Abstract
This study evaluated the impact of feeding Lactiplantibacillus plantarum RS5 probiotic and its products on gut health and immune status in laying hens under heat stress. A total of 192 Isa White pullets were randomly assigned to individual cages in four environmentally controlled chambers. During the adaptation period from 1 to 21 d, all birds received the same basal diet. On d 22, the birds were divided into six treatment groups, with half in two rooms subjected to cyclic daily heat stress reaching about 30 °C and the other half allocated to two houses under regular temperature conditions. The layers were fed one of three diets: (1) Control, (2) Control + L. plantarum RS5 probiotic, or (3) Control + L. plantarum RS5 postbiotic. The feeding trial lasted for five months. The study found that heat stress negatively impacted the birds’ blood profile, with significant decreases in red blood cells, haemoglobin, haematocrit, mean corpuscular volume, platelets, mean platelet volume and plateletcrit compared to the control groups at two and five months of the trial (P<0.05). The intestinal lesions caused by heat stress showed numeric improvements with probiotic/postbiotic supplementation, including mucosal cell degeneration and the number of villi (P>0.05). Dietary probiotics numerically increased the humoral immune response to Newcastle Disease vaccine at 2 and 4 weeks post-vaccination (P>0.05), compared to the control group. In conclusion, the study showed that dietary L. plantarum probiotics and postbiotics may alleviate the impact of heat stress in the poultry industry. The findings highlighted the potential for evaluating various dietary probiotics and postbiotics in new heat stress models for layers.
1 Introduction
The primary objective of the global poultry industry is to ensure a consistent supply of eggs and meat to consumers. However, the increasing environmental temperature worldwide poses a significant threat to the poultry industry in many countries (Ahmad et al., 2022). When birds experience heat stress, they employ various strategies, including behavioural, hormonal, physiological and biochemical adjustments, to maintain homeostasis (Bueno et al., 2017). The intestinal tract, being highly responsive, undergoes changes during heat stress. The effective functioning of the intestinal tract is crucial for poultry production, impacting overall health and performance of the birds (Rostagno, 2020).
Numerous studies have explored how heat stress affects the immune responses of chickens, yielding variable results (Mashaly et al., 2004). Heat stress can reduce antibody production in young chickens (Zulkifli et al., 2000). To combat such negative impacts on poultry gut health and immunity, incorporating feed additives such as antibiotics into the diet at sub-therapeutic levels has become a common practice. These antibiotics alter the intestinal flora in chickens and enhance immune response, improving their ability to control diseases (Danladi et al., 2022). When used as growth promoters in layer feeds, antibiotics can alleviate some effects of heat stress and enhance overall performance (Loh et al., 2014). However, the unregulated use of antibiotics has led to the development of antibiotic-resistant bacteria and can increase residues in animal products, posing significant risks to both animal and consumer health (Danladi et al., 2022). Thus, there is a compelling need for safer alternatives that can provide similar benefits to animal production, with probiotics and post-biotic metabolites emerging as promising substitutes.
Probiotics are live microorganisms that colonise the gastrointestinal tract, enhancing the natural microbial environment and inhibiting harmful pathogens (Loh, 2017). They exert positive effects through various mechanisms, including immunomodulation, the inhibition of bacterial toxins and competition with pathogens for adhesion to the intestinal epithelium and nutrients. Probiotics can promote gut maturation and integrity, regulate the immune system, prevent inflammation and improve growth and metabolism (Hu, 2024; Naeem and Bourassa, 2025). Although recent developments have produced stable probiotics (Terpou et al., 2019), the use of certain types carries concerns, particularly their potential to harbour antibiotic-resistant genes, which could transfer between organisms and diminish long-term usage (Imperial and Ibana, 2016; Liu et al., 2024).
In contrast, post-biotics consist of non-living microbial metabolites and components derived from probiotics. Because they do not contain live microorganisms, postbiotics eliminate the risks associated with potential infection, such as bacteraemia or fungemia, making them safer for use in animal production (Yelin et al., 2019). Postbiotics boast several advantages, such as diverse molecular structures, extended shelf life and well-defined safety profiles (Shigwedha et al., 2014). Post-biotics have demonstrated significant antioxidant properties, attributed to high levels of various antioxidant enzymes such as superoxide dismutase and peroxidases. These metabolites enhance the health of intestinal villi, boost lactic acid production and lower the levels of Enterobacteria spp.. These effects contribute to improved immune responses and overall gut health, as well as enhanced growth performance (Saeed et al., 2023).
While both probiotics and postbiotics promote gut health, enhance nutrient utilisation and support animal growth, postbiotics do this without the need for live cells. This results in a more stable and safe option for farmers and animal producers. Research has indicated that post-biotics produced from various Lactiplantibacillus plantarum cultivars display broad antagonistic actions against various pathogens and have shown effectiveness in improving growth metrics, meat quality and the ability to withstand heat stress in broilers, laying hens and pigs (Shenderov, 2013; Danladi et al., 2022).
Furthermore, substances like lactic acid and bacteriocins from lactic acid bacteria, found in postbiotics, exhibit antimicrobial properties and are increasingly being viewed as effective alternatives to traditional antibiotics (Zhong et al., 2022). Studies have shown that post-biotics derived from L. plantarum can inhibit a range of pathogenic bacteria, including Listeria monocytogenes, Salmonella typhimurium, Escherichia coli and vancomycin-resistant Enterococci spp. (Kareem et al., 2014). Notably, post-biotics have strong efficacy, especially under heat stress conditions, which suggests that they can provide benefits similar to those offered by probiotics, without the associated risks (Loh et al., 2014). Overall, the exploration of post-biotics illustrates a step forward in ensuring the health of poultry while addressing the problems relating to antibiotic use in animal production.
This study assessed the impact of dietary L. plantarum RS5 probiotic and post-biotic preparations on gut health and immunity parameters of laying hens, particularly under heat stress conditions.
2 Materials and methods
Ethical statement
All experimental procedures used in this study were approved by the Institutional Animal Care and Use Committee (IACUC) at the American University of Beirut (Approval letter # 20-10-583) and adhered to ARRIVE guidelines (Percie du Sert et al., 2020). This work complemented another study that has already been published (Farran et al., 2024). Consequently, both publications have the same approval letter number from IACUC.
Lactiplantibacillus plantarum strain
The L. plantarum strain RS5 (NCIMB 701088) was obtained from the NCIMB laboratory in the United Kingdom and was originally isolated from cheese. To cultivate the bacteria, it was suspended in a nutrient-rich De Man, Rogosa and Sharpe (MRS) broth and then kept at a temperature of 37 °C for 48 h. To confirm the presence of the bacteria, Gram staining was applied. This revealed the presence of Gram-positive, non-spore-forming rod-shaped cells when examined under a microscope. The bacterial suspension was then transferred to MRS agar growth medium and allowed to grow for 48 h at the same temperature (37 °C).
From the resulting colonies, white, circular forms were randomly selected. Some of these were further isolated by streaking on MRS agar to ensure pure cultures, while others were subjected to Gram staining for confirmation. Subsequently, these colonies were suspended in a sterile 0.85% saline solution. The optical density of this bacterial suspension was adjusted to 3% at a wavelength of 450 nm. After a series of dilutions, culturing on MRS agar and counting the colonies, it was determined that this initial solution contained 1015 colony-forming units (cfu/ml). The original bacterial cultures were preserved for future use at a temperature of -80 °C in MRS broth.
Preparation of post-biotics from Lactiplantibacillus plantarum strains
Cultures of L. plantarum were prepared by introducing 10% (by volume) of active bacterial cells with a concentration of 109 cfu/ml into MRS media. These cultures were incubated at a temperature of 30 °C for 10 h. Following the incubation, the mixture underwent centrifugation at 10,000×g and 4 °C for 15 min using an Eppendorf 5810 centrifuge (Eppendorf, Hamburg, Germany).
The resulting cell-free supernatant (CFS) was collected by filtering it through a cellulose acetate membrane with a pore size of 0.22 microns, as described by Loh et al. (2014). The CFS was preserved at a temperature of -20 °C until it was needed for the feeding trial. During the trial, the liquid post-biotics were combined with the animal feed using a horizontal feed mixer and a three-way mixing technique, whereby 300 ml of the CFS solution in MRS broth was added per 100 kg of feed.
Preparation of probiotics from Lactiplantibacillus plantarum strains
Bacterial growth was promoted using MRS agar. The overnight culture of the isolates was introduced and allowed to incubate for a period ranging from 24 to 48 h. Following this incubation, bacterial colonies were collected and re-suspended in a solution of phosphate-buffered saline (PBS) at pH 7.4. The bacterial count was fine-tuned to a concentration of 3.19 cfu/ml with spectrophotometry.
Subsequently, the bacterial suspension was blended with the basal diet at a concentration of 200 ml of the RS5 solution in MRS broth for every 100 kg of feed. This mixing process was achieved by employing a three-way mixing technique within a horizontal mixer. Blending the bacterial suspension with the basal diet was performed once every week to avoid any significant loss of L. plantarum viability upon storage or feeding under heat stress conditions (Trisnawita et al., 2018).
Bird housing and treatments
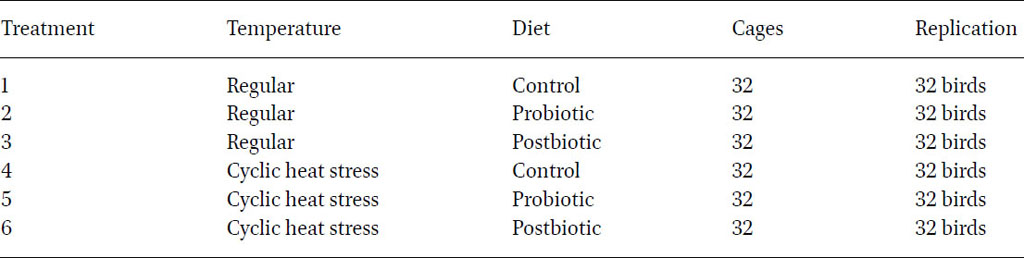
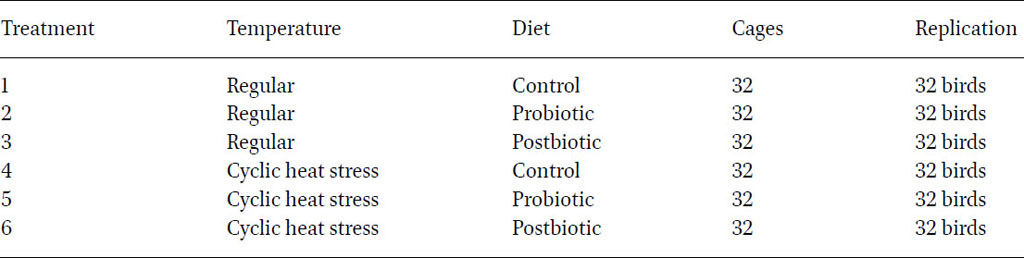
The study took place at the American University of Beirut’s research facilities (AREC) in the Beqaa region, utilising four identical environmentally controlled poultry houses. The experiment ran over six months, comprising a 21 d adaptation period followed by five months of the experimental phase. The initial live body weight was recorded for all birds at the end of the adaptation phase to allocate birds into different treatment homogeneously. Specifically, 192 twenty-week-old pullets of the Isa white strain were evenly distributed into six groups of 32 birds, each housed individually. Within each group, birds were further divided between two areas, accommodating 16 birds in each. The first two areas maintained regular temperature conditions, varying from 20 °C during the night to 26 °C during the day. The other two areas were exposed the birds to cyclic heat stress, reaching approximately 30 °C for 4 h consecutively on a daily basis (Humam et al., 2019; Farran et al., 2024). Temperature levels were monitored daily at 10:00 am, 1:00 pm and 4:00 pm, as well as weekly at 4:00 am. Additionally, birds in each house were allocated into one of three diet groups; a control basal diet, control with probiotic supplementation and control with postbiotic supplementation. Throughout the experiment, birds had free access to water and feed, provided in accordance with the recommendations from a husbandry manual (Institut de Sélection Animale BV, Boxmeer, the Netherlands). The experimental design, detailed in Table 1, encompassed six distinct treatments for the birds.

Control and experimental diets and heat exposure for laying hens.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
Macroscopic lesions and pathology scoring of the intestine
Four birds from each treatment were sacrificed at two and five months after heat stress initiation to evaluate macroscopic lesions and pathology scores of various parts of the intestines. Lesions included the presence of the following: thin/ thick intestinal wall, intestinal haemorrhage, watery intestinal content and undigested feed. Intestinal scores were recorded from 0 to 3 whereas a score of 3 indicated pronounced macroscopic lesions and a score of 0 had no lesions.
Histopathology
From the mid-portion of the jejunum, 5 cm was excised from four sacrificed birds per treatment at the end of the experiment to evaluate intestinal health using haemoxylin and eosin (H&E) staining (Shaib et al., 2004). Phosphate-buffered saline was used to wash the samples before storing the tissues in 10% formalin. Dehydration of the sample, embedding and cutting three 5 µm-thick sections was carried out to position them on glass slides, which were then heated until dry. The sections were stained using H&E and examined under the microscope to determine the number of villi and mucosal cells in the intestinal lining. The percentage of fields showing altered morphology of microvilli and mucosal cells degeneration was recorded.
Blood collection and seroconversion studies
Studying complete blood count (CBC) at various months after the experiment began was needed for monitoring health status, evaluating nutritional needs, assessing immune function and understanding environmental stress impacts. Therefore, blood samples were collected from eight laying hens from each treatment at two, four and five months post initiation of heat stress to test CBC. The haematological tests measured white blood cells (WBC, 109/l), red blood cells (RBC, 106/mm3), haemoglobin (HGB, mg/dl), haematocrit (HCT%), mean corpuscular volume (MCV, fl), mean corpuscular haemoglobin (MCH, pg), mean corpuscular haemoglobin concentration (MCHC, %), red cell distribution width (RDW, %), platelets (PLT, 103/ml), mean platelet volume (MPV, fl), plateletcrit (PCT, ng/ml) and platelet distribution width (PDW%).
On the other hand, blood samples were collected from eight laying hens from each treatment two and four weeks post vaccination with live NDV to evaluate humoral immunity. It was important to collect serum samples at this specific timeframe, as it was expected to show a significant immune response resulting from vaccination. Blood was centrifuged at 2,000 rpm for 10 mins and sera were collected in 1.5 ml microcentrifuge tubes, followed by storage at -80 °C for subsequent assessments. These included the determination of NDV hemagglutination inhibition (HI) titres and subsequent western immunoblotting to assess immunity at protective levels against the fusion protein.
Haemagglutination-inhibition test
The HI test for antibody titres against Newcastle disease virus (NDV) was performed using the OIE/FAO 2020 protocol (OIE/FAO, 2020). In brief, sera were serially diluted half in V-shaped wells and each well of the HI plates received 4 HAU of NDV virus or antigen. The plate was incubated at room temperature for 45 min and was later supplemented with 0.050 ml of 0.5% chicken red blood cells and incubated for an additional 30 min. Positive and negative sera samples were used.
SDS PAGE western immunoblotting:
The SDS-PAGE was followed by western immunoblotting and were applied on the sera from all birds to quantify the mean specific immune responses in the sera. This was collected onto trans-blotted fusion protein carried from the SDS resolving gel to the cellulose membrane of the western blotting. An amount of 100
Statistical design and analysis:
The experiment was conducted as a randomised block design with a 2 × 3 factorial design, resulting in a total of six treatments, with each treatment having 32 birds per replicate. Data analysis was carried out using multivariate methods, two-way ANOVA and mean comparisons were conducted at a 95% confidence level, where significance was attained at P≤0.05. Frequencies of intestinal gross lesions were statistically compared using one-way ANOVA at 95% confidence level, where significance was attained at P≤0.05. Statistical analysis was performed using SPSS software (version 25, IBM, Armonk, NY, USA).
3 Results and discussion
Complete blood count
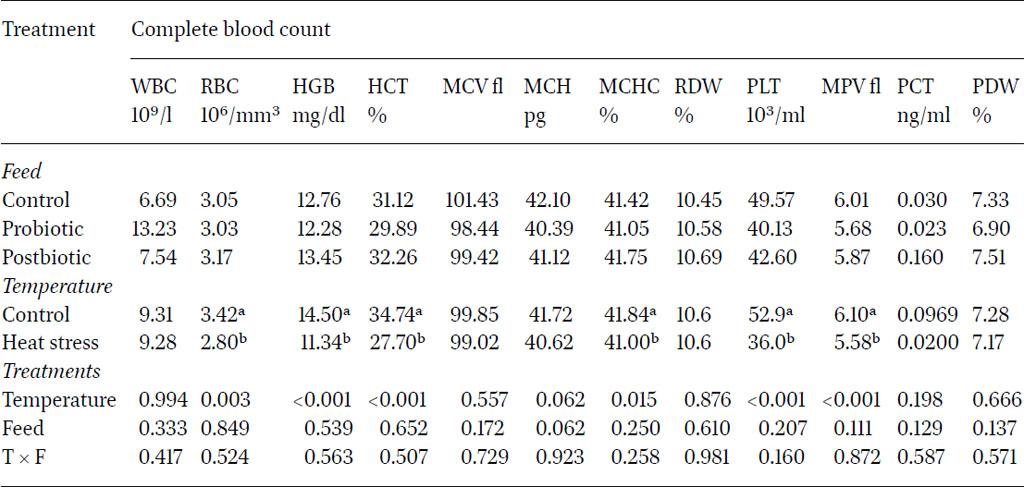
Levels of the CBC parameters are shown in Tables 2, 3 and 4. The normal ranges of the haematological parameters in chickens are RBC 2.5–3.5 ×106/µl, Hb 7–13 g/dl and WBC 12–30 × 103/µl. The MCV is used to calculate the average erythrocyte size, MCH haemoglobin amount per blood cell and the MCHC haemoglobin relative to the size of the cell per red blood cell. These have normal ranges are MCV 90–140 fl, MCH 33–47 pg/cell and MCHC 26–35 g/dl (Jain, 1993; Bounous and Stedman, 2000; Osadcha, 2023).
Mean blood count of layers for white blood cells, red blood cells, haemoglobin, haematocrit, mean corpuscular volume, mean corpuscular haemoglobin, mean corpuscular haemoglobin concentration, red cell distribution width, platelets, mean platelet volume, plateletcrit and platelet distribution width under different feed and temperature parameters at two months after heat stress initiation.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
Mean blood count of layers for red blood cells, haemoglobin, haematocrit, mean corpuscular volume, mean corpuscular haemoglobin, mean corpuscular haemoglobin concentration, red cell distribution width, platelets, mean platelet volume, plateletcrit and platelet distribution width in layers under different feed and temperature parameters at four months after heat stress initiation.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
Mean blood count of layers for white blood cells, red blood cells, haemoglobin, haematocrit, mean corpuscular volume, mean corpuscular haemoglobin, mean corpuscular haemoglobin concentration, red cell distribution width, platelets, mean platelet volume, plateletcrit and platelet distribution width in layers under different feed and temperature parameters at five months after heat stress initiation.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
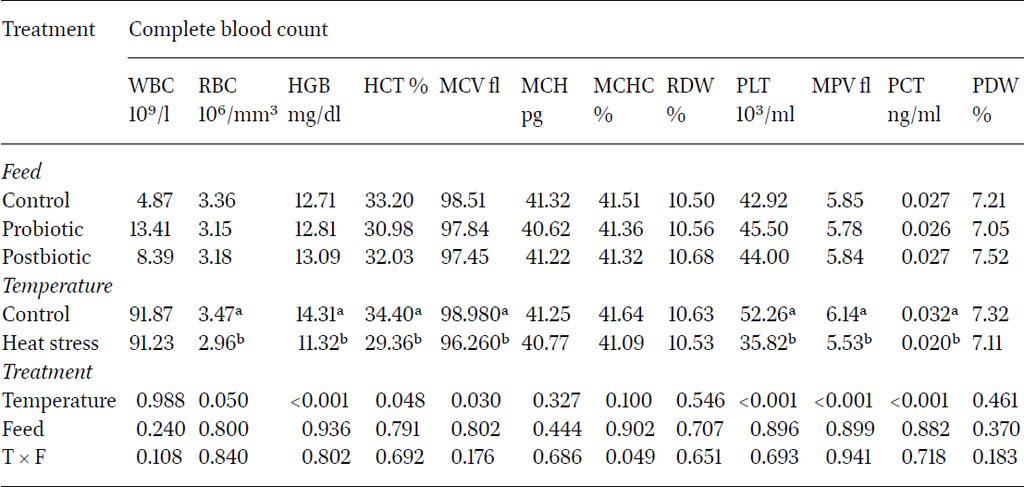
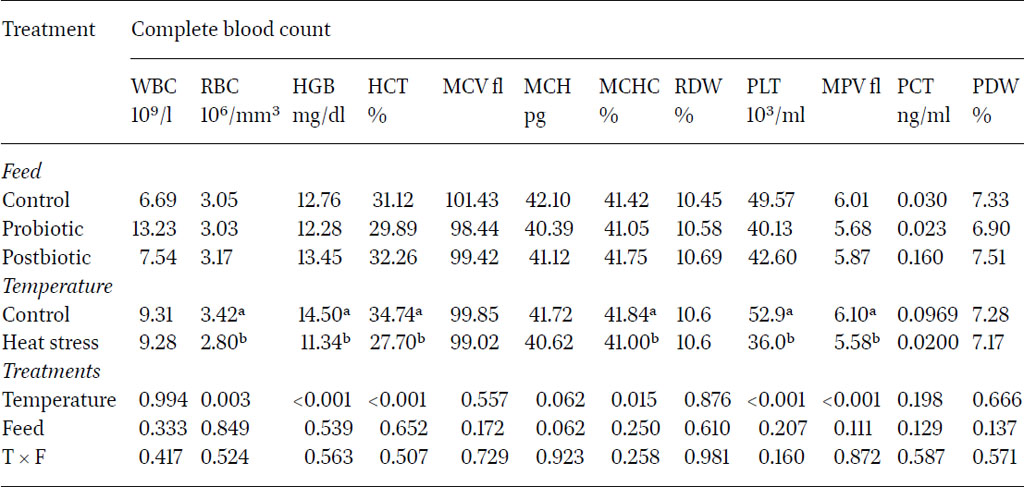
Results showed that exposure to heat significantly decreased the concentration of RBC, HGB, HCT, MCV, PLT, MPV and PCT compared to control groups at two and five months after the initiation of heat stress. Remarkably, heat stress significantly increased platelet distribution width (PDW) four months after the initiation of heat stress.
The decrease in RBC, ranging between 3.81×106/µl and 2.8×106/µl has been reported in another study which showed an indirect relationship between heat stress and the decrease in RBCs in heat-stressed quail (Mahmoud et al., 2013). The recorded HCT values, ranging between 27.32% and 36.11%, varied with ambient rearing temperature. According to Kubena et al. (1972) and Vo et al. (1978), the exposure of chickens to high temperatures causes a decrease in blood haematocrit values. Due to the positive relation between RBC number and HGB concentrations in blood, HGB showed a significant decline (14.5 g/dl vs 11.34 g/dl for control and heat stress birds, respectively).
Haemoglobin is only the protein constituent of red blood cells where it serves as a carrier of oxygen for use in various processes in cells. The current findings agreed with Dinu et al. (2004), who reported that reduction in the level of HGB and HCT values were the consequence of the heat stress. Thermal stress reduced the HCT, which may have been due to decreased production of erythrocytes or both the decreased erythrocyte number and size (Altan et al., 2000). The MCV indicates the size of the red blood cells, which was negatively affected with heat stress exposure. Other research showed that the concentrations of blood HCT, RBC, MCV, MCH, MCHC, RDW, HGB, PLT, PCT and MPV were lower in the heat stress groups compared with the control group in ducks (Byung-Sung et al., 2018). In the current study, the same pattern was further reflected in the MCHC values at two months whereby the results indicated that, while the type of feed alone did not significantly influence blood count, the interaction between feed type and temperature conditions had a significant impact (P<0.05). Feeding probiotics and post-biotics decreased blood count under heat stress conditions.
The platelets profile was assessed as platelet count PLT, mean platelet volume MPV, plateletcrit PCT and platelet distribution width PDW. The results clearly showed that heat stress significantly reduced platelet parameters all over the experimental period except for PDW. Previous research supported these results, whereby heat-stressed ducks and other birds showed lower RBC and PLT and lower blood gas concentration, except PO2 (Park et al., 2015). Although the platelet parameters were reduced due to heat stress, PDW showed an opposite trend after four months of heat stress exposure. The PDW represents the heterogenicity in platelet morphology due to the presence of large along with normal sized platelets. However the literature is not consistently reporting similar results, whereby Abudabos et al. (2018) and Köseman et al. (2021) reported a PDW reduction under heat stress in layers and broilers.
Platelets are involved in clotting. When heat stress conditions lead to a decrease in platelets, birds are more prone to bruising, which affects the healing process. This is highly correlated to enteric lesions, due to increased risk of haemorrhage (Bueno et al., 2017). A previous study confirmed a reduction in the number of platelets in the blood of broiler chickens in response to heat stress (Saif and Fadly, 2008).
Microscopic intestinal morphology
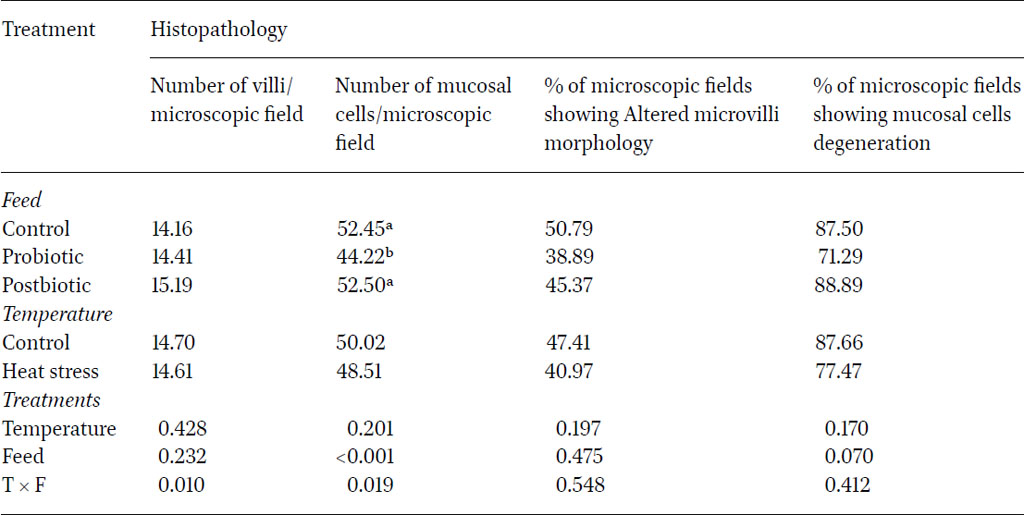
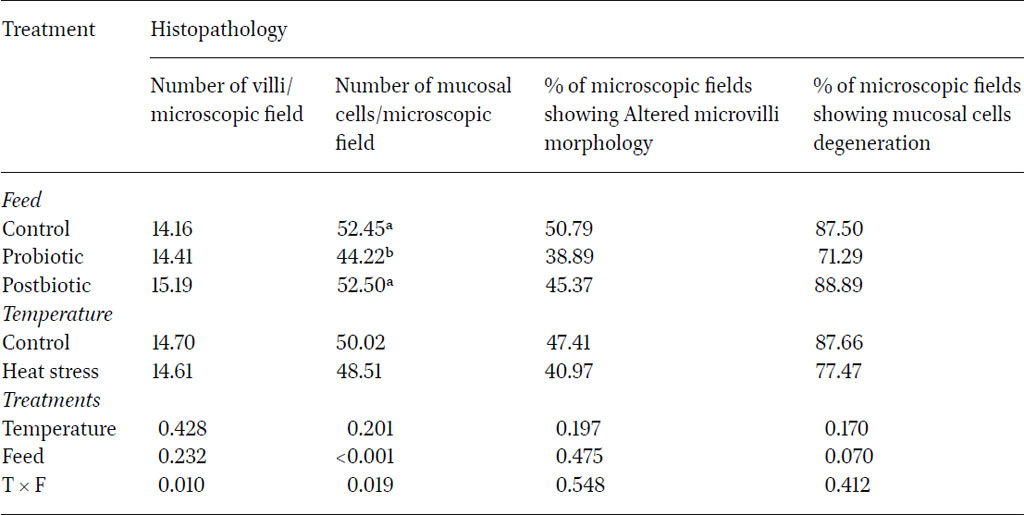
The number of villi and mucosal cells are important indicators of gut function and animal health. The villi are the key components responsible for the absorbance of nutrients in the small intestine. Increasing the villi number may result in higher nutrient absorption thereby improving growth performance. The results (Table 5) showed numerically high number of villi in the microscopic field in intestinal lining of birds fed diet containing post-biotic supplementation, with 15.19 compared to 14.16 in the control group. Heat stress slightly affected the density of the villi number, which had lower value compared to sections from the control group (P>0.05). The significant impact of the interaction between the two factors (P<0.05) suggested that the presence of probiotics under optimal temperature conditions may play a crucial role in promoting intestinal health by increasing the surface area for nutrient absorption, facilitated by the proliferation of intestinal villi.
Number of villi, mucosal cells and altered microvilli morphology areas under microscopic fields and percentage of microscopic fields showing no mucosal cells degeneration under different temperatures and feed parameters in layers during five months of the experiment
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
Microscopic fields showing altered morphology of microvilli in the intestines were insignificantly the highest in birds fed the control diet (50.79%) regardless of heat stress conditions (P>0.05). Probiotic and post-biotic treatments led to a numerical reduction in the areas showing microvilli alteration whereby only 38.9% and 45.4% of the microscopic fields observed during this study showed areas of altered microvilli morphology. These findings indicated that these feed additives can enhance the structural integrity of the mucosal layer in the jejunum and may potentially mitigate the effects of microvilli morphology alterations. Therefore, these additives could help maintain microvilli function that is essential for nutrient absorption (Abd El-Ghany et al., 2022).
The enhancement in morphological aspects of the intestinal lining contributes to increased absorption, likely arising from the competitive interactions between lactic acid bacteria (LAB) and pathogenic microorganisms, influencing resource utilisation and ecological niches (Shojadoost et al., 2022). It is worth noting that although the probiotic treatment significantly reduced the number of mucosal cells, it limited the percentage of degenerating mucosal cells (71.29 per microscopic field) in comparison to the control (87.5) and postbiotic group (88.89), although this was not significant (P>0.05). These results emphasised the positive role of probiotics in supporting the mucosal layer integrity and function and are in agreement with other studies (Zhang et al., 2016; Gadde et al., 2017; Wang et al., 2018). Zhang et al. (2016) demonstrated that specific probiotic strains improved intestinal morphology, increased villus height and enhanced the overall gut barrier function. Similarly, Wang et al. (2018) reported that probiotics promoted beneficial gut microbiota and stimulated the immune response in poultry, leading to improved mucosal integrity and reduced incidence of gastrointestinal diseases. Together, these studies highlight the potential of probiotics as a strategic intervention to bolster mucosal health and improve the overall well-being of poultry. Overall, the adopted model in the current study was not stressful enough to induce significant changes in microscopic lesions (Nanto-Hara et al., 2020; Ahmad et al., 2022). This was reflected through the lack of significance between heat stressed and control birds regarding the parameters.
Gross lesions
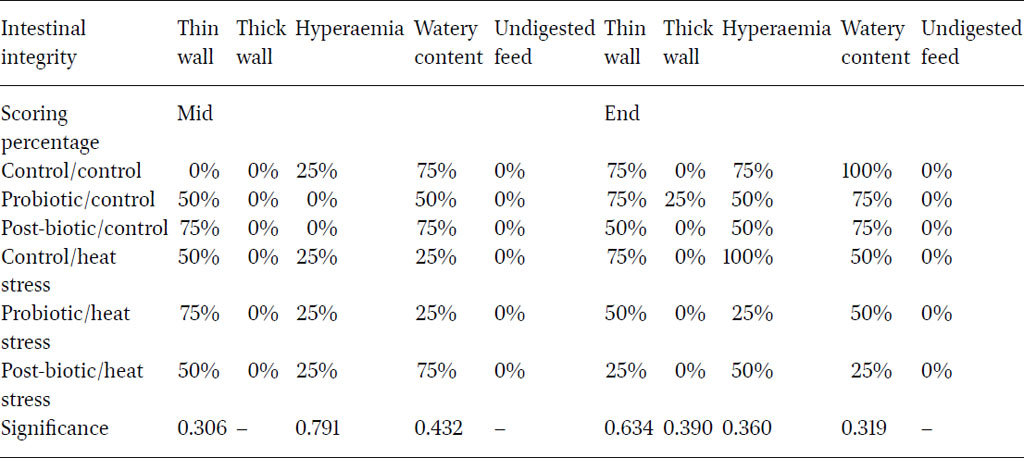
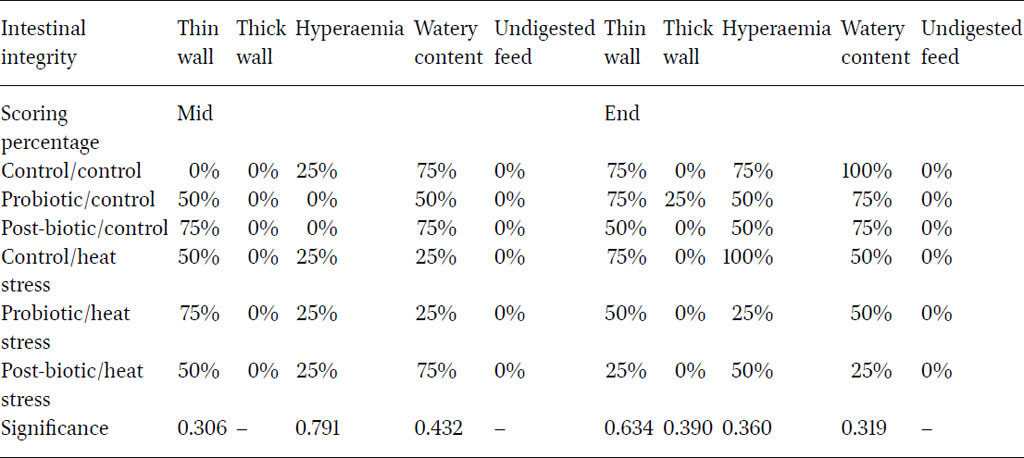
The least observed macroscopic lesions were the presence of thick intestinal wall and undigested feed in the intestinal lumen (Table 6). Post-biotics exerted a positive impact in reducing major macroscopic lesions such as thinning of the intestinal wall (intestinal tissue integrity), hyperaemia (inflammation) and watery digesta (against pathogens), specifically towards the end of the experiment. These results were in agreement with other literature emphasising the role of post-biotics and probiotics in relieving heat stress impact. Abd El-Ghany et al. (2022) illustrated the efficacy probiotics, postbiotics and antibiotics on the enteric lesions where the histopathological effects were significantly improved for the treated groups.
Percentages of intestinal thin wall, thick wall, haemorrhage, watery content and undigested feed in layers under six different treatments taken at the mid and the end of the five month experimental trial.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
The intestinal lesion evaluation reflected a mild impact of heat stress on intestinal health during the first half of the experiment. Although there were no significant differences in the second half of the experiment, there was a trend for heat stress to increase score for thin walls (75%), hyperaemia (100%) and watery digesta (50%). Post-biotics reduced the frequency of birds with watery digesta to 25%; while probiotics lessened hyperaemia frequency to 25%. A recently published study (Xu et al., 2020) observed that L. plantarum supplementation in layer diets enhanced mucosal integrity and reduced intestinal inflammation.
Adding to this, the duodenum, jejunum and ileum intestinal sections were scored for lesions in support for the above results (Table 7). The jejunum was affected, mostly scoring 1 in the control group, whereas feeding the probiotic enhanced integrity, reducing the score to 0.375. For the ileum, feeding post-biotics showed a positive impact, scoring 0.125 in comparison to the control (0.75). Nevertheless, heat stress affected all the intestinal parts negatively showing lower scoring compared to the control. This emphasised the negative role of heat stress on the integrity of the mucosal tissue of the intestine reflected in the reduction of mucosal cells and the positive impact of pro- and post-biotics on decreasing morphology alterations.
Average scoring of the duodenum, jejunum and ileum parts of the intestine under different feed and temperature parameters in layers at the mid and end of the five month experimental period.
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
Previous studies have reported that alterations in the intestinal epithelium integrity and microbiota, which could disturb the homeostasis of the intestinal ecosystem and lead to enteritis, have been linked to heat stressed birds (Burkholder et al., 2008; Awad et al., 2018).
Inflammation of the intestinal mucosa can directly affect gut barrier function, impairing the absorption of nutrients and slowing down the growth of birds. It has been indicated that exposure to acute heat stress causes a substantial impairment in the gut microbiota, intestinal integrity and villus morphology (Burkholder et al., 2008).
Haemagglutination inhibition titres against Newcastle Disease virus
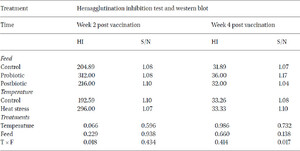
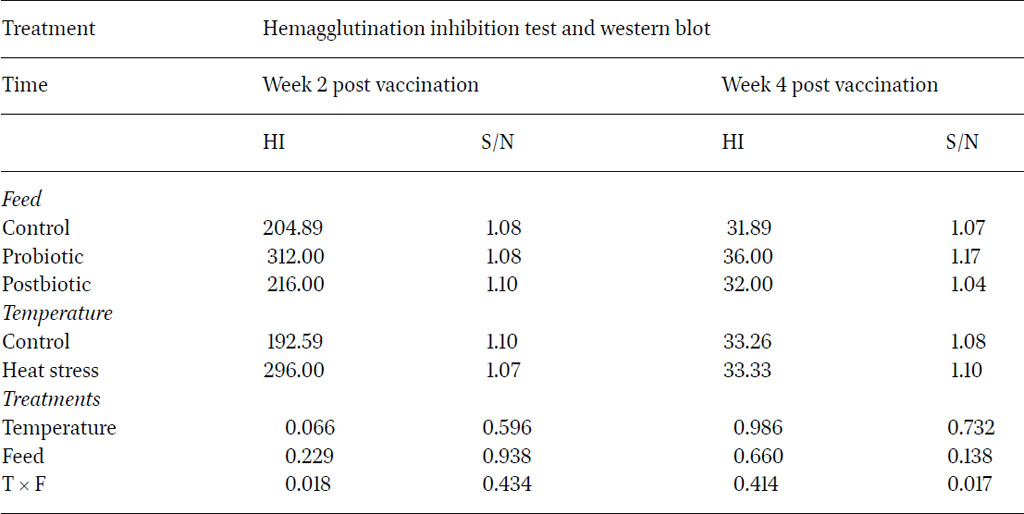
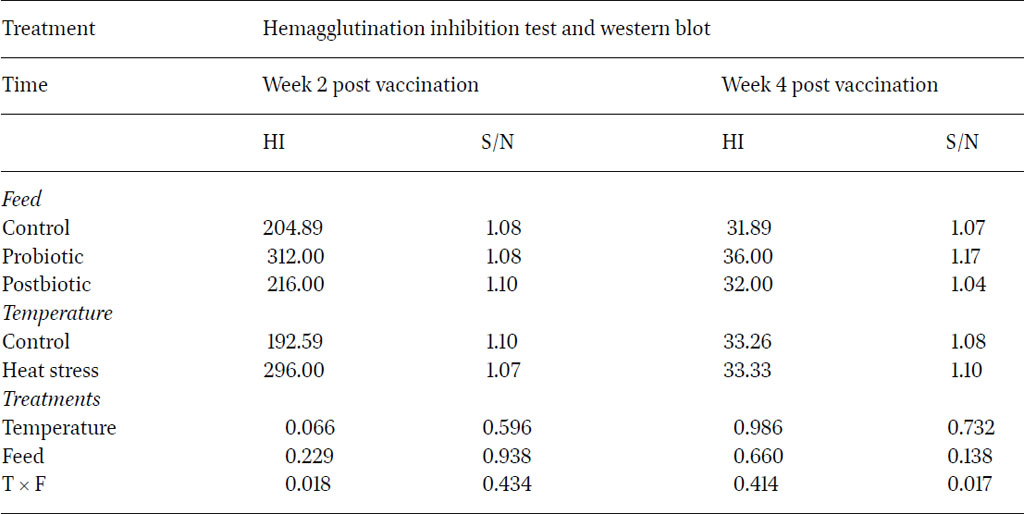
To determine the effect of probiotic and postbiotic supplementation on humoral immunity of layers under heat stress conditions, serum samples were tested using haemagglutination inhibition (HI) method to depict antibody production in response to live NDV vaccine. The immune response results at two and four weeks post vaccination against NDV vaccine are shown in Table 8. No significant differences were observed for HI titres against NDV vaccine across treatments. Although the results were not significant at two and four weeks post vaccination with NDV, the HI titre from the probiotic and postbiotic groups were numerically higher than the control group. The results at week two post vaccination ranged between 192.59 and 312 titre and decreased after four weeks post vaccination to range between 31.89 and 36.00. This decline of HI titre after four weeks post vaccination agreed with previous trials, which reported a maximal antibody titre obtained at two weeks post vaccination (Saif and Fadly, 2008). The magnitude of humoral response depended on the vaccine/infecting strain, bird age and breed and the environmental conditions.
Haemagglutination inhibition titres and S/N ratios of western blotting of bird sera under different temperature and feed parameters during five months of experimental period
Citation: Journal of Applied Animal Nutrition 13, 2 (2025) ; 10.1163/2049257x-20251017
The significant interaction between the type of feed and the temperature conditions at two weeks post NDV vaccination (P<0.05) clearly delineated the positive impact of dietary L. plantarum probiotics on the improvement of the humoral immune response under heat stress conditions. The results obtained in this study did not provide compelling evidence to suggest that dietary postbiotics derived from L. plantarum had a significant impact on humoral immunity in poultry. Humam et al. (2019) reported a slight efficacy in utilising L. plantarum RI11 post-biotics in comparison to RS5 metabolites, used in this study, to improve IgG and IgM antibody levels in heat stressed broilers. Nevertheless, previous studies reported that dietary pro-and post-biotics alone were not able to induce a significant humoral immunity improvement in heat stressed birds (Rahimi et al., 2006; Humam et al., 2019). Their impact was improved by the inclusion of other dietary additives, such as inulin. This shows that the role of dietary probiotics and post-biotics as immunopotentiators of the humoral response is debatable.
Humoral immunity against NDV fusion
In Western blots, the signal-to-noise (S/N) ratio is determined by comparing the signal intensity to the noise in the local background for each band present on the blot. When a blot exhibited bands of relatively low intensity, a higher S/N ratio indicated that these are more likely to be discernible and, consequently, potentially measurable, in contrast to a scenario with a low S/N ratio. The results showed no significant difference in S/N ratio against NDV-F protein between the control and treatment groups at two and four weeks post vaccination (Table 8). As for the value of the post-biotic group, this had higher S/N (1.1) than the control group (1.08) two weeks post vaccination, whereas the probiotic group showed greater (1.17) S/N ratio compared to control group (1.07) at four weeks after vaccination. The statistical analysis of the quantitative seroconversions for antigen F (fusion protein) from NDV, revealed complete failure of each factor separately to enhance seroconversion to the protective F‐protein from NDV in birds (P>0.05). However, there was a significant impact of the interaction of the two factors included in this study on the improvement of the S/N ratio against the F protein whereby probiotics clearly enhanced the specific immune response against this specific viral protein at four weeks post vaccination under heat stress conditions (P<0.05).
Positive HI titres against the haemagglutinin protein coupled with the low antibody titres against the F protein question the protective efficacy of the vaccine against velogenic NDV strains. The importance of obtaining a significant seroconversion to the F‐protein is due to its role in the pathogenesis of ND, in which it assists the virus to be involved in cell fusion leading to penetration of the host cell, a prerequisite for multiplication (Saif and Fadly, 2008).
These results will allow future investigation and evaluation of specific NDV vaccines to provide protective immune response against highly velogenic NDV strains circulating locally. This evaluation should not be only based on ELISA or HI tests, but should include WBC analysis as well.
4 Conclusions
This research revealed that heat stress adversely affected blood parameters, particularly RBC, HGB, HCT, MCV, PLT, MPV and PCT in layers subjected to cyclic heat stress. These parameters showed a significant (P<0.05) decrease compared to the control groups after two and five months of the trial.
Pro- and post-biotic supplementation numerically improved lesions scores attributed to heat stress, manifested by the mitigation of mucosal cell degeneration and microvilli morphology alteration, along with an increase in the number of villi. The impact of L. plantarum pro- and post-biotics was not significant for humoral immunity, specifically against the F protein from NDV.
The postbiotic metabolite showed promise as a feed additive particularly under heat stress conditions. However, it should not be viewed as a complete replacement for chemical therapeutics. In addition, the paucity of significant differences in various immunity parameters between heat stressed and control birds, suggested new heat stress models may be needed that include different temperature, cycling hours and layer breeds.
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
Abd El-Ghany, W.A., Abdel-Latif, M.A., Hosny, F., Alatfeehy, N.M., Noreldin, A.E., Quesnell, R.R., Chapman, R., Sakai, L. and Elbestawy, A.R., 2022. Comparative efficacy of postbiotic, probiotic and antibiotic against necrotic enteritis in broiler chickens. Poultry Science 101(8): 101988. https://doi.org/10.1016/j.psj.2022.101988
Abudabos, A.M., Al-Owaimer, A.N., Hussein, E.O., Ali, M.H. and Al-Ghadi, M.Q., 2018. Effect of natural ascorbic acid on performance and certain haemato-biochemical values in layers exposed to heat stress. Journal of Animal and Plant Sciences 28(2): 441–448. https://doi.org.10.17582/journal.pjz/2018.50.3.951.955
Ahmad, R., Yu, Y.H., Hsiao, F.S., Su, C.H., Liu, H.C., Tobin, I., Zhang, G. and Cheng, Y.H. 2022. Influence of heat stress on poultry growth performance, intestinal inflammation and immune function and potential mitigation by probiotics. Animals 12(17): 2297. https://doi.org.10.3390/ani12172297
Altan, O., Altan, A., Cabuk, M. and Bayraktar, H., 2000. Effects of heat stress on some blood parameters in broilers. Turkish Journal of Veterinary and Animal Sciences 24: 145–148. https://journals.tubitak.gov.tr/veterinary/vol24/iss2/8
Awad, E.A., Idrus, Z., Soleimani Farjam, A., Bello, A.U. and Jahromi, M.F., 2018. Growth performance, duodenal morphology and the caecal microbial population in female broiler chickens fed glycine-fortified low protein diets under heat stress conditions. British Poultry Science 59: 340–348. https://doi.org/10.1080/00071668.2018.1440377
Bounous, D. and Stedman, N., 2000. Normal avian hematology: chicken and turkey. In: Feldman, B.F., Zinkl, J.G. and Jain, N.C. (eds.) Schalm’s Veterinary Hematology. Wiley, New York, NY, USA, pp. 1147–1154.
Bueno, J.P.R., de Mattos Nascimento, M.R.B., da Silva Martins, J.M., Marchini, C.F.P., Gotardo, L.R.M., de Sousa, G.M.R., Mundim, A.V., Guimarães, E.C. and Rinaldi, F.P., 2017. Effect of age and cyclical heat stress on the serum biochemical profile of broiler chickens. Semina: Ciências Agrárias 38(3): 1383–1392. https://doi.org.10.5433/1679-0359.2017v38n3p1383
Burkholder, K.M., Thompson, K.L., Einstein, M.E., Applegate, T.J. and Patterson, J.A., 2008. Influence of stressors on normal intestinal microbiota, intestinal morphology and susceptibility to Salmonella enteritidis colonization in broilers. Poultry Science 87: 1734–1741. https://doi.org.10.3382/ps.2008-00107
Byung-Sung, P., Um, K., Park, S. and Zammit, V.A., 2018. Effect of stocking density on behavioral traits, blood biochemical parameters and immune responses in meat ducks exposed to heat stress. Archiv für Tierzucht 61(4): 425–432. https://doi.org/10.5194/aab-61-425-2018
Danladi, Y., Loh, T.C., Foo, H.L., Akit, H., Tamrin, N.A.Md. and Naeem Azizi, M., 2022. Effects of postbiotics and paraprobiotics as replacements for antibiotics on growth performance, carcass characteristics, small intestine histomorphology, immune status and hepatic growth gene expression in broiler chickens. Animals 12(7): 917. https://doi.org/10.3390/ani12070917
Dinu, C., Pârvu, M., Cureu, I. and Brăslaşu, M.C., 2004. Interaction between the diet, temperature and some blood constituents in Roso hens. Archivos de Zootecnia 7: 27–32.
Farran, M., El Masry, B., Kaouk, Z. and Shaib, H., 2024. Impact of dietary Lactobacillus plantarum postbiotics on the performance of layer hens under heat stress conditions. Open Journal of Veterinary Medicine 14: 39–55. https://doi.org.10.4236/ojvm.2024.143004
Gadde, U., Oh, S.T., Lee, Y.S., Davis, E., Zimmerman, N., Rehberger, T. and Lillehoj, H., 2017. The effects of direct-fed microbial supplementation, as an alternative to antibiotics, on growth performance, intestinal immune status and epithelial barrier gene expression in broiler chickens. Probiotics and Antimicrobial Proteins 9: 397–405. https://doi.org.10.1007/s12602-017-9275-9
Hu, Z. 2024. Probiotics: common genera, mechanisms, current phase and the future. Highlights in Science, Engineering and Technology 109: 204–224. https://doi.org/10.54097/gajke929
Humam, A.M., Loh, T.C., Foo, H.L., Samsudin, A.A., Mustapha, N.M., Zulkifli, I. and Izuddin, W.I., 2019. Effects of feeding different postbiotics produced by Lactobacillus plantarum on growth performance, carcass yield, intestinal morphology, gut microbiota composition, immune status and growth gene expression in broilers under heat stress. Animals 9(9): 644.
Imperial, I.C. and Ibana, J.A. 2016. Addressing the antibiotic resistance problem with probiotics: reducing the risk of its double-edged sword effect. Frontiers in Microbiology 7: 1983. https://doi.org/10.3389/fmicb.2016.01983
Jain, N.C., 1993. Essentials of veterinary hematology. Lea and Febringers, Philadelphia, PA, USA, pp. 365–372.
Kareem, K.Y., Ling, F.H., Chwen, L.H., Foong, O.M. and Asmara, S.A., 2014. Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathogens 6(1): 23. https://doi.org.10.1186/1757-4749-6-23
Köseman, A., Akdemir, F., Üremiş, N., Şeker, İ. and Türköz, Y., 2021. Effects of Alchemilla vulgaris on haematology and antioxidant status of heatstressed quails during the late laying period. Journal of the Hellenic Veterinary Medical Society 72(4): 3463–3472. https://doi.org/10.12681/jhvms.29396
Kubena, L.F., May, J.D., Reece, F.N. and Deaton, J.W., 1972. Hematocrit and hemoglobin levels of broilers as influenced by environmental temperature and dietary iron level. Poultry Science 51: 759–763. https://doi.org/10.3382/ps.0510759
Liu, X., Zhao, H. and Wong, A. 2024. Accounting for the health risk of probiotics. Heliyon 10(6): e27908. https://doi.org/10.1016/j.heliyon.2024.e27908
Loh, T.C., 2017. Animal feed, the way forward. Penerbit UPM, Seri Kembangan, Malaysia.
Loh, T.C., Choe, D.W., Foo, H.L., Sazili, A.Q. and Bejo, M.H., 2014. Effects of feeding different postbiotic metabolite combinations produced by Lactobacillus plantarum strains on egg quality and production performance, faecal parameters and plasma cholesterol in laying hens. BMC Veterinary Research. 10: 149. https://doi.org/10.1186/1746-6148-10-149
Mahmoud, U.T., Abdel-Rahman, M.A., Darwish, M.H.A. and Mosaad, G.M., 2013. The effect of heat stress on blood picture of Japanese quail. Journal of Advanced Veterinary Research 3(2): 69–76. https://advetresearch.com/index.php/AVR/article/view/118
Mashaly, M.M., Hendricks, G.L., Kalama, M.A., Gehad, A.E., Abbas, A.O. and Patterson, P.H., 2004. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poultry Science 83(6): 889–894. https://doi.org/10.1093/ps/83.6.889
Naeem, M. and Bourassa, D. 2025. Probiotics in poultry: unlocking productivity through microbiome modulation and gut health. Microorganisms 13(2): 257. https://doi.org/10.3390/microorganisms13020257
Nanto-Hara, F., Kikusato, M., Ohwada, S. and Toyomizu, M., 2020. Heat stress directly affects intestinal integrity in broiler chickens. Journal of Poultry Science 57(4): 284–290. https://doi.org/10.2141/jpsa.0190004
OIE/FAO, 2020. Haemagglutination inhibition (HI) tests for paramyxoviruses and orthomyxoviruses (PMV and Flu), infectious bronchitis virus (IBV) and egg drop syndrome (EDS). SOP: VI.441 Edition 21. 13 pages
Osadcha, Y., 2023. Haematological parameters of laying hens under different group size. Anatomia, Histologia, Embryologia 52: 919–926. https://doi.org/10.1111/ahe.12951
Park, S.O., Park B.S. and Hwangbo, J., 2015. Effect of cool water and inverse lighting on growth performance of broiler chickens under extreme heat stress. Journal of Environmental Biology 36: 865–873. https://pubmed.ncbi.nlm.nih.gov/26364463/
Percie du Sert, N., Ahluwalia, A., Alam, S., Avey, M.T., Baker, M., Browne, W.J., Clark, A., Cuthill, I.C., Dirnagl, U., Emerson, M., Garner, P., Holgate, S.T., Howells, D.W., Hurst, V., Karp, N.A., Lazic, S.E., Lidster, K., MacCallum, C.J., Macleod, M., Pearl, E.J., Petersen, O.H., Rawle, F., Reynolds, P., Rooney, K., Sena, E.S., Silberberg, S.D., Steckler, T. and Wurbel, H., 2020. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biology 18: e3000411 https://doi.org/10.1371/journal.pbio.3000411
Rahimi, S.H. and Khaksefidi, A., 2006. A comparison between the effects of a probiotic (Bioplus 2B) and an antibiotic (virgenia mycin) on the performance of broiler chicken under heat stress. Iranian Journal of Veterinary Research 7: 48–56. https://doi.org/10.22099/ijvr.2006.2645
Rostagno, M.H., 2020. Effects of heat stress on the gut health of poultry. Journal of Animal Science 98(4): skaa090. https://doi.org.10.1093/jas/skaa090
Saeed, M., Afzal, Z., Afzal, F., Khan, R.U., Elnesr, S.S., Alagawany, M. and Chen, H. 2023. Use of postbiotic as growth promoter in poultry industry: A review of current knowledge and future prospects. Food Science of Animal Resources 43(6): 1111–1127. https://doi.org/10.5851/kosfa.2023.e52
Saif, Y.M. and Fadly, A.M., 2008. Diseases of Poultry, 12th edition. Blackwell Pub., Ames, IA, USA.
Shaib, H.A., 2004. PCR use in epidemiological study of avian mycoplasmosis and control of gumboro by herbal extracts. MSc dissertation, Department of Animal Sciences, American University of Beirut, Beirut, Lebanon.
Shenderov, B.A., 2013. Metabiotics: Novel idea or natural development of probiotic conception. Microbial Ecology in Health and Disease 24: 20399. https://doi.org.10.3402/mehd.v24i0.20399
Shigwedha, N., Sichel, L., Jia, L. and Zhang, L., 2014. Probiotical cell fragments (PCFs) as ‘novel nutraceutical ingredients’. Journal of Biosciences and Medicines 2: 43–55. https://doi.org.10.4236/jbm.2014.23007
Shojadoost B., Alizadeh M., Boodhoo N., Astill J., Karimi S.H., Doost J.S., Taha-Abdelaziz, K., Kulkarni, R. and Sharif, S., 2022. Effects of treatment with Lactobacilli on necrotic enteritis in broiler chickens [e-pub ahead of print]. Probiotics and Antimicrobial Proteins 14(6): 1110–1129. https://doi.org/10.1007/s12602-021-09901-5
Terpou, A., Papadaki, A., Lappa, I.K., Kachrimanidou, V., Bosnea, L.A. and Kopsahelis, N. 2019. Probiotics in food systems: Significance and emerging strategies towards improved viability and delivery of enhanced beneficial value. Nutrients 11(7): 1591. https://doi.org/10.3390/nu11071591.
Trisnawita, Y., Silalahi, J. and Morin Sinaga, S. 2018. The effect of storage condition on viability of lactic acid bacteria in probiotic product. Asian Journal of Pharmaceutical and Clinical Research 11: 84–86. http://dx.doi.org/10.22159/ajpcr.2018.v11s1.26574
Vo, K.V., Boone, M. A. and Torrence, A.K., 1978. Electrolyte content of blood and bone in chickens subjected to heat stress. Poultry Science 57(2): 542–544. https://doi.org/10.3382/ps.0570542
Wang, J., Ji, H., Wang, S., Liu, H., Zhang, W., Zhang, D. and Wang, Y., 2018. Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Frontiers in Microbiology 9: 1953. https://doi.org/10.3389/fmicb.2018.01953
Xu, T.; Chen, Y.; Yu, L.; Wang, J.; Huang, M. and Zhu, N., 2020. Effects of Lactobacillus plantarum on intestinal integrity and immune responses of egg-laying chickens infected with Clostridium perfringens under the free-range or the specific pathogen free environment. BMC Veterinary Research 16: 47. https://doi.org/10.1186/s12917-020-2264-3
Yelin, I., Flett, K.B., Merakou, C., Mehrotra, P., Stam, J. and Snesrud, E., 2019. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nature Medicine 25: 1728–1732. https://doi.org.10.1038/s41591-019-0626-9
Zhang, L., Zhang, L., Zhan, X., Zeng, X., Zhou, L., Cao, G., Chen, A. and Yang, C., 2016. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. Journal of Animal Science and Biotechnology 7: 3. https://10.1186/s40104-016-0061-4
Zhong, Y., Wang, S., Di, H., Deng Z., Liu, J. and Wang, H., 2022. Gut health benefit and application of postbiotics in animal production. Journal of Animal Science and Biotechnology 13: 38. https://doi.org/10.1186/s40104-022-00688-1
Zulkifli, I., Norma, M.T., Israf, D.A. and Omar, A.R., 2000. The effect of early age feed restriction on subsequent response to high environmental temperatures in female broiler chickens. Poultry Science 79: 1401–1407. https://doi.org/10.1093/ps/79.10.1401








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}