Abstract
Yeast cell wall (YCW) prebiotics from Saccharomyces cerevisiae, can be added to the diet of animals as an effective decoy molecule for Gram negative bacteria, helping to prevent colonisation of the gut by disease-causing organisms. However, the form and processing of YCW can affect its efficacy in preventing attachment of pathogens to the gut wall and, hence, colonisation. Several commercial yeast cell wall products, A, B and C, including a mannan rich fraction (MRF) product, were assessed for their ability to interfere with the attachment of Salmonella spp. to intestinal IPEC-J2 cells. Attachment of three serovars S. enterica Enteritidis, S. enterica Heidelberg, and S. enterica Typhimurium to IPEC-J2 cells was consistently inhibited with addition of MRF and, to a lesser extent, products B and C, while product A resulted in a greater attachment compared to the control. The reduced attachment of the three Salmonella spp. serovars to IPEC-J2 cells in the presence of MRF coincided with lower gene expression and protein of inflammatory markers interleukin (IL)-1
1 Introduction
Salmonella spp. are a leading cause of bacterial foodborne illness for human consumers, second only to Campylobacter spp. (EFSA, ECDC, 2018; WHO, 2016) although salmonellosis persists as a leading foodborne infection from animal products, including pork (Mkangara et al., 2023) and eggs (CDC, Foodnet Fast Data, 2023; EFSA, ECDC, 2018). Humans can become infected if they consume animal products that are contaminated with the bacterium. Salmonella enterica (SE) serotypes are persistent in pig production with geographical regions affected by different serotypes. For example, of all SE, S. Typhimurium and S. Derby are the most predominant in North American and Oceania (Ferrari et al., 2019; Hendriksen et al., 2011). However, S. Enteritidis is ranked as the most common serovar in Europe, S. Meleagridis serovar and S. Hardar predominate in South America and Africa respectively (Ferrari et al., 2019).
To date most of the research on SE prevalence has focussed on the fattening stage of pigs (Casanova-Higes et al., 2016; Hill et al., 2016) although early intervention by the producer at weaning may reduce pathogen load at slaughter (Casanova-Higes et al., 2019). In recent years pork meat and its derived products have become one of the most significant sources of infection as pork is one of the most consumed and traded meat products worldwide. The SE serotypes that mainly cause infections in humans include S. Enteritidis, S. Typhimurium and its monophasic variant S. Heidelberg (Campos et al., 2019).
Sub-therapeutic antibiotic usage in pig diets previously prevented infections, which promoted growth. However, due to the emergence and spread of antibiotic resistance, the EU banned the use of antibiotics in livestock feed in 2006 (IP/05/1687, EU, 2005). This was followed by the restriction on zinc oxide usage in 2022 to non-therapeutic levels of 150 ppm (EMA/394961/2017), which has led to greater demand for effective alternatives. These included acidifiers, probiotics and prebiotics, with the latter becoming an increasingly effective alternative to zinc oxide. Prebiotics in the form of mannan-rich fractions (MRF), derived from the yeast cell wall (YCW) of Saccharomyces cerevisiae, can be added to the diet of animals as an effective decoy molecule for Gram negative bacteria (Browne et al., 2019; Healy and Horgan 2017). Pathogenic bacteria need to bind to the gut wall in order to proliferate and colonise the gut, where their numbers then cause disease. The MRF binds to pathogenic bacteria which subverts the normal bacterial attachment to the host intestinal cells (Haas and Nochta, 2012). This mode of binding specificity, where mannose carbohydrate binds to the surface lectins of enteric bacteria, such as S. Typhimurium, has been previously described by Firon et al. (1983).
The following study was conducted to assess if foodborne strains of Salmonella spp. incubated in the presence of prebiotic YCW products could alter attachment ability of the bacterium to IPEC-J2 cells. The use of the intestinal pig epithelial cell line (IPEC-J2) has previously been proven to be a useful model to examine the attachment and invasion process of Salmonella (Grzymajlo et al., 2017; Healy and Horgan, 2017) and, hence, was utilised in the study.
2 Materials and methods
Salmonella culture conditions
Three serovars of SE; S. Enteritidis (NCTC 12694, UK), S. Heidelberg (ATCC, 8326, USA) and S. Typhimurium (KCTC, 2514, Korea Research Institute of Bioscience) were used for the duration of these experiments. The cells were grown in liquid nutrient broth (Avantor, Difco™ BD, Franklin Lakes, NJ, USA) at 37 °C and 150 rpm overnight. The cultures were plated on nutrient broth agar at 37 °C overnight and stored at 4 °C. Prior to sub-culturing, plates were stored for a maximum of 14 d. The cultures were washed in 10 mM phosphate buffered saline solution (PBS; Oxoid Ltd, Basingstoke, UK, BR0014G). Optical densities of Salmonella were measured using a UV spectrophotometer set at 650 nm and adjusted to an OD of 0.2 (1×108 cfu/ml) for attachment assessments.
Mammalian cell culture
Sterile high glucose Dulbecco’s modified eagle medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA, D6546) was prepared using DMEM high glucose, 10% (v/v) foetal bovine serum (FBS) and 1% (v/v) L-glutamine Gibco® (Grand Island, NY, USA). A stock of IPEC-J2 cells, at passage 20, was taken up from frozen. The cells were split using trypsin-EDTA and were kept at 37 °C with a 5% CO2 atmosphere. The cells were seeded onto 12-well cell culture plates at a density of 1×105 cells per well. The plate was placed in a 37 °C incubator with a 5% CO2 atmosphere overnight to ensure the cells attached to the surface of the plate.
For the attachment assessment, six wells of the 12-well cell culture plate were labelled for as Control, MRF and products A, B and C. The wells were seeded at 1×105 IPEC-J2 cells per well and cells were left to adhere to the plate in the 37 °C incubator overnight in 5% CO2. The following day media was removed and IPEC-J2 cells were washed with pre-warmed CO2-independent media, supplemented with 10% (v/v) FBS and 1% (v/v) L-glutamine. The CO2 dependent media was removed and replaced with CO2 independent media containing 1×108 cfu/ml SE alone (Control) or with treatments MRF, product A, product B or product C (16 mg/ml). Once the treatments were added to their corresponding wells, the plate was covered and placed in 37 °C incubator for 1 h.
After 1 h the media was removed from each well and retained for later analysis. The IPEC-J2 cell monolayer was washed a further three times with 1 ml CO2 independent media, prior to lysing the intestinal cells with the addition of 300 μl of sterile TritonX (Sigma-Aldrich, X-114) (1% v/v) PBS to each well, this was incubated for 5 min at room temperature followed by pipette mixing. The cell suspension was diluted with PBS and 100 μl from each treatment was plated on agar plates and incubated at 37 °C overnight. Colonies on each plate were enumerated using the plate count method to determine the number of Salmonella cells that adhered to the IPEC-J2 cells (Healy and Horgan, 2017).
Quantification of inflammatory markers using ELISA
The ELISA from AssayGenie were performed to measure secreted tumour necrosis factor (TNF)-
Gene expression analysis
The IPEC-J2 cells grown in the presence and absence of YCW products plus SE challenge were suspended in 350 μl of RTL lysis buffer (Qiagen, Hilden, Germany, 74004). The cell suspension was ruptured for 30 s with a tissue rupture probe on ice to generate a homogenous mixture. The RNA was isolated following the RNeasy Micro Kit (Qiagen, 74004) procedure as detailed in the user manual. RNA quality was determined by Qubit 4 Fluorometer (Invitrogen, Carlsbad, CA, USA) with IQ values above 6.5 utilised and RNA concentrations ˃1.8
Reverse transcription was performed on RNA acting as a template to synthesise cDNA using SuperScript® III First-Strand Synthesis System (Invitrogen, 11752–050) prior to PCR analysis being performed. Real-time PCR was performed in triplicate using a two-step cycling programme, consisting of a heat activation step (95oC for 10 min) and a cycling step (40 cycles, 95 °C for 15 s and 60 °C for 1 min) (ABI 7500 Fast; Applied Biosystems, Foster City, CA, USA). The PCR analysis of gene expression change associated with inflammatory associated markers IL-8, IL-1

PCR primers for IPEC-J2 cells.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
Statistical analysis
Statistical analysis of adhesion experiments was carried out using PRISM software employing One-way ANOVA followed by Tukey’s post-hoc test, with results determined significant at an alpha threshold of P≤0.05. Bar charts are represented as the mean average and standard error mean.
3 Results
S. Enteritidis attachment to IPEC-J2 cells
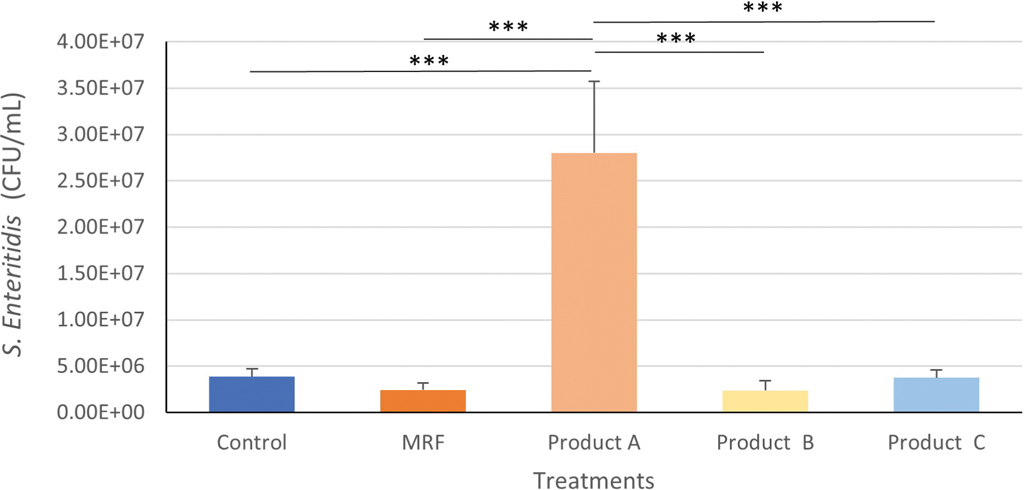
The SE attachment to IPEC-J2 cells was significantly higher with addition of product A (P≤0.001) compared to all other treatments and both controls (Figure 1).
Attachment of Salmonella Enteritidis to IPEC-J2 cells treated with YCW products. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.001 ***.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
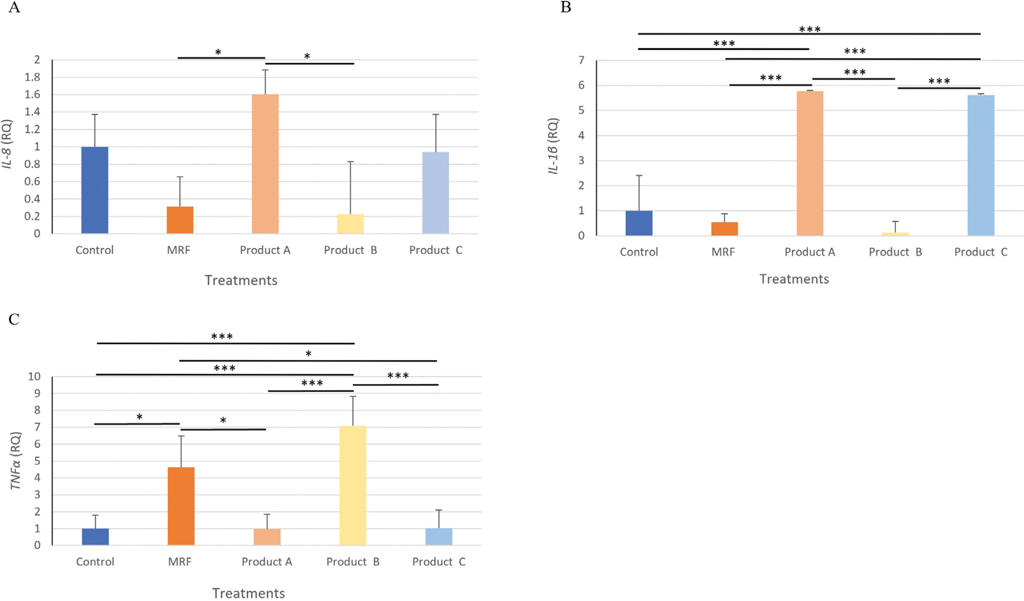
Inflammatory protein changes in response to Salmonella Enteritidis exposure to IPEC-J2 cells
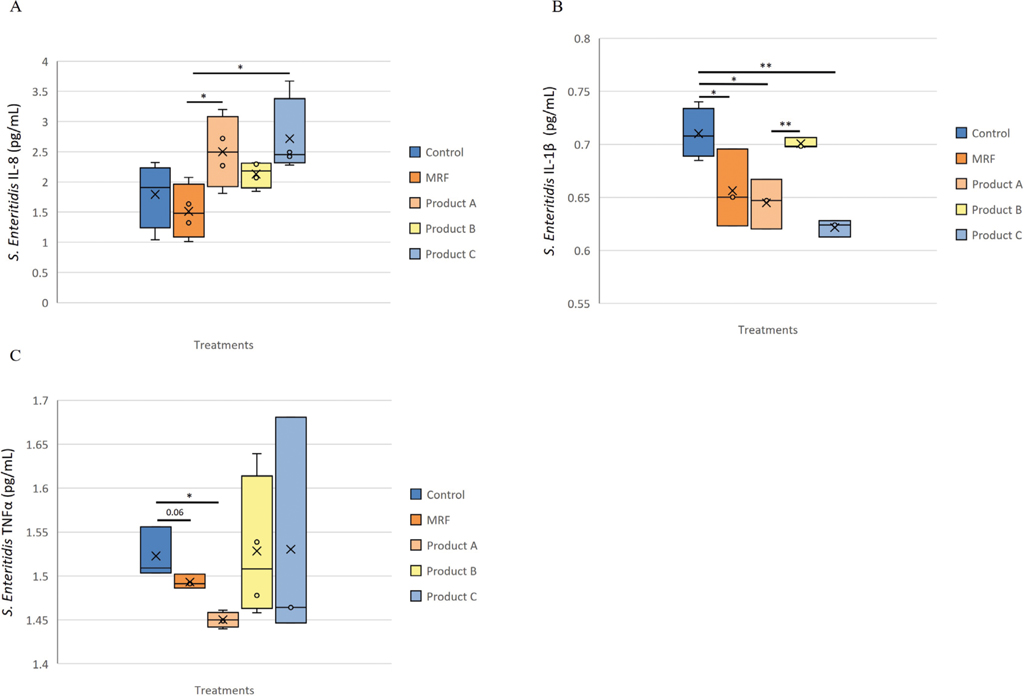
Secretion of IL-8 was significantly (P≤0.05) higher with product A and product C compared to MRF but not the Control (Figure 2A). Application of MRF resulted in significantly (P≤0.05) lower secretion of IL-1
Inflammatory protein response of IPEC-J2 cells exposed to Salmonella Enteritidis and supplemented with YCW treatment, A (IL-8), B (IL-1β ) and C (TNF-α ). Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.05 *, P≤0.01 **
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
Inflammatory gene expression changes in response to Salmonella Enteritidis exposure to IPEC-J2 cells
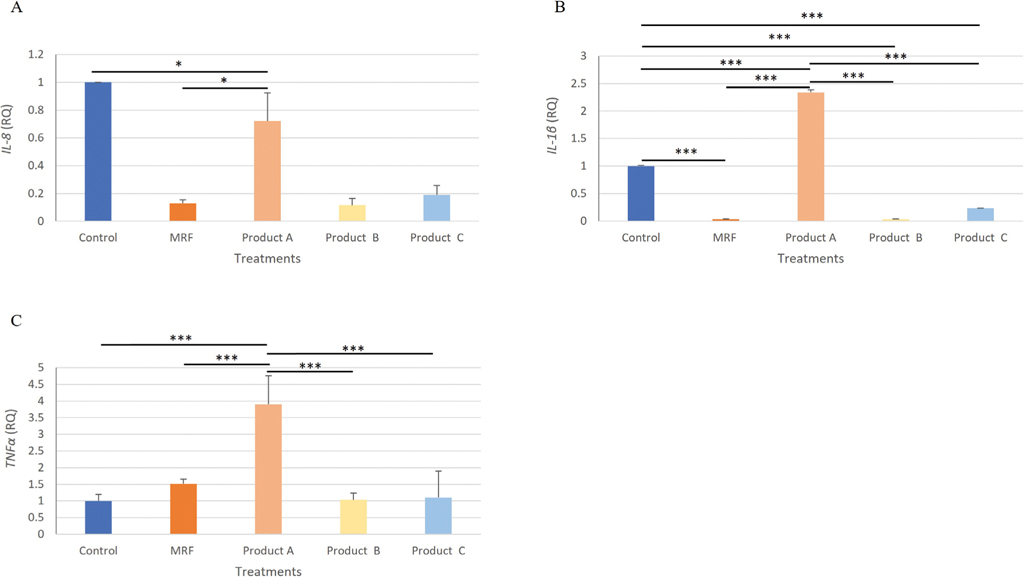
The expression of chemokine IL-8 from SE challenged IPEC-J2 cells was significantly (P≤0.05) lower in the MRF and product B treated cells compared to the Control (Figure 3A). Expression of Il-1
Gene expression analysis of IL-8 (A), IL-1β (B) TNF-α (C), IPEC-J2 cells exposed to Salmonella Enteritidis and treated with YCW treatment. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, n=4, P≤0.05 * and P≤0.001 ***.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
Salmonella Heidelberg attachment to IPEC-J2 cells
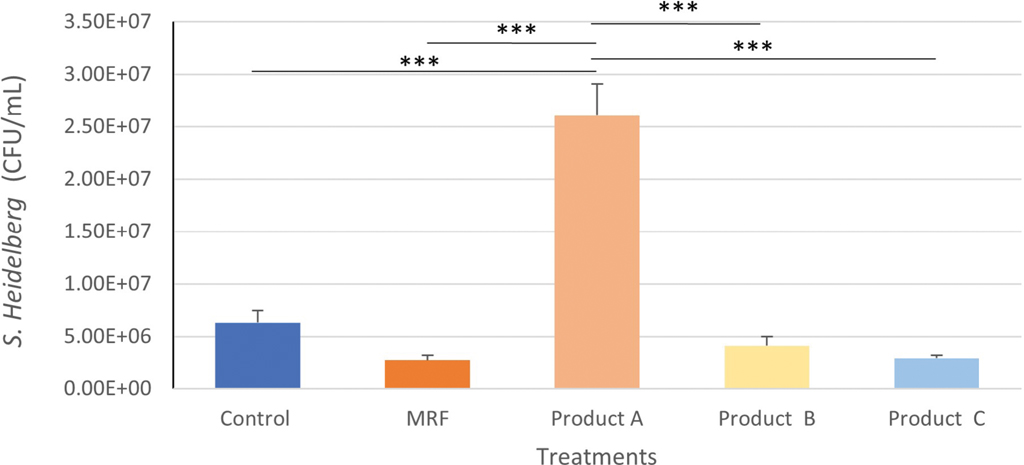
The inclusion of product A to IPEC-J2 cells resulted in significantly greater (P≤0.001) attachment of S. Heidelberg compared with the Control, MRF, product B and product C (Figure 4).
Attachment of Salmonella Heidelberg to IPEC-J2 cells treated with YCW products. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.001 ***.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
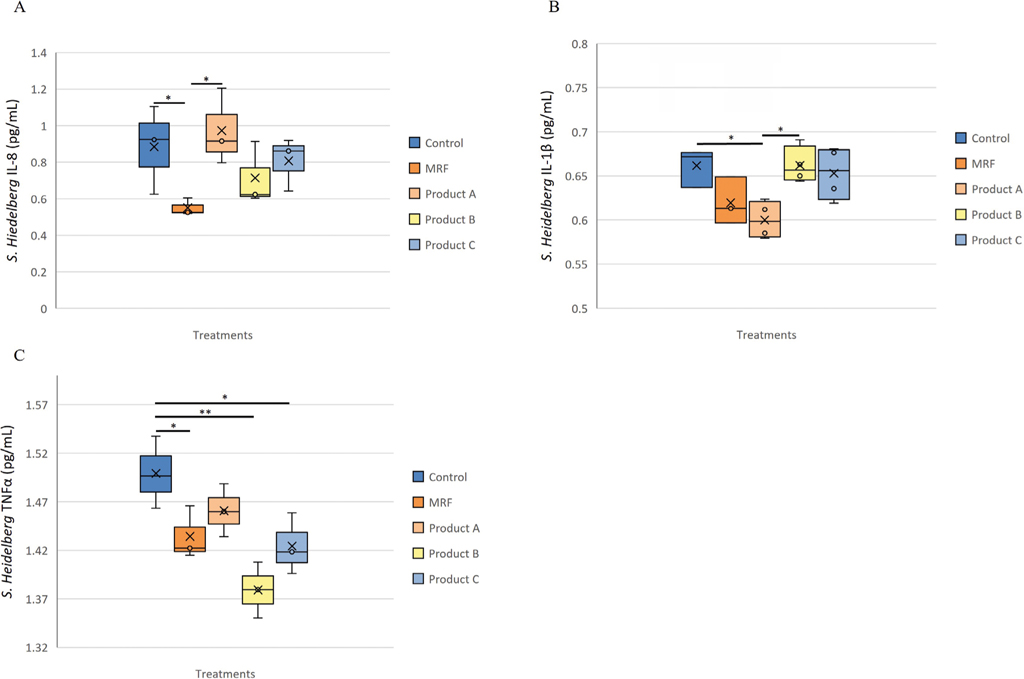
Inflammatory protein changes in response to Salmonella Heidelberg exposure to IPEC-J2 cells
Addition of MRF to S. Heidelberg challenged IPEC-J2 cells lead to significantly (P≤0.05) lower IL-8 (Figure 5A) compared to the Control and product A. Challenged IPEC-J2 cells receiving product A had significantly (P≤0.05) lower IL-1
Inflammatory protein response of IPEC-J2 cells exposed to Salmonella Heidelberg and supplemented with YCW treatment, A (IL-8), B (IL-1β ) and C (TNF-α ). Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.05 *, P≤0.01 **.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
Inflammatory gene expression changes in response to Salmonella Heidelberg exposure to IPEC-J2 cells
Expression of the IL-8 gene was significantly lower for both IPEC-J2 cells receiving MRF (P≤0.05) and product B (P≤0.05) compared to product A (Figure 6A) but showed a similar trend to IL-8 protein abundance (Figure 5A). Gene IL-1
Gene expression analysis of IL-8 (A), IL-1β (B) TNF-α (C), IPEC-J2 cells exposed to Salmonella Heidelberg and treated with YCW treatment. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, n=4, P≤0.05 *, P≤0.01 ** and P≤0.001 ***.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
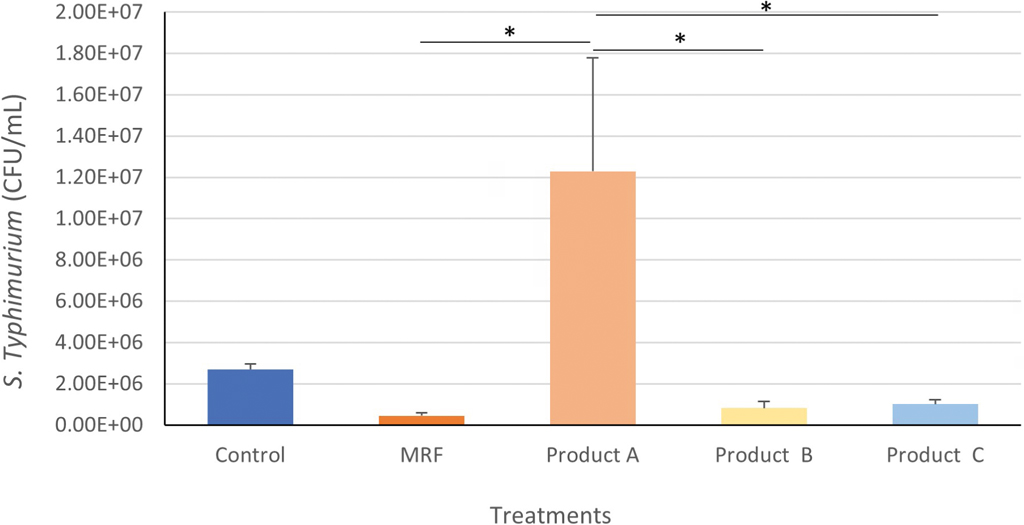
Salmonella Typhimurium attachment to IPEC-J2 cells
Product A resulted in a significantly (P≤0.05) greater number of attached S. Typhimurium cells to IPEC-J2 cells compared to all other treatments except for the Control (Figure 7). The application of MRF led to the lowest number of S. Typhimurium cells attached to IPEC-J2 intestinal cells (4.5x105) compared to the control (2.7×106), product A (1.2x107), B (8.2×105) and C (1.0×106).
Attachment of Salmonella Typhimurium to IPEC-J2 cells treated with YCW products. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.05 *.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
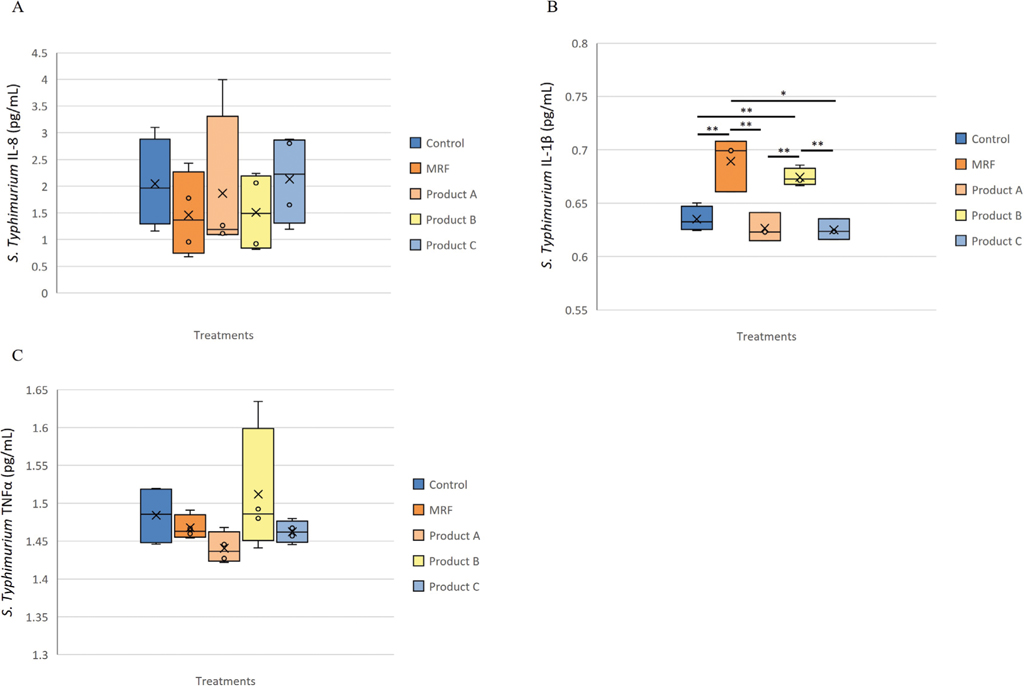
Inflammatory protein changes in response to Salmonella Typhimurium exposure to IPEC-J2 cells
No significant change was observed in IL-8 and TNF-
Inflammatory protein response of IPEC-J2 cells exposed to Salmonella Typhimurium and supplemented with YCW treatment, A (IL-8), B (IL-1β ) and C (TNFα ). Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, (n=4), P≤0.05 *, P≤0.01 **.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
Inflammatory gene expression changes in response to Salmonella Typhimurium exposure to IPEC-J2 cells
The S. Typhimurium challenged IPEC-J2 cells had significantly lower gene expression of IL-8 with inclusion of MRF (P≤0.01), product A (P≤0.05), product B (P≤0.001) and product C (P≤0.01) compared to the Control (Figure 9A). The IPEC-J2 cells challenged with S. Typhimurium and treated with MRF or product B demonstrated the lowest gene expression of IL-8, IL-1
Gene expression analysis of IL-8 (A), IL-1β (B) TNF-α (C), IPEC-J2 cells exposed to Salmonella Typhimurium and treated with YCW treatment. Statistical analysis by one-way ANOVA, Tukey’s post-hoc test, n=4, P≤0.05 *, P≤0.01 ** and P≤0.001 ***.
Citation: Journal of Applied Animal Nutrition 13, 1 (2025) ; 10.1163/2049257x-20251015
4 Discussion
Prebiotics such as YCW products offer a solution to mitigate the issue of SE in the pig digestive tract. There are a broad range of YCW products available on the market, most of which are derived from S. cerevisiae. However, product efficacy can vary greatly due to factors such as yeast source, production methods and the yeast fraction utilised. These aspects may impact the ability of YCW products to impact Salmonella attachment. This study evaluated four YCW based products to determine if there were differences in their capacity to lower Salmonella attachment rates. The attachment of S. Enteritidis, S. Heidelberg and S. Typhimurium to IPEC-J2 cells was consistently lower with addition of MRF. This suggested a potential role for MRF in lowering Salmonella colonisation in pigs that consequently could lead to lower risk of transmission to human consumers through contaminated food.
The inclusion of products B and C to IPEC-J2 cells resulted in lower Salmonella attachment, although not to the same extent as MRF, while inclusion of product A, unexpectedly increased attachment of SE strains to IPEC-J2 cells. Further interpretation of the YCW products impact on SE attachment to IPEC-J2 cells required assessment of changes in inflammatory markers. Lower secretion of inflammatory markers IL-1
Furthermore, expression of these three inflammatory genes appeared significantly elevated in response to product A, potentially due to higher attachment of SE to IPEC-J2 cells with product A. Adding MRF and product B resulted in lower inflammatory gene expression, which may have corresponded to lower Salmonella attachment to IPEC-J2 cells. Razzuoli et al. (2017) reported that the lower attachment of SE to IPEC-J2 cells was linked to lower inflammatory markers produced. With exception of TNF-
There is evidence to support that different SE serovars can modulate expression of inflammatory markers from mammalian intestinal cells, including IL-1
The expression of TNF-
Despite this, product A demonstrated an interesting trend in the secretion of both IL-1
5 Conclusions
The addition of MRF and, to a lesser extent, products B and C, impaired the attachment of the three SE strains to IPEC-J2 cells that potentially led to lower inflammatory markers. The SE attachment rates corresponded strongly to changes observed in inflammatory gene expression. Product A demonstrated higher SE attachment to IPEC-J2 cells which corresponded to significantly higher gene expression of inflammatory markers and IL-8 protein secretion when compared to the control. The presence of MRF may offer a means to limit the abundance of foodborne strains of SE found in the pig intestinal tract. The specific composition, variance in production processing and the specific yeast source may have contributed to the heightened effectiveness of MRF compared to the other YCW products.
References
Browne, N., Traynor, A. and Horgan, A.K., 2019. Mannan rich fraction from yeast modulates inflammatory responses in intestinal cells (HT-29) exposed to Escherichia coli. Journal of Applied Animal Nutrition 7: 5.
Campos, J., Mourao, J., Peixe, L. and Antunes, P., 2019. Non-typhoidal Salmonella in the pig production chain: A comprehensive analysis of its impact on human health. Pathogens 8: 1.
Casanova-Higes, A., Andrés-Barranco, S. and Mainar-Jaime, R.C., 2016. Influence of on-farm pig Salmonella status on Salmonella shedding at slaughter. Zoonoses and Public Health 64: 328–336
Casanova-Higes, A., Marín-Alcalá, C.M., Andrés-Barranco, S., Cebollada-Solanas, A., Alvarez, J. and Mainar-Jaime, R.C., 2019. Weaned piglets: another factor to be considered for the control of Salmonella infection in breeding pig farms. Veterinary Research 50: 45.
CDC, Foodborne Diseases Active Surveillance Network (FoodNet), 2023. Pathogen surveliance Salmonella. Available at: https://wwwn.cdc.gov/foodnetfast/
Claud, E.C., Savidge, T. and Walker, W.A., 2003. Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatric Research 53: 419–425.
European Food Safety Authority (EFSA), European Centre for Disease Prevention and Control (ECDC), 2017. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2015. EFSA Journal 15: 4694. https://doi.org/10.2903/j.efsa.2017.4694.
European Medicines Agency (EMA), 2017. Questions and answers on veterinary medicinal products containing zinc oxide to be administered orally to food-producing species. EMA. EMA/394961/2017 Veterinary Medicines Division.
European-Commission (EC), 2005. Ban on antibiotics as growth promoters in animal feed enters into effect. Available at: http://europa.eu/rapid/press-release_IP-05-1687_en.htm.
Ferrari, R.G.F., Rosario, D.K.A., Cunha-Neto, A., Mano, S.B., Figueiredo, E.E.S. and Conte-Junior, C.A., 2019. Worldwide epidemiology of Salmonella serovars in animal-based foods: a eta-analysis. Applied and Environmental Microbiology 85(14): 00591–00519.
Firon, N., Ofek, I. and Sharon, N., 1983. Carbohydrate specificity of the surface lectins of Escherichia coli, Klebsiella pneumoniae, and Salmonella Typhimurium. Carbohydrate Research 120: 235–249.
Grzymajlo, K., Ugorski, M., Suchanski, J., Kedzierska, AE., Kolenda, R., Jarzab, A., et al., 2017. The novel type 1 fimbriae FimH receptor calreticulin plays a role in Salmonella host specificity. Frontiers in Cellular and Infection Microbiology 19: 7.
Halas, V. and Nochta, I., 2012. Mannan oligosaccharides in nursery pig nutrition and their potential mode of action. Animals 23: 261–274.
Haraga, A. and Miller, S.I., 2003. A Salmonella enterica serovar Typhimurium translocated leucine-rich repeat effector protein inhibits NF-κB-dependent gene expression. Infection and Immunity 71(7): 4052–4058.
Hazuda, D.J., Lee, J.C. and Young, P.R., 1988. The kinetics of interleukin 1 secretion from activated monocytes. Differences between interleukin 1 alpha and interleukin 1 beta. JBC 263: 8473–8479. https://doi.org/10.1016/S0021-9258(18)68502-3.
Healy, D. and Horgan, K., 2017. Yeast mannan rich faction reduces adherence of salmonella to porcine intestinal cell line. American Association of Swine Veterinarians 48(1): 1–6.
Hendriksen, R.S., Vieira, A.R., Karlsmose, S., Danilo, M.A., Wong, L.F., Jensen, A.B., Wegener, H.C. and Aarestrup, F.M., 2011. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: results of quality assured laboratories from 2001 to 2007. Foodborne Pathogens and Disease 8(8): 887–900. https://doi.org/10.1089/fpd.2010.0787.
Hill, A.A., Simons, R.R., Kelly, L. and Snary, E.L., 2016. A farm transmission model for Salmonella in pigs, applicable to EU Member States. Risk Analysis 36: 461–481.
Liang, Y., Yang, X.M., Gu, Y.R., Tao, X., Zhong, Z.Z., Gong, J.J., Chen, X.H. and Lv, X.B., 2015. Developmental changes in the expression of the GLUT2 and GLUT4 genes in the longissimus dorsi muscle of Yorkshire and Tibetan pigs. Genetics and Molecular Research 14: 1287–1292.
Mazumder, B., Li, X. and Barik, S., 2010. Translation control: a multifaceted regulator of inflammatory response. Journal of Immunology 184(7): 3311–3319.
Mkangara, M., 2023. Prevention and control of human Salmonella enterica infections: an implication in food safety. International Journal of Food Science 2023: 8899596. https://doi.org/10.1155/2023/8899596
Oliver, J.C., Bland, L.A., Oettinger, C.W., Arduino, M.J., McAllister, S.K., Aguero, S.M. and Favero, M.S., 1993. Cytokine kinetics in an in vitro whole blood model following an endotoxin challenge. Lymphokine and Cytokine Research 12(2): 115–120.
Razzuoli, E., Amadori, M., Lazzara, F., Bilato, D., Ferraris, M., Vito, G. and Ferrari, A., 2017. Salmonella serovar-specific interaction with jejunal epithelial cells. Veterinary Microbiology 207: 219–225.
Spring, P., Wenk, P., Connolly, A.J. and Kiers, A., 2015. A review of 733 published trials on Bio-Mos®, a mannan oligosaccharide, and Actigen®, a second generation mannose rich fraction, on farm and companion animals. Journal of Applied Animal Nutrition 3: 1–11. https://doi.org/10.1017/jan.2015.6
Stecher, B., Robbiani, R., Walker, A.W., Westendorf, A.M., Barthel, M., Kremer, M., Chaffron, S., Macpherson, A.J., Buer, J., Parkhill, J., Dougan, G., Von Mering, C. and Hardtet, W.D., 2007. Salmonella enterica serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biology 5: 2177–2189.
World Health Organization & Food and Agriculture Organization of the United Nations (WHO/FAO), 2016. Interventions for the control of non-typhoidal Salmonella spp. in beef and pork: meeting report and systematic review. World Health Organization, Geneva, Switzerland. Available at: http://www.who.int/iris/handle/10665/249529.
Wu, L., Liao, P., He, L., Ren, W., Yin, J., Duan, J. and Li, T., 2015. Growth performance, serum biochemical profile, jejunal morphology, and the expression of nutrients transporter genes in deoxynivalenol (DON)-challenged growing pigs. BMC Veterinary Research 11: 144.
Zeng, M., Inohara, N. and Nuñez, G., 2017. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunology 10: 18–26.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}