcomes. van Heugten et al. (2022) found a linear increase in serum cholesterol with 2–6% H. illucens larvae oil inclusion. However, this was accompanied by increased body weight and improved feed efficiency, suggesting that the rise in cholesterol may have been influenced by metabolic changes associated with weight gain rather than solely by the dietary fat source (Li et al., 2012).
In dogs, Seo et al. (2021) found that inclusion of 5% H. illucens larvae for 12 weeks reduced serum cholesterol, potentially due to the high lauric acid content of the larvae (328 g/kg), which has been shown to modulate hepatic cholesterol metabolism (Wood and Migicovsky, 2011). Controversially, in cats, Bosch et al. (2024) reported increased cholesterol with 37.5% H. illucens larvae after 28 days of dietary treatment, though no adverse health effects were noted. While both diets had similar fat content and digestibility, the H. illucens diet had a lower protein content, which may have influenced lipid metabolism and contributed to the observed cholesterol increase.
In rodents, most studies used diet-induced obese models (n = 3), high-fat diets (n = 3), or fatty liver disease models (n = 2). In obese Zucker rats, partial replacement (25–50%) of casein with H. illucens protein dose-dependently reduced hepatic triglycerides and cholesterol, as well as the expression of lipogenic genes (e.g. Fasn, Scd1, and Fads1) (Marschall et al., 2023). Plasma cholesterol also decreased in a dose-dependent manner, while triglycerides and body weight remained unchanged, suggesting lipid improvements occurred independently of weight. In a 28-day study with obese rats, 6.9% H. illucens oil reduced hepatic triglycerides and downregulated hepatic mRNA concentrations of three genes involved in lipogenesis (Fasn, Gpam, and Hmgcr) compared to palm oil, but not soybean oil, while plasma triglycerides and cholesterol were unaffected (Marschall et al., 2024).
In high-fat diet-induced obese mice, Kang et al. (2023) showed that T. molitor and A. diaperinus protein (50–100% casein replacement) improved serum HDL/LDL ratios and modulated genes linked to lipid and glucose metabolism (e.g. Srebf1, Dgat2, Slc2a1). However, plasma triglyceride and cholesterol levels remained elevated compared to lean controls, while body weight reductions were noted. Mixed results were observed in healthy rodents fed a high-fat diet. Xia et al. (2013) demonstrated that a dietary inclusion of 5% chito-oligosaccharides from C. bilineata larvae improved lipid profiles (triglycerides, cholesterol, and low-density lipoprotein cholesterol) and increased faecal fat and cholesterol excretion. Similarly, 1 g/kg of B. mori (Kim et al., 2023) and 3000 mg/kg of A. dichotoma powder (Yoon et al., 2015) reduced adiposity and serum triglycerides in mice fed a high-fat diet, effects that were associated with reduced feed intake and/or decreased feed conversion efficiency. In fatty liver disease models, insect-derived compounds consistently showed hepatoprotective effects, though typically at pharmacological doses (50–200 mg/kg body weight per day); thus, effects may not translate directly to whole-insect foods or other mammals without dose normalisation. For instance, Im et al. (2018) found that extracts from G. bimaculatus, O. c. sinuosa, and P. brevitarsis (100-200 mg/kg body weight per day) reduced hepatic lipid accumulation, lowered serum cholesterol, and downregulated genes important for hepatic lipid droplet accumulation and free fatty acid uptake in a dose-dependent manner, suggesting enhanced fatty acid oxidation. However, body weight reductions were also noted. Similarly, Lee et al. (2020) reported that 50 mg/kg body weight of B. mori powder attenuated ethanol-induced steatosis by activating the AMPK/SIRT1 pathway and reducing hepatic triglycerides.
Overview of included studies evaluating the impact of insect-based diets on lipid and glucose metabolism in humans, livestock, companion animals and rodent models
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
A summary of lipid and glucose metabolism findings across studies is presented in Table 4. Overall, insect-based diets, particularly those incorporating H. illucens larvae or B. mori, show promising potential to modulate lipid metabolism, especially in rodent models of metabolic dysfunction. Although mechanistic evidence is limited, the proposed mechanisms include the combined effects of bioactive compounds. Medium-chain fatty acids may enhance hepatic fatty acid oxidation and reduce cholesterol synthesis, while bioactive peptides can modulate the expression of genes involved in lipid and glucose metabolism (Cao et al., 2024; Liu et al., 2017). Chitin and chitosan promote the production of SCFA by the gut microbiota, which may improve lipid metabolism and insulin sensitivity (Figure 3) (Bosch et al., 2024; Marschall et al., 2023, 2024; Meyer et al., 2020a,b; Seo et al., 2021; van Heugten et al., 2022). However, it is important to note that the changes in lipid markers were often accompanied by alterations in body weight, making it difficult to distinguish whether the metabolic changes were directly attributable to insect-derived components or secondary to weight fluctuations. Nonetheless, even at pharmacological dosing, these results are encouraging and provide proof-of-concept that warrants testing of food-realistic inclusion levels and standardised processing in humans and companion animals. On the other hand, the lipid effects were more variable in pigs, and evidence in humans and companion animals remains limited.
Antioxidant, anti-inflammatory effects and immune modulation
Visualisation of the potential effects of edible insect consumption on human and mammalian health. Edible insects may provide bioactive peptides with antimicrobial, antioxidant and antihypertensive activities. They may modulate gut-microbiota-derived metabolites and impact pathways related to oxidative stress, immune modulation and glucose and lipid metabolism in the liver. Green arrows indicate the potential positive effects of insect consumption. SCFA, short-chain fatty acids; BSCFA, branched-chain fatty acids; TNF, tumor necrosis factor; IL, interleukin; ROS, reactive oxygen species; SOD, superoxide dismutase; CAT, catalase; GSH, glutathione; 8-OH-dG, 8-hydroxydeoxyguanosine; MDA, malondialdehyde; 4-HNE, 4-hydroxynonenal; ACE, angiotensin converting enzyme; hk, hexokinase; gpi, glucose phosphate isomerase; pfkm, phosphofructokinase; pdh, pyruvate dehydrogenase; cs, citrate synthase; acly, ATP-citrate lyase; acc, acetyl-CoA carboxylase; fasn, fatty acid synthase; scd, stearoyl-CoA desaturase; fads, fatty acid desaturase; dgat, diacylglycerol O-acetyltransferase; lpl, lipoprotein lipase; pksG, polyketide synthase G.
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Summary of included studies investigating the antioxidant, anti-inflammatory and immunomodulatory effects of insect-derived ingredients on mammalian species
Citation: Journal of Insects as Food and Feed 2026; 10.1163/23524588-bja10384
Thirteen studies explored the antioxidant, anti-inflammatory, and/or immunomodulatory effects of insect-derived ingredients in pigs (n = 5), rodents (mice n = 3; rats n = 2), dogs (n = 1), cats (n = 1), and humans (n = 1). Frequently examined insect species were H. illucens (n = 7), followed by B. mori (n = 2), crickets (A. domesticus, G. bimaculatus; G. sigillatus n = 3), and others (n = 3). While most studies involved healthy animals, some murine models targeted disease states, such as metabolic dysfunction-associated steatotic liver disease (MASLD), colitis, and ethanol-induced injury. An overview of these outcomes is presented in Table 5.
In a short-term (2-week) crossover trial in healthy humans, Stull et al. (2018) reported that 14 days of G. sigillatus consumption (25 g/day) reduced plasma tumour necrosis factor alpha (TNF-α), indicating mild anti-inflammatory effects. However, the diets were not isocaloric or isonitrogenous, limiting comparability.
Among the studies conducted in healthy pigs, Boontiam et al. (2022) showed that replacing 5% of fish meal with G. bimaculatus improved antioxidant enzymes (increased superoxide dismutase (SOD), glutathione peroxidase (GPx)), and reduced lipid peroxidation (malondialdehyde (MDA)) and pro-inflammatory cytokines (interleukin-6 (IL-6), TNF-α). Yu et al. (2019) reported that a 4% inclusion of H. illucens larvae reduced intestinal inflammation markers (toll-like receptor 4, interferon gamma), increased anti-inflammatory IL-10, and improved gut barrier integrity (increased tight junction proteins zonula occludens-1 and occludin). These effects were attributed to increased levels of butyrate and short-chain fatty acid (SCFA)-producing bacteria, as well as a reduction of total amines, cadaverine, tryptamine, phenol, p-cresol, and skatole. However, a further increase to 8% of H. illucens inclusion had no additional benefit, potentially due to higher chitin or non-protein nitrogen, which was hypothesised to reduce nutrient availability or affect gut microbiota. Phaengphairee et al. (2023) observed that 12% H. illucens larvae alone reduced MDA and IL-1β, but had minimal effects on inflammation and antioxidant enzymes (SOD, GPx, TNF-α, IL-6, immunoglobulin G (IgG), or total antioxidant capacity (TAC)), though co-administration with probiotics elicited broader immunomodulatory effects (reduced MDA, IL-1β and TNF-α; increased IgG and SOD). Liu et al. (2023) reported elevated ileal IgM and IgG levels in piglets fed diets containing 25% H. illucens larvae. Similarly, Cho et al. (2023) observed increased serum IgG and IgA concentrations in piglets where 3% of dietary protein was replaced with T. molitor. Inflammatory markers were not evaluated, however.
In healthy companion animals, Seo et al. (2021) found no significant change in IL-10, TNF-α, or IgG in dogs fed 5% H. illucens larvae vs 10% fermented oatmeal for 12 weeks. In contrast, Li et al. (2024) showed that cats fed a protein blend of H. illucens larvae – Schizochytrium algae (10–15% inclusion) increased SOD and GPx, reduced MDA, and decreased IL-1β and IL-8 levels. However, the absence of H. illucens larvae-only or Schizochytrium algae-only group made it unclear whether the effects were driven by insect-derived components or the omega-3-rich algal inclusion.
In rodent disease models, several insect-derived compounds demonstrated notable antioxidant and anti-inflammatory effects. In murine models of MASLD, supplementation with insect extracts or peptides consistently attenuated hepatic inflammation. For instance, Im et al. (2018) showed that G. bimaculatus, O. c. sinuosa, and P. brevitarsis extracts (100–200 mg/kg body weight per day) reduced hepatic TNF-α, IL-1β, and IL-6 expression in mice with high-fat induced MASLD. Similarly, 100 mg/kg of a multi-insect-derived peptide (AGL9 derived from larvae from A. dichotoma, M. domestica, C. pilifera, T. molitor, H. illucens, B. mori, Orthoptera, and V. aspersus) restored hepatic antioxidant enzyme expression (increased SOD and GPx) and suppressed pro-inflammatory cytokines (IL-1β, TNF-α, IL-6) via AMP-activated protein kinase/nuclear factor erythroid 2–related factor 2 pathway (AMPK/Nrf2) activation (Fan et al., 2021). In a model of ethanol-induced hepatic damage, effects were mitigated by B. mori powder at 50 mg/kg body weight (Lee et al., 2020) with a reduction of oxidative stress markers (e.g. 8-hydroxy-2’-deoxyguanosine, MDA) and inflammatory mediators (e.g. TNF- α, IL-1β). In an ethanol-induced gastric injury model, B. mori also conferred gastroprotective effects: enhanced mucosal integrity, reduced lipid peroxidation (MDA), restored antioxidant capacity (TAC), and suppressed cytokine expression (TNF-α, IL-1β, cyclooxygenase-2, inducible nitric oxide synthase), potentially via inhibition of mitogen-activated protein kinase and protein kinase B signalling (Yun et al., 2017). In colitis-induced mice, fermented H. illucens with Lactobacillus planta reduced inflammatory cytokine expression (TNF-α, IL-1β, IL-6) and disease severity (Son et al., 2023), although the fact that the H. illucens was fermented made it difficult to attribute the effects solely to the crude insect.
Collectively, these findings suggest that insect-derived ingredients/extracts may exert anti-inflammatory and antioxidant effects across both healthy and diseased humans and animals, by modulating cytokine expression (e.g. TNF-α, IL-1β, IL-6, IL-10) and redox balance (e.g. SOD, GPx, MDA, TAC).
Antihypertensive effects
Only two studies evaluated the antihypertensive potential of insect-derived compounds in a hypertensive rat model. Anuduang et al. (2024) supplemented spontaneously hypertensive rats with 50, 100, and 200 mg/kg body weight per day of crude mature B. mori larvae hydrolysate. Medium and high doses significantly reduced both systolic/diastolic blood pressure by 60/49 and 50/40 mmHg, respectively, compared to baseline. These reductions were dose-dependent and comparable to those achieved with captopril (25 mg/kg body weight per day), although captopril resulted in the lowest overall blood pressure. Similarly, Stella et al. (2022) fed hypertensive rats a diet supplemented with 4.5% T. molitor protein extract (approx. 2.9 g/kg body weight per day), which prevented the rise in systolic blood pressure observed in untreated hypertensive controls. The antihypertensive effect only became evident after 4 weeks and was comparable to captopril (8 mg/kg body weight per day) by the study’s end. In both studies, the antihypertensive effects were attributed to potential angiotensin-converting enzyme (ACE) inhibition, with additional involvement of nitric oxide pathways, angiotensinogen regulation, or fatty acid metabolism. While captopril showed more immediate action, insect-derived peptides appeared to act more gradually, suggesting a sustained physiological modulation.
Memory effects
Research on the memory-enhancing effects of edible insects is limited, with only two studies, i.e. one in humans and one in a cognitive impairment mouse model, both investigating the effects of B. mori-derived bioactive compounds. In a placebo-controlled, double-blind human trial, Kang et al. (2018) evaluated the effects of fibroin enzymatic hydrolysate from B. mori on cognitive performance over three weeks. Participants consumed 0, 280, 400, or 600 mg/day, with significant improvements in verbal memory recall and attention observed at 400 and 600 mg/day, but not at the lower dose. These findings suggest a dose-dependent effect, possibly mediated by neuroprotective peptides, though no neurochemical markers were assessed to confirm the mechanisms involved. Complementary findings were reported by Nguyen et al. (2021), who evaluated memory-enhancing effects in a scopolamine-induced mild cognitive impairment mouse model using mature B. mori powder, extract of the Korean angelica plant, and their combinations. While the B. mori powder and the Korean angelica extract alone showed moderate benefits, the combined treatment (particularly at a 1:0.8 ratio) led to the greatest improvements in spatial, social, and fear-conditioned memory, suggesting potential synergism. These effects were linked to increased brain mitochondrial activity, elevated ATP levels, and reduced acetylcholinesterase activity.
Synthesis of health effects and translational insights
Across the included studies, most studies documented anti-inflammatory and antioxidant effects in response to the intake of H. illucens and B. mori (e.g. reductions in TNF-α, IL-1β, and IL-6, alongside higher SOD/GPx and lower MDA), with shifts (increased faecal SCFAs and enrichment of Lactobacillus/Bifidobacterium) pointing towards higher microbial fermentation when feeding H. illucens. Conversely, metabolic outcomes remain heterogeneous: rodent models often show favourable hepatic lipid markers (lower hepatic triglycerides and downregulated lipogenic genes), whereas the results of companion-animal and pig studies vary widely. Differences may relate to factors such as the fraction type (e.g. oil, protein, whole, defatted), inclusion level, rearing substrate, and study duration, but the limited number of studies per fraction and great variation in studied endpoints preclude firm conclusions. For example, defatted meals proportionally contain more protein but fewer bioactive lipids, whereas hydrolysates consist of small peptides with distinct functional activity, and oils lack both chitin and protein, indicating that different fractions derived from the same insect species may elicit very different physiological effects (Meyer-Rochow et al., 2021). Preliminary findings on hypertensive models and memory-enhancing effects are promising; however, these are based on only two studies each, typically using insect extracts rather than whole fractions. Translational relevance to real dietary intake is therefore limited, and further research, including studies in additional animal models, is warranted.
Several insect-derived components (e.g. lauric acid and chitin) are hypothesised to contribute to these effects. For instance, lauric acid has been linked to lipid modulation, potentially reducing cholesterol (Hashim et al., 1960; Seo et al., 2021; Wood and Migicovsky, 2011), whereas chitin has been linked to microbiota modulation (Gasco et al., 2020). However, across reviewed studies, these compounds were not isolated, and reporting of nutritional composition was inconsistent, limiting causal inference. Discrepancies further underscore this complexity: some H. illucens larvae studies report neutral or increased cholesterol despite its high inclusion levels (Kawasaki et al., 2023; van Heugten et al., 2022; Zacharis et al., 2023). Similarly, chitin-associated microbial shifts vary across studies. For instance, Lactobacillus abundance increased in pigs fed H. illucens (Jin et al., 2021; Phaengphairee et al., 2023; Yu et al., 2019; Yu, et al., 2020) and dogs fed B. mori (Areerat et al., 2023), whereas decreases were reported in pigs fed H. illucens and T. molitor (Håkenåsen et al., 2021; Zacharis et al., 2023), and in humans consuming Gryllus spp. (Stull et al., 2018). The variability in microbial shifts may reflect differences in insect fraction, inclusion level, host physiology, baseline microbiota, and methodological differences. Taken together, these findings highlight the potential health-promoting properties of edible insects, which are summarised in Figure 3.
4 Potential hazards and risks
Safety and toxicity
Only four rodent studies included in this review addressed subchronic toxicity of insect-based ingredients over durations spanning 70 to 90 days, providing valuable but limited insight into safety at high dosages.
Kim et al. (2020) and Machona et al. (2024) administered high daily doses of freeze-dried Z. atratus (1250–5000 mg/kg) and T. molitor larvae powder (up to 3000 mg/kg) to rats for 90 and 70 days, respectively. Neither study reported overt toxicity, and although minor liver and kidney variations were observed, these remained within physiological norms and showed no significant differences across groups. Li et al. (2013) evaluated dietary inclusion of M. domestica larval extracts (13.2–33 g/kg body weight per day) over 13 weeks in Sprague Dawley rats, noting slight increases in male liver weight but no histopathological damage or adverse biochemical effects, thus supporting a no-observed-adverse-effect-level (NOAEL) at the highest dose tested. Li et al. (2022) studied B. mori sericin extract (250–1000 mg/kg body weight per day) in pregnant rats and reported no maternal, foetal, or developmental toxicity, establishing a NOAEL at 1000 mg/kg body weight. An exception arose in Jeong et al. (2013), where rats fed a fungal-insect complex consisting of Paecilomyces sinclairii colonising B. mori larvae (5000–50 000 ppm) exhibited dose-dependent renal tubular hypertrophy and elevated kidney injury biomarkers (kidney injury molecule-1, cystatin C,
Collectively, the obtained evidence indicates that edible insects are well tolerated in mammals under standard conditions, with no clear signs of systemic toxicity. Nonetheless, the nephrotoxicity effects observed by Jeong et al. (2013) suggest that fungal contamination of insects may pose a health risk.
Although it was not within the scope of this paper to review health risks posed by contaminants, it is important to note that chemical contaminants, particularly heavy metals, pose an additional safety risk. Several monitoring studies have demonstrated the presence of arsenic, cadmium, mercury and lead in insect-based products, albeit generally in low concentrations, comparable to those found in conventional animal-derived foods (Aguilar-Toalá et al., 2022; Kolakowski et al., 2021; Poma et al., 2017). Comprehensive screening of multiple farmed insect species has shown that when insects are reared under controlled conditions with regulated diets, contaminants like heavy metals remain below established safety thresholds (De Paepe et al., 2019). Importantly, there is evidence that the occurrence and extent of chemical contamination and bioaccumulation in insects are strongly influenced by rearing substrates and production practices used, with contaminants likely originating from feed materials used during rearing (Meyer et al., 2021).
Allergic reactions
The potential for allergic reactions to edible insects is a critical safety consideration, particularly due to cross-reactivity with known allergens such as crustaceans and dust mites. Several insect proteins, including tropomyosin and arginine kinase, have been identified as major IgE-binding allergens. Structural similarities with homologous proteins in crustaceans and dust mites can trigger allergic reactions in sensitised individuals (Aguilar-Toalá et al., 2022; Broekman et al., 2016, 2017; Liu et al., 2009).
In this review, only three controlled studies were retained that assessed insect allergenicity: two in rodents and one in dogs. In dogs, Areerat et al. (2021) tested diets containing 10–20% A. domesticus or 7–14% B. mori pupae for 28 days and found higher eosinophil levels in all insect groups compared to the control group, although this was not statistically significant. Additionally, no allergic signs were observed in dogs.
Dai et al. (2023) evaluated the allergenic potential of B. mori pupa hydrolysates in mice exposed via different immunisation routes, comparing intact (80–30 kDa) and enzymatically hydrolysed B. mori (500 Da). The study found that whole B. mori proteins induced strong allergic responses, such as wheezing, tremors, elevated Th2-cytokines (IL-4, IL-5, IL-13), and high IgG/IgG1 titers, particularly following intraperitoneal sensitisation, while enzymatic hydrolysis reduced allergenicity. This suggests that protein processing may play a key role in modulating immune responses. Additionally, Kim et al. (2020) assessed subchronic toxicity in rats fed freeze-dried Z. atratus larvae powder (1250–5000 mg/kg of body weight/day) for 13 weeks, reporting no overt allergic reactions and no significant changes in IgE concentrations.
Overall, these findings suggest that both protein composition and processing methods influence the allergenic potential of edible insects. Processing strategies, such as enzymatic hydrolysis, can reduce IgE binding and clinical symptoms, whereas heat treatments may not fully eliminate allergenicity (Aguilar-Toalá et al., 2022; Dai et al., 2023).
Limitations of this review
Despite growing regulatory approval for insect-based ingredients in food and feed, several limitations constrain the current evidence base. First, most studies have been conducted in rodent (n = 25) or porcine models (n = 18), with relatively fewer trials in companion animals (n = 6) or humans (n = 2), raising concerns about the applicability of findings to real-world human and pet health. In particular, porcine studies often focus on growth and nutrient efficiency rather than health-specific endpoints (Biasato et al., 2020; Yu et al., 2019), further limiting comparability across outcomes. Human evidence is extremely limited. The two included trials were short-term (14 days and 3 weeks), with a small number of samples, using specific interventions (whole cricket powder at breakfast vs purified B. mori fibroin hydrolysate) (Kang et al., 2018; Stull et al., 2018). Diets were not fully isonitrogenous/isocaloric or lacked mechanistic biomarkers, restricting generalizability to typical diets and long-term outcomes. Second, most studies were short-term (2-4 weeks), of which five were only two weeks (Hwang et al., 2019; Malla et al., 2024; Spranghers et al., 2018; Stull et al., 2018; Yun et al., 2017). For example, of these studies, three assessed gut microbiota (Stull et al., 2018; Spranghers et al., 2018; Malla et al., 2024), with only one reporting significant changes. While studies of 2-4 weeks may capture early dietary responses, some effects may require longer adaptation periods, so short trials can miss both beneficial and adverse outcomes. Therefore, short study durations might limit conclusions about sustained health effects, and longer-term studies are still warranted.
Third, the diversity of insect species and inclusion levels used across studies introduces high variability in nutrient composition and bioactive content, which complicates cross-comparison and generalisation of results. This is further influenced by differences in rearing substrates and environmental conditions (Zacharis et al., 2023), which may affect both efficacy and safety. Processing methods add additional variability: defatting, hydrolysis, and thermal treatments can e.g. alter nutrient availability, bioactive peptide profiles, and allergenic potential (see Meyer-Rochow et al. (2021) for a detailed review). While most studies used whole insect meals, only a small number investigated oils, hydrolysates, defatted fractions, or purified components, and these studies targeted diverse endpoints. Moreover, inclusion levels varied widely across studies, ranging from low dietary incorporations to pharmacological doses, further limiting the possible comparison of dose–response relationships or translating findings to realistic intake scenarios. This uneven representation across fraction types and dosing schemes constrains robust fraction-specific or intake-relevant interpretations and contributes to the heterogeneity observed across outcomes.
Fourth, although no severe toxicological effects were reported, safety data remain incomplete. Mild hepatic and renal changes observed in high-dose animal studies warrant caution (Jeong et al., 2013), especially for sensitive populations (e.g. with renal diseases). To date, no studies have combined subchronic toxicity assessment with health outcomes, and robust clinical trials in humans remain lacking. Moreover, long-term studies in humans and companion animals are lacking, leaving uncertainty around cumulative exposure and potential risks such as heavy metal accumulation or microbial contamination, although this is most likely of relevance (Ng’ang’a et al., 2021).
Finally, the design heterogeneity and lack of standardised methodologies limit cross-study comparability. Most microbiota studies relied on 16S ribosomal RNA gene sequencing (n = 13), though workflows and DNA extraction kits varied; a few studies used alternative approaches such as qPCR (n = 3) or agar-plate culturing (n = 3). Importantly, 16S sequencing provides taxonomic composition but not functional insights (Suchodolski 2021), highlighting the need for metagenomic or metabolomic analyses in future research. Although SCFA quantification was most often done using gas chromatography (n = 7) or liquid chromatography (n = 2), these assays are not inherently comparable across laboratories given differences in sample preparation (e.g. extraction, derivatisation, internal standards), chromatographic conditions (columns, temperature programs), and calibration strategies. Cytokine and antioxidant profiles were measured by enzyme-linked immunosorbent assay, but with assays mostly from different manufacturers. As these differences may affect absolute values, the direction of change rather than absolute concentrations is more relevant for comparison.
Future perspectives
To better understand the health effects of edible insects, future research should focus on understanding specific functional components, such as chitin or insect-derived fatty acids. Isolating and testing these compounds individually may help clarify their specific roles and mechanisms of action. However, given the complexity of whole-food matrices and potential synergistic effects, isolated compound studies should be complemented by whole-diet models to ensure translational relevance. Long-term intervention studies in target populations are needed to evaluate sustained health outcomes. These should explore dose–response relationships, clarify biological mechanisms, and account for how insect rearing conditions affect nutritional composition and bioactivity. Safety remains a critical area for future work. Chronic toxicity studies, the establishment of safe upper intake levels, and deeper investigation into organ-specific risks are essential to support regulatory frameworks and ensure consumer protection. Additionally, allergic responses require further investigation. Although severe allergic reactions have not been widely reported, the potential for cross-reactivity, species-specific immune responses, and the influence of protein processing on allergenicity remains a concern. Future studies should focus on assessing immune responses in at-risk individuals and pets, and evaluate how processing methods (e.g. hydrolysis) influence allergenic potential. Standardised testing in humans and controlled trials in companion animals are considered essential to better understand and manage these risks.
5 Conclusion
This review highlights the multifaceted health implications of insect consumption across mammals, covering antioxidant and anti-inflammatory responses, immune modulation, metabolic regulation, and potential cognitive benefits. Overall, insect-derived proteins and lipids appear to exert favourable effects on lipid profiles, oxidative status, and immune parameters, particularly in animal models of metabolic dysfunction or inflammatory conditions. In addition, some studies reported improvements in blood pressure and cognitive performance, suggesting potential neuromodulatory and cardiovascular benefits. Mechanistically, the observed beneficial effects may be attributed to bioactive compounds present in edible insects (such as chitin and lauric acid), alongside their ability to modulate the gut microbiota. Nonetheless, these pathways remain incompletely characterised. Safety outcomes generally support the use of edible insects in animal and human diets, with no severe adverse effects reported in the reviewed subchronic toxicity studies. Nevertheless, signs of mild hepatic and renal alterations at high doses, as well as potential immunogenicity, depending on processing methods, underscore the need for more robust allergenicity and safety assessments. Taken together, while insect-based ingredients hold significant promise for promoting health and sustainability in mammalian diets, current evidence calls for cautious optimism. Well-designed studies addressing the variability, safety, and functional effects of insect consumption are essential to be able to fully harness their potential as a novel food and feed source.
Corresponding author; e-mail: LieselotY.Hemeryck@UGent.be
Conflict of interest
Myriam Hesta has collaborated with several pet food companies and has received financial support from them. The remaining authors declare no conflicts of interest.
Funding
This work was supported by Research Foundation Flanders (FWO), grant number S001922N. Lieselot Y. Hemeryck is supported by FWO (1297623N).
References
Aguilar-Toalá, J.E., Cruz-Monterrosa, R.G. and Liceaga, A.M., 2022. Beyond human nutrition of edible insects: health benefits and safety aspects. Insects 13: 1007. https://doi.org/10.3390/INSECTS13111007
Anuduang, A., Mustapha, W.A.W., Lim, S.J., Jomduang, S., Phongthai, S., Ounjaijean, S. and Boonyapranai, K., 2024. Evaluation of Thai silkworm (Bombyx mori L.) hydrolysate powder for blood pressure reduction in hypertensive rats. Foods 13: 943. https://doi.org/10.3390/foods13060943
Areerat, S., Chundang, P., Lekcharoensuk, C. and Kovitvadhi, A., 2021. Possibility of using house cricket (Acheta domesticus) or mulberry silkworm (bombyx mori) pupae meal to replace poultry meal in canine diets based on health and nutrient digestibility. Animals 11: 2680. https://doi.org/10.3390/ANI11092680
Areerat, S., Chundang, P., Lekcharoensuk, C., Patumcharoenpol, P. and Kovitvadhi, A., 2023. Insect-based diets (house crickets and mulberry silkworm pupae): A comparison of their effects on canine gut microbiota. Veterinary World 16: 1627-1635. https://doi.org/10.14202/VETWORLD.2023.1627-1635
Belluco, S., Mantovani, A. and Ricci, A., 2018. Edible insects in a food safety perspective. In: Halloran, A., Flore, R., Vantomme, P. and Roos, N. (eds.) Edible insects in sustainable food systems. Springer, Cham, pp. 109-126. https://doi.org/10.1007/978-3-319-74011-9_7/TABLES/1
Biasato, I., Ferrocino, I., Colombino, E., Gai, F., Schiavone, A., Cocolin, L., Vincenti, V., Capucchio, M.T. and Gasco, L., 2020. Effects of dietary Hermetia illucens meal inclusion on cecal microbiota and small intestinal mucin dynamics and infiltration with immune cells of weaned piglets. Journal of Animal Science and Biotechnology 11: 64. https://doi.org/10.1186/S40104-020-00466-X
Boontiam, W., Hong, J., Kitipongpysan, S. and Wattanachai, S., 2022. Full-fat field cricket (Gryllus bimaculatus) as a substitute for fish meal and soybean meal for weaning piglets: effects on growth performance, intestinal health and redox status. Journal of Animal Science 100: skac080. https://doi.org/10.1093/JAS/SKAC080
Bosch, G., Loureiro, B.A., Schokker, D., Kar, S.K., Paul, A. and Sluczanowski, N., 2024. Black soldier fly larvae meal in an extruded food: effects on nutritional quality and health parameters in healthy adult cats. Journal of Insects as Food and Feed 10: 1595-1606. https://doi.org/10.1163/23524588-00001093
Broekman, H., Verhoeckx, K.C., Den Hartog Jager, C.F., Kruizinga, A.G., Pronk-Kleinjan, M., Remington, B.C., Houben, G.F., Knulst, A.C., de Jong, G. and Gaspari, M., 2016. Majority of shrimp-allergic patients are allergic to mealworm. Journal of Allergy and Clinical Immunology 137(4): 1261-1263. https://doi.org/10.1016/J.JACI.2016.01.005
Broekman, H.C.H.P., Knulst, A.C., de Jong, G., Gaspari, M., den Hartog Jager, C.F., Houben, G.F. and Verhoeckx, K.C.M., 2017. Is mealworm or shrimp allergy indicative for food allergy to insects? Molecular Nutrition and Food Research 61: 1601061. https://doi.org/10.1002/mnfr.201601061
BugGuide.Net, n.d. Cricket – Gryllodes sigillatus. Available at https://bugguide.net/node/view/723787 (accessed 11 February 2025).
BugGuide.Net, n.d. Gryllus assimilis. Available at https://bugguide.net/node/view/1576980/bgimage (accessed 11 February 2025).
Buy Silkworms for Reptiles: Nutritious Live Food, n.d. Available at https://www.reptilecentre.com/products/silkworms (accessed 11 February 2025).
Cao, Y., Araki, M., Nakagawa, Y., Deisen, L., Lundsgaard, A., Kanta, J.M., Hasegawa, Y., Aoki, N., Sato, M. and Watanabe, K., 2024. Dietary medium-chain fatty acids reduce hepatic fat accumulation via activation of a CREBH-FGF21 axis. Molecular Metabolism 87: 101991. https://doi.org/10.1016/J.MOLMET.2024.101991
Chia, S.Y., Tanga, C.M., Osuga, I.M., Alaru, A.O., Mwangi, D.M., Githinji, M., Ekesi, S., Fiaboe, K.K.M. and van Huis, A., 2019. Effect of dietary replacement of fishmeal by insect meal on growth performance, blood profiles and economics of growing pigs in Kenya. Animals 910: 705. https://doi.org/10.3390/ANI9100705
Cho, K.H., Sampath, V., Kim, A.J., Yoo, J.S. and Kim, I.H., 2023. Evaluation of full-fatted and hydrolysate mealworm (Tenebrio molitor) larvae as a substitute for spray-dried plasma protein diet in weaning pigs. Journal of Animal Physiology and Animal Nutrition 107: 589-597. https://doi.org/10.1111/JPN.13763
Cusack, D.F., Kazanski, C.E., Hedgpeth, A., Cordeiro, A.L., Karpman, J. and Ryals, R., 2021. Reducing climate impacts of beef production: a synthesis of life cycle assessments across management systems and global regions. Global Change Biology 27: 1721-1736. https://doi.org/10.1111/gcb.15509
Dai, C., Ma, H., Luo, L. and Yin, X., 2013. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. European Food Research and Technology 236: 681-689. https://doi.org/10.1007/S00217-013-1923-Z/FIGURES/6
Dai, Y., Huang, M., Xu, Y., Mu, L., Gao, J., Chen, H., Wu, Y. and Li, L., 2023. Enzymatic hydrolysis of silkworm pupa and its allergenicity evaluation by animal model with different immunization routes. Food Science and Human Wellness 12: 774-782. https://doi.org/10.1016/J.FSHW.2022.09.011
De Paepe, E., Wauters, J., Van Der Borght, M., Claes, J., Huysman, S., Croubels, S. and Vanhaecke, L., 2019. Ultra-high-performance liquid chromatography coupled to quadrupole orbitrap high-resolution mass spectrometry for multi-residue screening of pesticides, (veterinary) drugs and mycotoxins in edible insects. Food Chemistry 293: 187-196. https://doi.org/10.1016/J.FOODCHEM.2019.04.082
Fan, M., Choi, Y.J., Tang, Y., Kim, J.H., Kim, B.G., Lee, B., Lee, S. and Park, H., 2021. Agl9: A novel hepatoprotective peptide from the larvae of edible insects alleviates obesity-induced hepatic inflammation by regulating ampk/nrf2 signaling. Foods 10: 1973. https://doi.org/10.3390/FOODS10091973
Ferri, I., Canala, B. and Rossi, L., 2024. Unravelling the role of chitin and chitosan in prebiotic activity and correlation with cancer: a narrative review. Nutrition Reviews 83: e2015. https://doi.org/10.1093/NUTRIT/NUAE168
Gasco, L., Biancarosa, I. and Liland, N.S., 2020. From waste to feed: a review of recent knowledge on insects as producers of protein and fat for animal feeds. Current Opinion in Green and Sustainable Chemistry 23: 67-79. https://doi.org/10.1016/j.cogsc.2020.03.003
German Federal Institute for Risk Assessment (BfR), N. R. L. for A. protein in F. N., Garino, C., Zagon, J. and Braeuning, A., 2019. Insects in food and feed – allergenicity risk assessment and analytical detection. EFSA Journal 17(Suppl. 2). https://doi.org/10.2903/J.EFSA.2019.E170907
Gierisch, J.M., Beadles, C., Shapiro, A., McDuffie, J.R., Cunningham, N. and Bradford, D., 2014. Newcastle-Ottawa Scale coding manual for cohort studies. National Center for Biotechnology Information. Available at https://www.ncbi.nlm.nih.gov/books/NBK299087/ (accessed 5 February 2025).
Gil, G., Del Mónaco, S., Cerrutti, P. and Galvagno, M., 2004. Selective antimicrobial activity of chitosan on beer spoilage bacteria and brewing yeasts. Biotechnology Letters 26: 569-574. https://doi.org/10.1023/B:BILE.0000021957.37426.9B/METRICS
Godfray, H.C.J., Aveyard, P., Garnett, T., Hall, J.W., Key, T.J., Lorimer, J., Pierrehumbert, R.T., Scarborough, P., Springmann, M. and Jebb, S.A., 2018. Meat consumption, health and the environment. Science 361: eaam5324. https://doi.org/10.1126/SCIENCE.AAM5324/ASSET/BE346352-3C74-4B08-A6DA-FA0B1CC23D22/ASSETS/GRAPHIC/361_AAM5324_F5.JPEG
Håkenåsen, I.M., Grepperud, G.H., Hansen, J.Ø., Øverland, M., Ånestad, R.M. and Mydland, L.T., 2021. Full-fat insect meal in pelleted diets for weaned piglets: Effects on growth performance, nutrient digestibility, gastrointestinal function and microbiota. Animal Feed Science and Technology 281: 115086. https://doi.org/10.1016/J.ANIFEEDSCI.2021.115086
Hashim, S.A., Arteaga, A. and Van Itallie, T.B., 1960. Effect of a saturated medium-chain triglyceride on serum-lipids in man. The Lancet 275: 1105-1108. https://doi.org/10.1016/S0140-6736(60)90986-7
Hooijmans, C.R., Rovers, M.M., De Vries, R.B.M., Leenaars, M., Ritskes-Hoitinga, M. and Langendam, M.W., 2014. SYRCLE’s risk of bias tool for animal studies. BMC Medical Research Methodology 14: 43. https://doi.org/10.1186/1471-2288-14-43
Hu, Y., Du, Y., Yang, J., Kennedy, J.F., Wang, X. and Wang, L., 2007. Synthesis, characterization and antibacterial activity of guanidinylated chitosan. Carbohydrate Polymers 67: 66-72. https://doi.org/10.1016/J.CARBPOL.2006.04.015
Im, A.R., Yang, W.K., Park, Y.C., Kim, S.H. and Chae, S., 2018. Hepatoprotective effects of insect extracts in an animal model of nonalcoholic fatty liver disease. Nutrients 10: 735. https://doi.org/10.3390/NU10060735
Jafarzadeh, S., Qazanfarzadeh, Z., Majzoobi, M., Sheiband, S., Oladzadabbasabad, N., Esmaeili, Y. and Zare, M., 2024. Alternative proteins: a path to sustainable diets and environment. Current Research in Food Science 9: 100882. https://doi.org/10.1016/J.CRFS.2024.100882
Jantzen da Silva Lucas, A., Menegon de Oliveira, L., da Rocha, M. and Prentice, C., 2020. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chemistry 311: 126022. https://doi.org/10.1016/J.FOODCHEM.2019.126022
Jeong, M., Kim, Y.W., Min, J.R., Kwon, M., Han, B.S., Kim, J.G. and Jeong, S.H., 2013. Change in kidney damage biomarkers after 13weeks of exposing rats to the complex of Paecilomyces sinclairii and its host Bombyx mori larvae. Food and Chemical Toxicology 59: 177-186. https://doi.org/10.1016/j.fct.2013.05.041
Jian, S., Zhang, L., Ding, N., Yang, K., Xin, Z., Hu, M., Cheng, L. and Liu, X., 2022. Effects of black soldier fly larvae as protein or fat sources on apparent nutrient digestibility, fecal microbiota and metabolic profiles in beagle dogs. Frontiers in Microbiology 13: 1044986. https://doi.org/10.3389/FMICB.2022.1044986/BIBTEX
Jin, X., Yuan, B., Liu, M., Zhu, M., Zhang, X., Xie, G., Wang, F. and Li, Y., 2021. Dietary Hermetia illucens larvae replacement alleviates diarrhea and improves intestinal barrier function in weaned piglets challenged with enterotoxigenic Escherichia coli K88. Frontiers in Veterinary Science 8: 746224. https://doi.org/10.3389/FVETS.2021.746224/BIBTEX
Kang, Y., Applegate, C.C., He, F., Oba, P.M., Vieson, M.D., Sánchez-Sánchez, L. and Swanson, K.S., 2023. Yellow mealworm (Tenebrio molitor) and lesser mealworm (Alphitobius diaperinus) proteins slowed weight gain and improved metabolism of diet-induced obesity mice. Journal of Nutrition 153: 2237-2248. https://doi.org/10.1016/j.tjnut.2023.06.014
Kang, Y.K., Lee, B.Y., Bucci, L.R. and Stohs, S.J., 2018. Effect of a fibroin enzymatic hydrolysate on memory improvement: a placebo-controlled, double-blind study. Nutrients 10: 233. https://doi.org/10.3390/NU10020233
Kawasaki, K., Zhao, J., Takao, N., Sato, M., Ban, T., Tamamaki, K., Okuda, T. and Yamamoto, K., 2023. Sustenance trial to analyze the effects of black soldier fly larvae meal on the reproductive efficiency of sows and the hematological properties of suckling and weaning piglets. Animals 13: 3410. https://doi.org/10.3390/ANI13213410
Kim, M.W., Ham, Y.J., Kim, H.B., Lee, J.Y., Lim, J.D. and Lee, H.T., 2023. Anti-obesity effects of the larval powder of steamed and lyophilized mature silkworms in a newly designed adult mouse model. Foods 12: 3613. https://doi.org/10.3390/FOODS12193613
Kim, S.Y., Kwak, K.W., Park, E.S., Yoon, H.J., Kim, Y.S., Park, K. and Lee, J., 2020. Evaluation of subchronic oral dose toxicity of freeze-dried skimmed powder of Zophobas atratus larvae (frpfdZAL) in rats. Foods 9: 995. https://doi.org/10.3390/FOODS9080995
Kolakowski, B.M., Johaniuk, K., Zhang, H. and Yamamoto, E., 2021. Analysis of microbiological and chemical hazards in edible insects available to canadian consumers. Journal of Food Protection 84: 1575-1581. https://doi.org/10.4315/JFP-21-099
Kouřimská, L. and Adámková, A., 2016. Nutritional and sensory quality of edible insects. NFS Journal 4: 22-26. https://doi.org/10.1016/J.NFS.2016.07.001
Lähteenmäki-Uutela, A., Marimuthu, S.B. and Meijer, N., 2021. Regulations on insects as food and feed: a global comparison. Journal of Insects as Food and Feed 7: 849-856. https://doi.org/10.3920/JIFF2020.0066
Lanng, S.K., Zhang, Y., Christensen, K.R., Hansen, A.K., Nielsen, D.S., Kot, W. and Bertram, H.C., 2021. Partial substitution of meat with insect (Alphitobius diaperinus) in a carnivore diet changes the gut microbiome and metabolome of healthy rats. Foods 10: 1814. https://doi.org/10.3390/FOODS10081814
Lee, D.Y., Hong, K.S., Song, M.Y., Yun, S.M., Ji, S.D., Son, J.G. and Kim, E.H., 2020. Hepatoprotective effects of steamed and freeze-dried mature silkworm larval powder against ethanol-induced fatty liver disease in rats. Foods 9: 285. https://doi.org/10.3390/FOODS9030285
Li, G., Lee, P., Mori, N., Yamamoto, I., Kawasumi, K., Tanabe, H. and Arai, T., 2012. Supplementing five-point body condition score with body fat percentage increases the sensitivity for assessing overweight status of small to medium sized dogs. Veterinary Medicine: Research and Reports 3: 63-71. https://doi.org/10.2147/VMRR.S34665
Li, J., Wen, P., Qin, G., Zhang, J., Zhao, P. and Ye, Y., 2022. Toxicological evaluation of water-extract sericin from silkworm (Bombyx mori) in pregnant rats and their fetus during pregnancy. Frontiers in Pharmacology 13: 982841. https://doi.org/10.3389/FPHAR.2022.982841
Li, P. and Wu, G., 2023. Amino acid nutrition and metabolism in domestic cats and dogs. Journal of Animal Science and Biotechnology 14: 19. https://doi.org/10.1186/s40104-022-00827-8
Li, Y., Liu, M., Wei, Y., Li, L., Ma, D., Weng, Y. and Chen, X., 2024. Influence of a mixture of protein hydrolysate from black soldier fly larvae and schizochytrium on palatability, plasma biochemistry and antioxidative and anti-inflammatory capacity in cat diets. Animals 14: 751. https://doi.org/10.3390/ani14050751
Li, Y.X., Jin, X.B., Chu, F.J., Liu, M.Y., Shi, D.Y. and Zhu, J.Y., 2013. Scientific evaluation of the subchronic toxicity of Musca domestica larvae extracts in Sprague–Dawley rats. Food and Chemical Toxicology 59: 464-469. https://doi.org/10.1016/j.fct.2013.06.001
Liu, S., Wang, J., Li, L., Duan, Y., Zhang, X., Wang, T. and Chen, H., 2023. Endogenous chitinase might lead to differences in growth performance and intestinal health of piglets fed different levels of black soldier fly larva meal. Animal Nutrition 14: 411-424. https://doi.org/10.1016/J.ANINU.2023.05.008
Liu, Y., Zhang, Y., Zhang, X., Xu, Q., Yang, X. and Xue, C., 2017. Medium-chain fatty acids reduce serum cholesterol by regulating the metabolism of bile acid in C57BL/6J mice. Food and Function 8: 291-298. https://doi.org/10.1039/C6FO01207H
Liu, Z., Xia, L., Wu, Y., Xia, Q., Chen, J. and Roux, K.H., 2009. Identification and characterization of an arginine kinase as a major allergen from silkworm (Bombyx mori) larvae. International Archives of Allergy and Immunology 150: 8-14. https://doi.org/10.1159/000210375
Lu, S., Taethaisong, N., Meethip, W., Surakhunthod, J., Sinpru, B., Sroichak, T. and Sriket, P., 2022. Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: a review. Insects 13: 831. https://doi.org/10.3390/insects13090831
Machona, O., Mutanga, M., Chidzwondo, F. and Mangoyi, R., 2024. Sub-chronic toxicity determination of powdered Tenebrio molitor larvae as a novel food source. Toxicology Reports 12: 111-116. https://doi.org/10.1016/j.toxrep.2024.01.002
Malla, N., Roos, N., Van der Heide, M.E. and Nørgaard, J.V., 2024. Effect of feeding meal of yellow and lesser mealworm and defatted black soldier fly larvae on growth performance and gut health of weaned piglets. Animal Feed Science and Technology 309: 115917. https://doi.org/10.1016/j.anifeedsci.2024.115917
Marschall, M.J.M., Grundmann, S.M., Gessner, D.K., Wen, G., Most, E., Eder, K. and Ringseis, R., 2023. Feeding of Hermetia illucens larvae meal attenuates hepatic lipid synthesis and fatty liver development in obese Zucker rats. Nutrients 15: 287. https://doi.org/10.3390/nu15020287
Marschall, M.J.M., Grundmann, S.M., Seel, W., Simon, M.C., Schuchardt, S., Most, E., Eder, K. and Ringseis, R., 2024. Fat from Hermetia illucens alters the cecal gut microbiome and lowers hepatic triglyceride concentration in comparison to palm oil in obese Zucker rats. Journal of Nutrition 154: 455-468. https://doi.org/10.1016/j.tjnut.2023.09.019
Meyer, A.M., Meijer, N., van den Hil, E.F.H. and van der Fels-Klerx, H.J., 2021. Chemical food safety hazards of insects reared for food and feed. Journal of Insects as Food and Feed 7: 823-831. https://doi.org/10.3920/JIFF2020.0085
Meyer-Rochow, V.B., Gahukar, R.T., Ghosh, S. and Jung, C., 2021. Chemical composition, nutrient quality and acceptability of edible insects are affected by species, developmental stage, gender, diet and processing Method. Foods 10: 1036. https://doi.org/10.3390/FOODS10051036
Meyer, S., Gessner, D.K., Braune, M.S., Friedhoff, T., Most, E., Höring, M., Eder, K. and Ringseis, R., 2020a. Comprehensive evaluation of the metabolic effects of insect meal from Tenebrio molitor L. In growing pigs by transcriptomics, metabolomics and lipidomics. Journal of Animal Science and Biotechnology 11: 1-17. https://doi.org/10.1186/S40104-020-0425-7
Meyer, S., Schäfer, L., Röhrig, J., Maheshwari, G., Most, E., Zorn, H., Eder, K. and Ringseis, R., 2020b. Supplementation of sulfur-containing amino acids or essential amino acids does not reverse the hepatic lipid-lowering effect of a protein-rich insect meal in obese zucker rats. Nutrients 12(4): 1-15. https://doi.org/10.3390/NU12040987
Ng’ang’a, J., Fombong, F., Kiiru, S., Kipkoech, C. and Kinyuru, J., 2021. Food safety concerns in edible grasshoppers: a review of microbiological and heavy metal hazards. International Journal of Tropical Insect Science 41: 2103-2111. https://doi.org/10.1007/S42690-020-00372-9/TABLES/2
Nguyen, P., Kim, K.Y., Kim, A.Y., Kang, S.K., Osabutey, A.F., Jin, H. and Park, S., 2021. The additive memory and healthspan enhancement effects by the combined treatment of mature silkworm powders and Korean angelica extracts. Journal of Ethnopharmacology 281: 114520. https://doi.org/10.1016/J.JEP.2021.114520
Phaengphairee, P., Boontiam, W., Wealleans, A., Hong, J. and Kim, Y.Y., 2023. Dietary supplementation with full-fat Hermetia illucens larvae and multi-probiotics, as a substitute for antibiotics, improves the growth performance, gut health and antioxidative capacity of weaned pigs. BMC Veterinary Research 19: 1-11. https://doi.org/10.1186/S12917-022-03550-8
Poma, G., Cuykx, M., Amato, E., Calaprice, C., Focant, J.F. and Covaci, A., 2017. Evaluation of hazardous chemicals in edible insects and insect-based food intended for human consumption. Food and Chemical Toxicology 100: 70-79. https://doi.org/10.1016/J.FCT.2016.12.006
Premrov Bajuk, B., Zrimšek, P., Kotnik, T., Leonardi, A., Križaj, I. and Jakovac Strajn, B., 2021. Insect protein-based diet as potential risk of allergy in dogs. Animals 11: 1-15. https://doi.org/10.3390/ANI11071942
Ramos-Elorduy, J., 2009. Anthropo-entomophagy: Cultures, evolution and sustainability. Entomological Research 39: 271-288. https://doi.org/10.1111/J.1748-5967.2009.00238.X
Reed, M., Norwood, B.F., Hoback, W.W. and Riggs, A., 2021. A survey of willingness to consume insects and a measure of college student perceptions of insect consumption using Q methodology. Future Foods 4: 100046. https://doi.org/10.1016/J.FUFO.2021.100046
Regulation (EU) No 142/2011 of 25 February 2011 implementing Regulation (EC) No 1069/2009 on health rules concerning animal by-products not intended for human consumption. Official Journal of the European Union L 54, 26 February 2011.
Regulation (EU) No 1069/2009 of 21 October 2009 laying down health rules as regards animal by-products and derived products not intended for human consumption and repealing Regulation (EC) No 1774/2002. Official Journal of the European Union L 300, 14 November 2009.
Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Regulation (EU) No 142/2011 of the Commission as regards the provisions on processed animal protein. Official Journal of the European Union L 138, 25 May 2017, pp. 92-116.
Regulation (EU) 2021/1925 of the European Parliament and of the Council of 20 October 2021 on contestable and fair markets in the digital sector and amending Directives (EU) 2019/1937 and (EU) 2020/1828 (Digital Markets Act). Official Journal of the European Union L 313, 12 November 2021.
Reilly, L.M., Hu, Y., Von Schaumburg, P.C., De Oliveira, M.R.D., He, F., Rodriguez-Zas, S.L. and Swanson, K.S., 2022. Chemical composition of selected insect meals and their effect on apparent total tract digestibility, fecal metabolites and microbiota of adult cats fed insect-based retorted diets. Journal of Animal Science 100: 1-12. https://doi.org/10.1093/JAS/SKAC024
Scimago Journal and Country Rank, n.d. Website. Availalbe at https://www.scimagojr.com/ (accessed 6 January 2026).
Seo, K., Cho, H.W., Chun, J., Jeon, J., Kim, C., Kim, M. and Park, S., 2021. Evaluation of fermented oat and black soldier fly larva as food ingredients in senior dog diets. Animals 11: 1-16. https://doi.org/10.3390/ANI11123509
Son, S.J., Han, A.R., Sung, M.J., Hong, S.M. and Lee, S.H., 2023. Hermetia illucens fermented with Lactobacillus plantarum KCCM12757P alleviates dextran sodium sulfate-induced colitis in mice. Antioxidants 12: 1822. https://doi.org/10.3390/ANTIOX12101822
Spranghers, T., Michiels, J., Vrancx, J., Ovyn, A., Eeckhout, M., De Clercq, P. and De Smet, S., 2018. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Animal Feed Science and Technology 235: 33-42. https://doi.org/10.1016/J.ANIFEEDSCI.2017.08.012
Stella, R., Peggion, C., Bergantin, C., Biancotto, G., Frosini, M., Dreassi, E. and Damiani, E., 2022. Serum metabolomics and proteomics to study the antihypertensive effect of protein extracts from Tenebrio molitor. Nutrients 14: 3288. https://doi.org/10.3390/NU14163288
Stull, V.J., Finer, E., Bergmans, R.S., Febvre, H.P., Longhurst, C. and Manter, D.K., 2018. Impact of edible cricket consumption on gut microbiota in healthy adults, a double-blind, randomized crossover trial. Scientific Reports 8: 113. https://doi.org/10.1038/S41598-018-29032-2
Suchodolski, J.S., 2021. Analysis of the gut microbiome in dogs and cats. Veterinary Clinical Pathology 50: 6. https://doi.org/10.1111/VCP.13031
Tabata, E., Kashimura, A., Kikuchi, A., Masuda, H., Miyahara, R., Hiruma, Y. and Okamoto, T., 2018. Chitin digestibility is dependent on feeding behaviors, which determine acidic chitinase mRNA levels in mammalian and poultry stomachs. Scientific Reports 8: 1461. https://doi.org/10.1038/s41598-018-19940-8
Thompson, A., 2008. Ingredients: where pet food starts. Topics in Companion Animal Medicine 23: 127-132. https://doi.org/10.1053/j.tcam.2008.04.004
van Heugten, E., Martinez, G., McComb, A. and Koutsos, L., 2022. Improvements in performance of nursery pigs provided with supplemental oil derived from black soldier fly (Hermetia illucens) larvae. Animals 12: 3210. https://doi.org/10.3390/ANI12233251
Verhoeckx, K.C.M., van Broekhoven, S., den Hartog-Jager, C.F., Gaspari, M., de Jong, G.A.H. and Wichers, H.J., 2014. House dust mite (Der p 10) and crustacean allergic patients may react to food containing yellow mealworm proteins. Food and Chemical Toxicology 65: 364-373. https://doi.org/10.1016/J.FCT.2013.12.049
Wood, J.D. and Migicovsky, B.B., 2011. The effect of dietary oils and fatty acids on cholesterol metabolism in the rat. Canadian Journal of Animal Science 36: 433-438. https://doi.org/10.1139/O58-049
Xia, Z., Chen, J. and Wu, S., 2013. Hypolipidemic activity of the chitooligosaccharides from Clanis bilineata (Lepidoptera), an edible insect. International Journal of Biological Macromolecules 59: 96-98. https://doi.org/10.1016/J.IJBIOMAC.2013.04.017
Yoon, Y.I., Chung, M.Y., Hwang, J.S., Han, M.S., Goo, T.W. and Yun, E.Y., 2015. Allomyrina dichotoma (Arthropoda: Insecta) larvae confer resistance to obesity in mice fed a high-fat diet. Nutrients 7: 1978-1991. https://doi.org/10.3390/NU7031978
Yu, M., Li, Z., Chen, W., Rong, T., Wang, G. and Ma, X., 2019. Hermetia illucens larvae as a potential dietary protein source altered the microbiota and modulated mucosal immune status in the colon of finishing pigs. Journal of Animal Science and Biotechnology 10: 1-11. https://doi.org/10.1186/S40104-019-0358-1
Yu, M., Li, Z., Chen, W., Wang, G., Rong, T., Liu, Z. and Ma, X., 2020. Hermetia illucens larvae as a Fishmeal replacement alters intestinal specific bacterial populations and immune homeostasis in weanling piglets. Journal of Animal Science 98(3): skaa112. https://doi.org/10.1093/JAS/SKZ395
Yun, S.M., Cho, J.M., Hong, K.S., Lee, D.Y., Ji, S.D., Son, J.G. and Kim, E.H., 2017. Gastroprotective effect of mature silkworm, Bombyx mori, against ethanol-induced gastric mucosal injuries in rats. Journal of Functional Foods 39: 279-286. https://doi.org/10.1016/J.JFF.2017.10.036
Zacharis, C., Bonos, E., Giannenas, I., Skoufos, I., Tzora, A., Voidarou, C. and Dimitriadis, G., 2023. Utilization of Tenebrio molitor larvae reared with different substrates as feed ingredients in growing pigs. Veterinary Sciences 10: 393. https://doi.org/10.3390/VETSCI10060393
Zielińska, E., Karaś, M. and Jakubczyk, A., 2017. Antioxidant activity of predigested protein obtained from a range of farmed edible insects. International Journal of Food Science and Technology 52: 306-312. https://doi.org/10.1111/IJFS.13282







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}