Abstract
Composition of the gut microbiota, and related short chain fatty acids (SCFAs) production can influence allergic status. Extensively hydrolysed casein (eHC) infant formulas are used for management of cow’s milk protein (CMP) allergy; however, effects of eHC on gut microbiota as a potential mechanism contributing to benefits of eHC for infants with allergy are not well understood. This exploratory study aimed to assess the in vitro effects of eHC on infant gut microbial composition and metabolism. Two batches of a specific eHC preparation, along with a mixture of individual amino acids (AA), were first digested in the in vitro TNO Intestinal Model-1 (TIM-1) simulating infant upper gastrointestinal digestion conditions followed by an in vitro colon fermentation model inoculated with faeces from healthy, formula-fed infants. Peptide profile after digestion of the test compounds, as well as effects of the compounds on microbiota community structure and SCFA production were assessed. Overall peptide profiles of the eHC preparations (qualitative, based on liquid chromatography-mass spectrometry) remained largely unaffected by simulated upper gastrointestinal digestion. Faecal fermentation of the specific eHC preparations influenced microbiota composition, with a major impact on Bacteroides abundance. SCFA production was increased beyond control conditions by eHC but not AA. The impact of the AA mixture differed considerably from that of the eHC containing specific peptides. Results suggest that eHC formulas can impact microbiota composition and SCFA production in an in vitro colon fermentation model. While the descriptive results of this study require future confirmation under a more extensive experimental setup, it can be hypothesised that mechanisms modulating microbiota composition and metabolism occurring in infants fed with eHC formula for the management of CMP allergy might contribute to clinical observations related to allergy and tolerance development.
1 Introduction
Overall, almost 2000 bacterial species have been identified in the human gut microbiota (Almeida et al., 2019), with microbiome diversity and number of species varying between individuals (Booijink et al., 2010), across gastrointestinal tract location (Seekatz et al., 2019), and over time (Booijink et al., 2010; Seekatz et al., 2019). Substantial evidence has emerged supporting the importance of gut microbiota in diverse areas of human health, such as development and functioning of the immune system, healthy weight maintenance, and even behaviour and cognition (Clarke et al., 2014; Dethlefsen et al., 2007; Marques et al., 2010). The gut microbial community is also proposed to play a significant role in disease outcomes of inflammatory conditions such as allergies, irritable bowel syndrome, and ulcerative colitis (Clemente et al., 2012; Parekh et al., 2015). Important mediators resulting from microbiome activity are short chain fatty acids (SCFA), such as acetate, propionate, and butyrate. These are produced by bacteria through fermentation of non-digestible complex carbohydrates or are produced from a selected range of amino acids. They are associated with myriad processes, including reduction of asthma and allergy development (Stinson, 2020), as well as immunomodulation and metabolism (Abdalkareem Jasim et al., 2022) and resistance to inflammation (Maslowski et al., 2009).
Infancy and early childhood are particularly important periods of life for microbial community development as health status in later childhood and adult life can be affected by early microbiota programming (Van Best et al., 2015). The gut microbiota in early life is largely dominated by Bifidobacterium, Clostridium and Bacteroides, with low species diversity compared to older children and adults (Marques et al., 2010). Factors such as type of feeding or the introduction of solid foods have a major impact on gut microbiota composition (Marques et al., 2010; Penders et al., 2006).
Emerging evidence suggests a link between a disbalance in gut microbiota composition, bacterial fermentation products, and cow’s milk protein (CMP) allergy (Goldberg et al., 2020; Thompson-Chagoyan et al., 2011). For instance, infants with CMP allergy have a different bacterial community composition than infants without this allergy (Thompson-Chagoyan et al., 2011). Hypoallergenic infant formulas containing extensively hydrolysed casein (eHC) have an important role in the dietary management of CMP allergy in infants where the goal is to avoid allergenic responses towards intact milk protein sequences (Bognanni et al., 2024; Mehta et al., 2022). The peptide profile of an industrial casein hydrolysate can differ from the profile of peptides formed during digestion of intact proteins in the stomach and small intestine. This implies that infants fed formulations containing eHC are likely to be exposed to peptides with different functionalities when compared with infants fed non-hydrolysed formulas or human milk. Studies have found that eHC can modulate T-helper (Th)1/Th2 cytokine expression and enhance expression of the anti-inflammatory, immunoregulatory cytokine interleukin 10 (IL-10) (Aitoro et al., 2017). However, while some guidelines recommend that a hydrolysed formula may offer ’advantages to prevent allergic disease and CMP allergy,’ (Bognanni et al., 2024; Fleischer et al., 2013), other guidelines cite no evidence or recommendation for, or against, such formulas (Fleischer et al., 2021).
EHC typically contains mainly short peptide sequences, such as di- and tri-peptides, and only a small proportion of peptides longer than 10 amino acids (Lambers et al., 2015). While milk-derived bioactive peptides with antimicrobial as well as bifidogenic properties have been identified (Liepke et al., 2002; Shah, 2000), the effects of such hydrolysates on gut microbial communities have not been widely studied to better understand potential mechanisms of action that would benefit CMP allergy. The extent to which eHC-derived peptides might reach the colon without being degraded in the stomach and small intestine is also not well understood. As such, the first aim of the current study was to assess the stability of small peptides, as present in eHC, during simulated upper gastrointestinal digestion conditions. Next, we aimed to establish the effects of eHC peptides on infant colonic microbiota composition and SCFA production, to elucidate related mechanisms of action that might, if translatable to physiological in vivo conditions, support CMP allergy management and accelerated tolerance development.
2 Material and methods
Hydrolysate digestion in the dynamic gastrointestinal model TIM-1
Two independently manufactured batches of eHC (with 2 years between production) and a preparation of amino acids (AA) were provided by Mead Johnson Nutrition™ (Mead Johnson & Company, LLC, Chicago, IL, USA). These are commercially used in infant formula products and have a comparable AA composition suited for application in infant nutrition for the dietary management of CMP allergy.
The two eHC batches and the AA mixture were individually digested in the TNO Intestinal Model (TIM-1) (TNO, Amsterdam, the Netherlands), a dynamic, multi-compartmental model of the stomach and small intestine. This system was used as previously described with adapted parameters simulating average conditions in the upper gastrointestinal tract of infants up to 6 months of age (Havenaar et al., 2013). Most peptides of interest in this study were only a few amino acids long and would pass through the dialysis filter normally used in TIM-1 (7−8 kDa cut-off). Therefore, instead of the standard filter, dialysis of low molecular weight compounds was performed in a separate step on homogenised material obtained from TIM-1 digestion using a dialysis tube with a cut-off of 500 Da. Freeze-dried retentate was used for peptide analyses.
High resolution liquid chromatography-mass spectrometry (LC-MS) peptide profiling
Peptide analyses of samples digested in the TIM-1 system, as well as after 0 and 6 hours faecal fermentation (see below), were performed on an Orbitrap™ high-resolution mass spectrometer (Thermo Fisher Scientific, Inc, Waltham, MA, USA) equipped with an Accela ultra-high-performance liquid chromatography system. An Acquity HSS T3 1.8 μm high-performance liquid chromatography column (Waters Ltd, Wilmslow, UK) was used with 0.1% formic acid in Milli-Q® (Merck KGaA, Darmstadt, Germany) as mobile phase A and 0.1% formic acid in acetonitrile as mobile phase B. A scan range of m/z 100−2,000 was applied at a resolution of 15.000. Samples were appropriately diluted with a solution containing the internal standards d3-paracetamol and reserpine. Peptide peaks in the chromatograms were integrated using XCalibur™ LCQuan software (Thermo Fisher Scientific, Inc). Peptide peaks were selected based on the analysis of a solution of the eHC.
For further processing of the data, a target table included the 15 most abundant peptides in the eHC solution, defined by their retention time and accurate mass. The target table was used to process the peptide data in all samples. Peak areas were scaled by dividing them by the peak area of the internal standard d3-paracetamol to obtain a measure of peptide abundance (normalised concentration) in the different samples.
The stability of peptides was evaluated by comparing the normalised concentrations after the digestion and fermentation steps to those before digestion and fermentation. If the normalised concentrations were >50% compared to the non-digested material, peptides were considered to be relatively stable or mostly unaffected by upper gastrointestinal digestion or faecal fermentation conditions. For concentrations between 20−50%, peptides were considered partially degraded. Concentrations <20% of the normalised intensity remaining compared to the initial intensity were described as readily degraded.
Infant faecal material collection and preparation for use in faecal fermentations
Following informed consent signed by the parents, fresh faecal material from five healthy, formula-fed infants aged 2.5 to 5.0 months was obtained from children’s day care centres in Houten and Utrecht, The Netherlands. The infants had no prior use of antibiotics and were fed with different formulas including Friso H.A. (FrieslandCampina, Amersfoort, the Netherlands), Nutrilon® (Danone, Paris, France), and Etos standard infant formula (Etos B.V., Zaandam, the Netherlands). In total, 12 diapers were collected, with the number of diapers provided per infant donor varying between one and four. Six diapers originated from three 5 month old infants, two diapers from one 4 month old infant, and four diapers were provided by one 2.5 month old infant.
On collection, diapers were immediately transferred to jars fitted with an Oxoid™ AnaeroGen™ sachet (Thermo Fisher Scientific Inc) ensuring anaerobiosis of faecal material. All subsequent handling was performed inside an anaerobic cabinet. Faecal samples were scraped from each diaper and were pooled, so as to reflect a diversity of samples and be relevant to a broader population. The experimental setup did not take into account proportions of faecal material from individual infants. The pooled samples were mixed with an equal amount of adapted standard ileum efflux medium (SIEM) (without casein) containing 6 mg/ml Bacto™ Peptone (Thermo Fisher Scientific, Inc). After addition of glycerol to a final concentration of 15% (w/v), samples were divided into aliquots, frozen, and stored at −80 °C until further use.
Faecal fermentations
In vitro fermentations were performed in a 96-well plate high throughput format as described previously (Ladirat et al., 2013) using the pooled infant faecal material. Pooling faecal material prior to fermentation is a common approach in exploratory in vitro microbiota studies, as individual donor samples often require donor-specific medium optimisation, and pooling provides a more stable and representative microbial inoculum under standardised fermentation conditions. An adapted version of the SIEM optimised for infant faecal fermentations was used, supplemented with short-chain galactooligosaccharides (1 mg/ml GOS-3 in-house mixture), long-chain fructo-oligosaccharides (1 mg/ml Fructo Oligo Saccharides from chicory, F8052, Sigma-Aldrich®, Merck, St Louis, MO, USA), inulin (0.5 mg/ml inulin from chicory, I2255, Sigma-Aldrich), and mucin (0.5 mg/ml mucin type III, M1778, Sigma-Aldrich). The addition of these prebiotics influences in vitro microbiota composition and metabolism; however, they are required to stabilise the microbial community under these experimental conditions to allow the study of the effects of eHC and AA exposure using a generally relevant infant faecal microbiota composition. Batch pre-fermentations were inoculated from the pooled faecal glycerol stock, suspended in infant-SIEM (without casein) containing 6 mg/ml Bacto Peptone and incubated for 4 h at 37 °C under anaerobic conditions.
The pre-fermentations were diluted in infant SIEM in the individual wells of a 96 well plate with the test materials to a final volume of 1.35 ml. The two eHC batches and AA mixture were tested in a concentration equivalent to protein levels in commercial infant formula and corresponded to final concentrations of 19 mg/ml (1 ×), 9.5 mg/ml (0.5 ×), and 4.75 mg/ml (0.25×). These lower concentrations may be physiologically relevant considering, for example, dilution by digestion fluids. By adding Bacto Peptone, all conditions were normalised to a total of 25.0 mg/ml protein content.
Sampling was performed after 0, 6, 18, and 43 h of anaerobic incubation at 37 °C. Positive controls (fructo-oligosaccharides, maltose, lactose at 10 mg/ml and 5 mg/ml) and negative controls (representing the four Bacto Peptone levels complementary to the test material concentrations: 6.0 mg/ml, 15.5 mg/ml, 20.25 mg/ml and 25.0 mg/ml) were included in the experimental setup. ‘Blank D’ included in descriptions below refers to the addition of 25.0 mg/ml Bacto Peptone without addition of eHC or AA material. For TIM-1 digested samples, addition of an equivalent volume of TIM-1 matrix was included as a negative control. Samples collected at 0 and 6 hours were stored at –20 °C and subjected to peptide and SCFA (acetate, butyrate, propionate) measurements; samples collected at 18 and 43 hours were stored at –80 °C for microbiological analyses. All test conditions were performed in duplicate. The duplicate samples were pooled before further analysis of peptides and SCFA. For microbiota analysis, the samples were pooled after DNA analysis.
Microbiota composition profiling
For microbiota composition analysis after faecal fermentation at 18 and 43 h, samples were analysed by 16S rDNA Illumina sequencing (Illumina, Inc, San Diego, CA, USA). For this purpose, total faecal DNA was extracted from collected samples by phenol/bead beating in combination with the Agowa Mag mini DNA extraction kit (20201, LGC Genomics, Berlin, Germany), as described previously (Crielaard et al., 2011) and quantified using Quant-iT™ PicoGreen™ dsDNA Assay Kit (Invitrogen™, Thermo Fisher Scientific, Inc). Analysis of the microbial composition was performed using Illumina sequencing of the 16S rDNA V4 hypervariable region. Barcoded DNA fragments spanning the Archaeal and Bacterial V4 hypervariable region were amplified from the extracted DNA using the adapted primers 533F (5’-GTG CCA GCA GCC GCG GTA A-3’) and 806R (5’-GGA CTA CVS GGG TAT CTA AT-3’). These amplicons were bi-directionally sequenced using the MiSeq system (Illumina, Inc) as described previously (Caporaso et al., 2012). Each fermentation condition was sequenced in duplicate (technical replicates), and reads were processed separately prior to merging at the ASV count table level.
Sequence pre-processing (filtering), analysis (dereplication, chimera identification, and merging of depaired-end reads), and classification was performed using the DADA2 software package release 1.16 (Callahan et al., 2016) with the bacterial Silva database release 138.1 for taxonomic classification. The classification was performed at the amplicon sequence variant level. Taxonomic count tables were exported into Phyloseq objects (McMurdie and Holmes, 2013). Microbial communities were characterised at the species and genus level. Differentially abundant species were identified for AA and for eHC batch 1 and 2, with Blank D as a reference, using DESeq2 (Love et al., 2014). Differential abundance values are expressed as log2 (fold change) relative to Blank D. Because donor-level biological replication was not available, DESeq2 results were interpreted descriptively based on fold-change directionality rather than statistical inference. Abundance levels were visualised with heatmaps and significance was assessed (adjusted
Short chain fatty acid measurement by LC-MS analysis
SCFA levels were measured in samples of the faecal fermentations by LC-MS after addition of acetate, propionate, and butyrate with stable isotopes as internal standards, to assure technical accuracy, and derivatization with a quarternary ammonium reagent to achieve sufficient retention on an LC column. LC-MS was carried out using a Q Exactive™ system (Thermo Fisher Scientific, Inc) with a Dionex ultra high-performance liquid chromatography pump and autosampler on an Acquity C18 column (100 × 2.1 mm, 1.7 μm (Waters, Ltd.) with a mobile phase gradient from 99.5% water + 0.1% formic acid to 90% acetonitrile + 0.1% formic acid. Data were processed using Quan Browser software (Thermo Fisher Scientific, Inc) for peak integration, calibration, and quantification, using an external calibration curve of each SCFA. Technical variability of the SCFA analysis for each analyte was determined based on the relative standard deviations of two blank fermentation samples (0 and 6 h). The fermentation was performed in triplicate. The relative standard deviations ranged from 1% (butyrate) to 12% (propionate).
3 Results
eHC and AA digestion in TIM-1
The peptide profiles of the two independently produced eHCs were very similar before and after digestion (not shown), indicating low variability across batches as well as reproducibility of the digestion experiments. Qualitative comparison based on visual observation of chromatograms indicated that overall, most peptide peaks were not, or were only partially, degraded after digestion and dialysis (Figure 1A displaying select example peptides 4, 5, 7, corresponding to Peptide Identifiers 455_508, 425_603 and 439_658 in Figure 1B). More detailed analysis revealed some differences in peak levels and patterns, indicating that some peptides were readily or partially digested under TIM-1 conditions (Figure 1C). However, the majority of peptides remained relatively stable.

(A) Liquid chromatography-mass spectrometry chromatograms mass/charge (m/z) 250−500 excerpt of two independent batches of specific eHC before and after TIM-1 digestion and dialysis. Arrows indicate selected example peptides in this m/z range that were relatively unaffected or partially degraded by upper gastrointestinal digestion conditions. (B) Peptide levels of 15 of the most abundant peptides in two independent batches of specific eHC after TIM-1 digestion and dialysis, expressed as percentage of non-digested material, i.e. 100% = levels of peptides before TIM-1 digestion (based on LC-MS peptide analyses). Shading is from red, indicating ≤1% of the original concentration retained (i.e. completely digested) to green, indicating ≥93% of the original concentration retained (i.e. not digested). (C) Relative concentrations of selected peptides after 6 hours of faecal fermentation, expressed as percentage of levels at t = 0, (i.e. 100% = levels of peptides before fermentation, but after digestion) for two concentrations of eHC batch 1. Colour shading in darkest green highlight peptides >125% of the original concentration, dark green depicts peptides 100−125% of the original concentration (deemed mainly stable), green indicates peptides <100% of original concentration (deemed slightly degraded). eHC, extensively hydrolysed casein.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00122
The intensity of 15 of the most abundant peptides was compared before and after TIM-1 digestion and dialysis (Figure 1B). Of those, only three were readily degraded in both eHC batches. The remaining peptides maintained concentrations between 20-100% of the original levels, indicating that they were either only partially degraded or remained relatively stable (Figure 1B). Additionally, the changes were consistent across the batches (Figure 1A). Figure 1A and 1B demonstrate similarity of the two eHC batches as well as reproducibility of the TIM-1 digestion experiment. LC-MS peptide analyses confirmed that the AA mixture did not contain any peptides (data not shown).
In vitro faecal fermentation
With regard to the stability of peptides during faecal fermentations, visual comparison of LC-MS was not possible due to noise in chromatograms (polyethylene glycol signals); however, individual signals of the 15 previously selected peptides were assessed and normalised concentrations were compared before and after faecal fermentation. Figure 1C (6 h only) shows that for the majority of the monitored peptides, levels increased during fermentation with only three of the 15 selected peptides deemed to have been slightly degraded. Relative concentrations were similar after faecal fermentations with different concentrations of eHC, with no specific pattern observed, confirming reproducibility of the faecal fermentation experiments. None of the selected peptides were detected in TIM blank control samples.
Infant faecal microbiota composition following incubation with eHC was assessed by 16S rDNA Illumina sequencing at t = 0, 18, and 43 h. Following 18 h of fermentation, the pH of the incubations with the eHC batches and the individual AA mixture did not decrease substantially (not lower than 5.8) and was comparable to blank conditions (data not shown). As illustrated in Figure 2, effects of the two independently produced batches of eHC on microbial community structure after 18 h of fermentation were similar.
Effects of the two independently manufactured eHC batches and the AA mix (19 mg/ml, 9.5 mg/ml, and 4.75 mg/ml) on the top 25 genera in infant faecal microbiota composition at t = 18 and t = 43 (relative abundance). AA = amino acid; eHC = extensively hydrolysed casein.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00122
The most pronounced observation for effects of eHC exposure was a >2-fold decline in relative abundance of the genus Bacteroides as compared to blank control conditions at t = 18 (Figure 2), which was sustained at 43 h of fermentation (Figure 3). Other genera were affected to a lesser extent by incubation with eHC. These included, for example, an increase in Parasutterella and Fusicatenibacter and an increase in Peptostreptococcus at t = 43 (Figure 3). However, those genera only represented minor parts of the total microbial composition in this analysis (see Figure 3 ‘Relative abundance %’)
Log2 fold changes in the abundance of genera at t = 18 and t = 43 after faecal fermentation with high doses (19 mg/ml) of eHC and AA relative to abundance in Blank D. Values represent log2 (fold change) relative to Blank D. Values > +1 indicate a more than two-fold increase; values < –1 indicate a more than two-fold decrease. The relative presence of each genus in Blank D (Relative abundance percentage, green shading in left columns) provides an indication of the total abundance of each genus in the initial bacterial community and thereby the influence of the relative changes. AA, amino acid; Adj, adjusted; eHC, extensively hydrolysed casein.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00122
With the AA mixture, there were distinct pronounced changes in several genera at t = 18, including Bacteroides, Escherichia/Shigella, Bifidobacterium and Lachnoclostridium, as well as of several less abundant genera (Figure 2 and 3). Overall, effects were even stronger than those observed with the eHC batches.
In vitro fermentations with eHC: effect on Bacteroides
Due to the large proportion of the blank sample being Bacteroides and a consistent effect on Bacteroides abundance in the incubations with eHC and AA, these were investigated further. More detailed comparison of the sequencing results demonstrated a dose-dependent, reproducible effect of the two eHC batches and AA mix on the genus Bacteroides (Figures 2 and 3). The available 16S rDNA sequences from the Illumina analysis were further analysed by oligotyping DNA sequence reads of the genus Bacteroides and tracing the different oligotypes to a species-level resolution (Figure 4).
Log2 fold changes in the abundance of Bacteroides species as an effect of eHC and AA exposure (4.75 mg/ml, 9.5 mg/ml, or 19 mg/ml, respectively) relative to the corresponding blank at t = 18 and 43 hours. Values represent log2(fold change) relative to Blank D. Values > +1 indicate a more than two-fold increase; values < -1 indicate a more than two-fold decrease. The Bacteroides species are sorted from top to bottom based on their general abundance relative to blank (green shading in left columns). Values <0 indicate a two-fold decrease (pink) compared to blank control samples. AA = amino acid; Adj = adjusted; eHC = extensively hydrolysed casein.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00122
No distinct specificity on particular species in the decrease in Bacteroides could be observed. There was a significant decrease in the most abundant Bacteroides uniformis ASV6, but also in several less abundant Bacteroides species. The effects of the AA mixture on Bacteroides species differed only marginally from those of the eHC batches, with a particular decrease in Bacteroides fragilis ASV54.
In vitro fermentations with eHC: effect on SCFA production
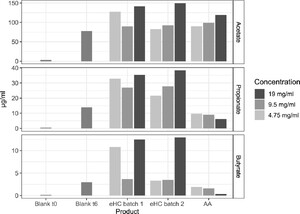
SCFA production from bacterial metabolism was assessed at t = 0 and t = 6 hours of faecal fermentations. As can be seen from Figure 5, the SCFAs acetate, propionate, and butyrate were very low at t = 0 but were produced to some extent during 6 hours of faecal fermentation under control conditions. Production of all three SCFAs increased in most cases (for all at 19 mg/ml) beyond control levels (blank) after 6 hours of faecal fermentations in both eHC batches. Levels of all three SCFAs in incubations with eHC batch 1 at 4.75 mg/ml were higher than in corresponding incubations with eHC batch 2. A dose-response could be observed for all SCFAs upon exposure to eHC batch 2. Faecal fermentations with the AA mix led to increased acetate levels compared to control conditions, but production of propionate and butyrate was lower than control levels at t = 6.
Short chain fatty acid (SCFA) levels in blank faecal fermentations (grey bars; average of triplicates) and after 6 hour incubation with the two eHC batches and AA mix (single measurements). AA = amino acid; eHC = extensively hydrolysed casein.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00122
4 Discussion
The aims of the present study were to investigate the stability of peptides in eHC during simulated upper gastrointestinal digestion conditions and to analyse the effects of eHC on infant gut microbiota composition and SCFA levels, which might be one of the mechanisms supporting infant health in the context of CMP allergy. Although it is known that an estimated 2-4% of infants develop CMP allergy (Tamazouzt et al., 2022; Warren et al., 2022), the exact mechanisms contributing to such development are yet to be elucidated. However, a number of studies indicate a role for gut microbiome alterations in contributing to a change in the balance of T-cell responses (Di Costanzo et al., 2016). Dietary management of CMP allergy is in general directed towards avoidance of cow’s milk and use of a specific hypoallergenic feeding formula (Dı́az et al., 2018). For instance, in a study in infants with CMP allergy, 12 months’ use of an eHC-based formula was associated with acquiring tolerance faster when compared with rice, soy, or AA-based formulas (Berni Canani et al., 2013), and in vitro eHC has been shown to reduce inflammatory cytokine production (Hu et al., 2019).
In the first part of the present study, it was demonstrated that most peptides present in the eHC mixture were partially or fully resistant to upper gastrointestinal digestion in vitro, supporting the hypothesis that specific peptides can be resistant to digestion and therefore can potentially reach the lower GI tract following oral ingestion. These peptides likely possess structural properties making them less accessible to digestive enzymes or do not contain digestive enzyme cleavage sites (Wada and Lonnerdal, 2014). Indeed, in a previous study assessing in vitro digestion of various formulas, all peptides in an eHC formula were also still present after pepsin and pancreatin digestion (Wada and Lonnerdal, 2015). This is also in line with a study by Paparo et al. (2021), in which peptide patterns of protein fractions from eHC formula and digests obtained after static in vitro infant gastroduodenal digestion did not substantially differ from each other and peptide fragments from eHC formula for the greatest part remained stable during digestion (Paparo et al., 2021). The current study extends these insights as in the present work the dynamic TIM-1 digestion model was used as compared to previously applied static digestion protocols, and in vitro digestion was combined with analysis of faecal microbiota composition and SCFA production, representing also colonic fermentation.
Following 6 h faecal fermentation of the eHC batches with an infant gut microbial community, concentrations of the majority of the most abundant peptides increased, most likely as a result of degradation of larger peptides by microbial fermentation. In general, the effects of the two independently manufactured eHC batches demonstrated consistent profiles even if minor differences in composition might be expected due to some raw material variability. This is reflected by findings in other studies, using eHC formulations, showing that batch-to-batch variation in the peptide profiles was low (Lambers et al., 2015; Paparo et al., 2021). However, peptide profiles in infant formula products may differ by brand; therefore, functionalities of a specific casein hydrolysate, and infant formulas based thereon, cannot necessarily be directly translated to other hydrolysates and formulations.
This study also showed that peptides in the eHC preparation could influence infant gut microbial community structure and metabolism during in vitro faecal fermentations. While there is inter-individual variation in infant gut microbiota, in general, Akkermansia, Bifidobacterium, Bacteroides, Citrobacter, Clostridium, Collinsella, Escherichia, Lactobacillus, Streptococcus, and Veillonella spp. (Le Huërou-Luron et al., 2010) are typically present, many of which were found in this study. Gut microbiota of formula-fed infants is, in general, more diverse compared to breast-fed infants and includes a larger abundance of some of these species, including Bacteroides, Clostridium, Escherichia, Streptococcus, and Veillonella spp. (Azad et al., 2013; Ma et al., 2020; Zimmermann and Curtis, 2018). Interestingly, a previous study showed that infants with CMP allergy had a higher diversity of bacterial species compared with healthy controls, which decreased following feeding with an eHC-based formula (Berni Canani et al., 2016).
Differences in gut microbiota composition have also been shown according to protein intake from infant formulas. One study investigating microbiome profiles in infants fed cow’s milk- or soy-based formulas showed Bacteroides to be one of the most abundant bacteria, alongside Bifidobacterium (Brink et al., 2020). A study with a casein-based formula, however, showed lower levels of Bifidobacterium (Hascoët et al., 2011). This is in contrast to studies in whey-protein dominant formula-fed infant faecal samples where Bifidobacterium represented over half of the total microbiota and Bacteroides was almost absent (Hascoët et al., 2011; Wang et al., 2015).
In the current study, one of the most abundant bacteria prior to eHC/AA exposure were Bacteroides. It is of note though that full details of the formulas the infants were fed prior to sample collection were not available as the aim was to collect samples from infants with a diverse background at relatively similar ages so as to represent the general population. It would be of interest for future studies to include only infants fed the same formula with known ingredients, and those from more narrowly defined periods after births.
Following eHC exposure, there was a decline in Bacteroides total abundance. Although Bacteroides are a natural part of a healthy infant’s gut microbiota (Adak and Khan, 2019), some studies have also linked them to disease pathogenesis (Davis-Richardson et al., 2014; Zafar and Saier, 2021). Differences have been found in Bacteroides levels comparing infants with or without non-IgE mediated CMP allergy. However, while in one study Bacteroides relative abundance was much higher in healthy infants (Dı́az et al., 2018), another study found the opposite (Berni Canani et al., 2018), and a further one showed that a reduction in Bacteroides (alongside others) was associated with food allergy development (Ling et al., 2014). Healthy-infant faecal inocula are a widely accepted proxy for early-life microbiota studies in vitro as they provide a basis for investigating general effects of nutritional compounds on infant gut microbiota and metabolism. The observed effects in the current study, including a reduction in the abundance of Bacteroides and an increase in SCFA production, are biologically meaningful and provide a rationale for future follow up research. For instance, further studies are needed to clarify whether changes in specific bacterial genera abundance would impact the risk of developing atopic diseases or influence tolerance development. Findings specific for benefits to infants with CMP allergy will have to be validated in future studies with faecal samples from infants with CMP due to differences in baseline gut microbiota composition compared to healthy infants. It can be speculated that the observed effects of eCH on infant gut microbiota might also be relevant for potential health effects other than CMP allergy. For example, balancing a dominance of Bacteroides in the infant gut community might be a target for modulating successive maturation of infant gut microbiota, which has been described to include a stage that is similar to the potentially dysbiotic Bacteroides 2 enterotype in adults that has been associated with inflammation and loose stools (Beller et al., 2021). In addition, a higher abundance of Bacteroides in the gut microbiota has been described in (children with) coeliac disease (Akobeng et al., 2020), suggesting that eCH might offer potential opportunities for nutritional strategies to reduce disease risk based on reducing Bacteroides abundance.
The impact of the AAs in the faecal fermentation differed largely from that of the eHC batches. While the AA mixture was included to benchmark effects of eHC to a reference sample not containing peptides, the results illustrate that the absence of peptides in an AA mix does not imply that there is no impact on gut microbiota. This might be due to the presence of antibacterial peptides in eHC but not in the AA mixture and, indeed, several casein-derived peptides with antimicrobial properties have been isolated and characterised (Lahov and Regelson, 1996; McCann et al., 2005, 2006). Those mechanisms might be of relevance for the current findings as several peptides originating from eHC were stable during faecal fermentations. In addition, bacterial metabolism might be differentially affected by exposure to single AAs as opposed to specific casein-derived peptides, thereby resulting in specific bacterial cross-feeding mechanisms and shifts in microbial populations. As an example, lactic acid bacteria (e.g. Lactobacillus spp.) have been shown to digest casein and their proteolytic system has been well characterised to include cell-wall associated proteases, peptidases, and peptide transporters enabling the bacteria to convert proteins to peptides and subsequently to AAs (Liu et al., 2010; Tulini et al., 2016). Thus, lactic acid bacteria may provide substrates to indirectly support the growth of saccharolytic bacteria as metabolic interactions between saccharolytic and AA fermenting species have been described (Newton et al., 1998; Tulini et al., 2016). As such, it is notable that the AA mixtures indeed induced pronounced changes in faecal microbiota composition and metabolism.
SCFAs have been linked with a plethora of immune tolerance factors, suppression of inflammatory responses, and prevention of autoimmunity (Kim, 2023). A further finding of this study was an increase in the production of three SCFAs during faecal fermentations – acetate, propionate and butyrate – especially at the highest dose of the two eHC batches. Acetate, the most prominent SCFA in infants aged 3 months (Differding et al., 2020), was increased in faecal fermentations at the highest dose of eHC and AA. This SCFA can contribute to the function of CD8+ T-cells (Qiu et al., 2019) and has an indirect role in the production of other SCFAs through cross-feeding in the gut microbiota (Chia et al., 2021; Rios-Covian et al., 2015). Butyrate and propionate production were particularly increased after fermentation with the highest dose of eHC, but not AA. This is in line with a previous study that showed higher butyrate and propionate levels in formula-fed versus breast-fed infants (Le Huërou-Luron et al., 2010), and with another showing increased butyrate levels after consumption of eHC (Canani et al., 2015). In one study, high faecal butyrate and propionate levels were significantly associated with lower incidence of allergic rhinitis, food allergies, and asthma in young children (McKenzie et al., 2017), with butyrate being shown to be involved in preventing food allergy development (McKenzie et al., 2017). This is likely due to both butyrate and propionate having roles in anti-inflammatory cytokine production and T regulatory cell (Treg) upregulation leading to IL-10 secretion (Adak and Khan, 2019; Donohoe et al., 2011; Hertli and Zimmermann, 2022; Lin et al., 2012; Morrison and Preston, 2016; Smith et al., 2013).
Gut microbial species identified as responsible for SCFA production include Akkermansia muciniphila, Anaerostipes spp., Bacteroides spp., Bifidobacterium spp., Blautia hydrogenotrophica, Clostridium groups IV and XIVa, Eubacterium rectale, Faecalibacterium prausnitzii, and Roseburia spp. (Louis and Flint, 2009; Mirzaei et al., 2022). However, while a number of these bacteria were detected in this study, no major changes in abundance of most of these were observed, except for the pronounced decrease in Bacteroides. This discrepancy might be due to differences in the time points for SCFA analyses (t = 6) and microbiota sequencing (t = 18 and 43) as well as differences between microbiota composition and bacterial metabolic capacity and activity, respectively. The differing timepoints for analyses were chosen in order to monitor acute effects of eHC on SCFA at early timepoints (t = 6) as these result from the direct impact of eHC on bacterial metabolism with SCFAs as end products, while effects on gut microbial community composition rely on bacterial growth and potential cross-feeding resulting in shifts in relative abundance and can therefore be rather determined at later timepoints (t = 18 and 43). A direct impact of the test formulations on bacterial abundance and metabolic activity might have impacted the generation of fermentation products and SCFA levels on the longer term, however these effects were not addressed in this study. To focus on the difference between microbiota composition and bacterial metabolic capacity and activity, bacterial gene expression could be assessed rather than 16S profiling. Such additional analyses would also help unravel the different effects of eHC versus AA on microbial community composition as well as on SCFA production. Effects on other metabolites beyond SCFA, such as tryptophan metabolites and indole derivatives (Fang et al., 2022), may also drive immunological or other physiological mechanisms. These would be relevant to analyse in the context of beneficial effects of eHC on allergy development. While the present exploratory study provides first indications of potential mechanisms mediated by shifts in microbiota composition and SCFA production, further functional analyses at the molecular level, assessment of additional metabolites and complementary research models will be required to elucidate the precise pathways by which eHC-derived peptides modulate infant gut microbial ecology.
Similar to this current study, significantly increased butyrate production was also reported after intake of the same eHC-based infant formula in a clinical trial (Canani et al., 2015). In that study, this was linked to an increase in butyrate-producing taxa such as Roseburia, which was not detected in the current samples. However, this might be due to differences in methodology and background bacterial abundance. Alternatively, subtle changes in microbiota composition in low abundant species might have induced increased cross-feeding of substrate to butyrate producing microorganisms, resulting in higher butyrate levels. These findings are of interest as a study that showed lower levels of butyrate in infants with CMP allergy compared to healthy infants found that use of an eHC formula significantly increased butyrate concentrations (Berni Canani et al., 2018).
The current study is exploratory in nature and needs to be interpreted with caution. For instance, one of the limitations of the experimental setup is that due to pooling of faecal samples for the fermentation experiments inter-individual variation between donors and biological variance could not be assessed since biological replicates cannot be compared. Also, pooling of technical replicates prevented analysis of technical variability in the experiments and greatly limits statistical assessment. Therefore, the results of this study are descriptive only and require further validation in a more extensive experimental setup, e.g. including assessment of inter-individual variations in both metabolite production and microbial composition. Nevertheless, the observed differential effects between eHC and AA formulations, in particular on butyrate and propionate production as well as the observed decrease in Bacteroides spp. provide direction for further investigations.
Under the present study conditions with a limited dataset no conclusions about causal relationships can be drawn. However, the observed associations can serve to generate hypotheses for future research. In particular, further evaluation of potential immunomodulatory and tolerogenic effects of functional peptides in extensive hydrolysates is a promising area. This study underpins the importance of addressing individual variations of responsiveness to functional peptides in the use of eHC for potential risk reduction or alleviation of CMP allergy symptoms. Such future studies will contribute to a better understanding of the full potential of eHC formulas containing functional peptides compared to AA based formulations.
In conclusion, the current findings suggest that not all peptides from eHC are digested in the upper gastrointestinal tract and therefore intact peptides might have the potential to induce changes in gut microbiota contributing to the beneficial effects of eHC-based formula in infants with CMP allergy. Other studies have shown that eHC formula intake in infancy has direct benefits in early life for the management of CMP allergy, but can also lead to reduced prevalence of asthma and eczema even 15 years after dietary intervention at a young age (von Berg et al., 2016). Although the results of the current in vitro experiments are descriptive and exploratory in nature and will have to be validated and confirmed in future in vivo studies, it can be speculated that effects mediated by the gut microbiota as observed in this in vitro study might contribute to beneficial health effects in infants receiving these formulas for management of CMP allergy in early and later life.
Corresponding author; e-mail: bart.keijser@tno.nl
Acknowledgements
The study was financially supported by Mead Johnson Nutrition. Writing support was provided by Dr Eleanor Roberts of Beeline Science Communications, Ltd, funded by the study sponsor.
Authors’ contribution
All authors have read and agreed to the published version of the manuscript. Conceptualization: GG, EAFvT, IBP, BJFK; methodology: GG, EAFvT, IBP, AMTO, BJFK; analyses: IBP, AMTO, MvdW, BJFK; writing-original draft preparation: GG, RS, IBP, BJFK; writing-review and editing: GG, IBP, AMTO, MvdW, RS, EAFvT, BJFK; funding acquisition: GG, EAFvT.
Conflict of interest
GG, RS and EAFvT were employees of Mead Johnson Nutrition/Reckitt at the time of study execution. This study was financially supported by Mead Johnson Nutrition.
References
Abdalkareem Jasim, S., Jade Catalan Opulencia, M., Alexis Ramı́rez-Coronel, A., Kamal Abdelbasset, W., Hasan Abed, M., Markov, A., Raheem Lateef Al-Awsi, G., Azamatovich Shamsiev, J., Thaeer Hammid, A., Nader Shalaby, M., Karampoor, S. and Mirzaei, R., 2022. The emerging role of microbiota-derived short-chain fatty acids in immunometabolism. International Immunopharmacology 110: 108983. https://doi.org/10.1016/j.intimp.2022.108983
Aitoro, R., Simeoli, R., Amoroso, A., Paparo, L., Nocerino, R., Pirozzi, C., di Costanzo, M., Meli, R., De Caro, C., Picariello, G., Mamone, G., Calignano, A., Nagler, C.R. and Berni Canani, R., 2017. Extensively hydrolyzed casein formula alone or with L. rhamnosus GG reduces β-lactoglobulin sensitization in mice. Pediatric Allergy and Immunology 28: 230-237. https://doi.org/10.1111/pai.12687
Azad, M.B., Konya, T., Maughan, H., Guttman, D.S., Field, C.J., Chari, R.S., Sears, M.R., Becker, A.B., Scott, J.A. and Kozyrskyj, A.L., 2013. Gut microbiota of healthy Canadian infants: profiles by mode of delivery and infant diet at 4 months. Canadian Medical Association Journal 185: 385-394. https://doi.org/10.1503/cmaj.121189
Beller, L., Deboutte, W., Falony, G., Vieira-Silva, S., Tito, R.Y., Valles-Colomer, M., Rymenans, L., Jansen, D., van Espen, L., Papadaki, M.I., Shi, C., Yinda, C.K., Zeller, M., Faust, K., van Ranst, M., Raes, J. and Matthijnssens, J., 2021. Successional stages in infant gut microbiota maturation. mBio 12(6): e0185721. https://doi.org/10.1128/mBio.01857-21
Berni Canani, R., De Filippis, F., Nocerino, R., Paparo, L., Di Scala, C., Cosenza, L., Della Gatta, G., Calignano, A., De Caro, C., Laiola, M., Gilbert, J.A. and Ercolini, D., 2018. Gut microbiota composition and butyrate production in children affected by non-IgE-mediated cow’s milk allergy. Science Reports 8: 12500. https://doi.org/10.1038/s41598-018-30428-3
Berni Canani, R., Nocerino, R., Terrin, G., Frediani, T., Lucarelli, S., Cosenza, L., Passariello, A., Leone, L., Granata, V., Di Costanzo, M., Pezzella, V. and Troncone, R., 2013. Formula selection for management of children with cow’s milk allergy influences the rate of acquisition of tolerance: a prospective multicenter study. Journal of Pediatrics 163: 771-777. https://doi.org/10.1016/j.jpeds.2013.03.008
Berni Canani, R., Sangwan, N., Stefka, A.T., Nocerino, R., Paparo, L., Aitoro, R., Calignano, A., Khan, A.A., Gilbert, J.A. and Nagler, C.R., 2016. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME Journal 10: 742-750. https://doi.org/10.1038/ismej.2015.151
Bognanni, A., Fiocchi, A., Arasi, S., Chu, D.K., Ansotegui, I., Assa’ad, A.H., Bahna, S.L., Berni Canani, R., Bozzola, M., Dahdah, L., Dupont, C., Dziechciarz, P., Ebisawa, M., Firmino, R.T., Chu, A., Galli, E., Horvath, A., Kamenwa, R., Lack, G., Li, H., Martelli, A., Nowak-Węgrzyn, A., Papadopoulos, N.G., Pawankar, R., Roldan, Y., Said, M., Sánchez-Borges, M., Shamir, R., Spergel, J.M., Szajewska, H., Terracciano, L., Vandenplas, Y., Venter, C., Waffenschmidt, S., Waserman, S., Warner, A., Wong, G.W.K., Schünemann, H.J. and Brozek, J.L., 2024. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) guideline update – XII – Recommendations on milk formula supplements with and without probiotics for infants and toddlers with CMA. World Allergy Organization Journal 17: 100888. https://doi.org/10.1016/j.waojou.2024.100888
Booijink, C.C., El-Aidy, S., Rajilić-Stojanović, M., Heilig, H.G., Troost, F.J., Smidt, H., Kleerebezem, M., De Vos, W.M. and Zoetendal, E.G., 2010. High temporal and inter-individual variation detected in the human ileal microbiota. Environmental Microbiology 12: 3213-3227. https://doi.org/10.1111/j.1462-2920.2010.02294.x
Brink, L.R., Mercer, K.E., Piccolo, B.D., Chintapalli, S.V., Elolimy, A., Bowlin, A.K., Matazel, K.S., Pack, L., Adams, S.H., Shankar, K., Badger, T.M., Andres, A. and Yeruva, L., 2020. Neonatal diet alters fecal microbiota and metabolome profiles at different ages in infants fed breast milk or formula. American Journal of Clinical Nutrition 111: 1190-1202. https://doi.org/10.1093/ajcn/nqaa076
Canani, R.B., Sangwan, N., Stefka, A.T., Nocerino, R., Paparo, L., Aitoro, R., Calignano, A., Khan, A.A., Gilbert, J.A. and Nagler, C.R., 2015. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME Journal 10: 742-750. https://doi.org/10.1038/ismej.2015.151
Chia, L.W., Mank, M., Blijenberg, B., Bongers, R.S., van Limpt, K., Wopereis, H., Tims, S., Stahl, B., Belzer, C. and Knol, J., 2021. Cross-feeding between Bifidobacterium infantis and Anaerostipes caccae on lactose and human milk oligosaccharides. Beneficial Microbes 12: 69-83. https://doi.org/10.3920/bm2020.0005
Crielaard, W., Zaura, E., Schuller, A.A., Huse, S.M., Montijn, R.C. and Keijser, B.J., 2011. Exploring the oral microbiota of children at various developmental stages of their dentition in the relation to their oral health. BMC Medical Genomics 4: 22. https://doi.org/10.1186/1755-8794-4-22
Davis-Richardson, A.G., Ardissone, A.N., Dias, R., Simell, V., Leonard, M.T., Kemppainen, K.M., Drew, J.C., Schatz, D., Atkinson, M.A., Kolaczkowski, B., Ilonen, J., Knip, M., Toppari, J., Nurminen, N., Hyoty, H., Veijola, R., Simell, T., Mykkanen, J., Simell, O. and Triplett, E.W., 2014. Bacteroides dorei dominates gut microbiome prior to autoimmunity in Finnish children at high risk for type 1 diabetes. Frontiers in Microbiology 5: 678. https://doi.org/10.3389/fmicb.2014.00678
Dı́az, M., Guadamuro, L., Espinosa-Martos, I., Mancabelli, L., Jiménez, S., Molinos-Norniella, C., Pérez-Solis, D., Milani, C., Rodrı́guez, J.M., Ventura, M., Bousoño, C., Gueimonde, M., Margolles, A., Dı́az, J.J. and Delgado, S., 2018. Microbiota and derived parameters in fecal samples of infants with non-IgE cow’s milk protein allergy under a restricted diet. Nutrients 10: 1481. https://doi.org/10.3390/nu10101481
Differding, M.K., Benjamin-Neelon, S.E., Hoyo, C., Østbye, T. and Mueller, N.T., 2020. Timing of complementary feeding is associated with gut microbiota diversity and composition and short chain fatty acid concentrations over the first year of life. BMC Microbiology 20: 56. https://doi.org/10.1186/s12866-020-01723-9
Fleischer, D.M., Chan, E.S., Venter, C., Spergel, J.M., Abrams, E.M., Stukus, D., Groetch, M., Shaker, M. and Greenhawt, M., 2021. A consensus approach to the primary prevention of food allergy through nutrition: guidance from the American Academy of Allergy, Asthma, and Immunology; American College of Allergy, Asthma, and Immunology; and the Canadian Society for Allergy and Clinical Immunology. Journal of Allergy and Clinical Immunology: Practice 9: 22-43.e24. https://doi.org/10.1016/j.jaip.2020.11.002
Goldberg, M.R., Mor, H., Magid Neriya, D., Magzal, F., Muller, E., Appel, M.Y., Nachshon, L., Borenstein, E., Tamir, S., Louzoun, Y., Youngster, I., Elizur, A. and Koren, O., 2020. Microbial signature in IgE-mediated food allergies. Genome Medicine 12: 92. https://doi.org/10.1186/s13073-020-00789-4
Hascoët, J.M., Hubert, C., Rochat, F., Legagneur, H., Gaga, S., Emady-Azar, S. and Steenhout, P.G., 2011. Effect of formula composition on the development of infant gut microbiota. Journal of Pediatric Gastroenterology and Nutrition 52: 756-762. https://doi.org/10.1097/MPG.0b013e3182105850
Havenaar, R., Anneveld, B., Hanff, L.M., de Wildt, S.N., de Koning, B.A., Mooij, M.G., Lelieveld, J.P. and Minekus, M., 2013. In vitro gastrointestinal model (TIM) with predictive power, even for infants and children? International Journal of Pharmaceutics 457: 327-332. https://doi.org/10.1016/j.ijpharm.2013.07.053
Ladirat, S.E., Schols, H.A., Nauta, A., Schoterman, M.H., Keijser, B.J., Montijn, R.C., Gruppen, H. and Schuren, F.H., 2013. High-throughput analysis of the impact of antibiotics on the human intestinal microbiota composition. Journal of Microbiological Methods 92: 387-397. https://doi.org/10.1016/j.mimet.2012.12.011
Le Huërou-Luron, I., Blat, S. and Boudry, G., 2010. Breast- v. formula-feeding: impacts on the digestive tract and immediate and long-term health effects. Nutrition Research Reviews 23: 23-36. https://doi.org/10.1017/s0954422410000065
Lin, H.V., Frassetto, A., Kowalik, E.J., Jr., Nawrocki, A.R., Lu, M.M., Kosinski, J.R., Hubert, J.A., Szeto, D., Yao, X., Forrest, G. and Marsh, D.J., 2012. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 7: e35240. https://doi.org/10.1371/journal.pone.0035240
Maslowski, K.M., Vieira, A.T., Ng, A., Kranich, J., Sierro, F., Yu, D., Schilter, H.C., Rolph, M.S., Mackay, F., Artis, D., Xavier, R.J., Teixeira, M.M. and Mackay, C.R., 2009. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461: 1282-1286. https://doi.org/10.1038/nature08530
Mirzaei, R., Dehkhodaie, E., Bouzari, B., Rahimi, M., Gholestani, A., Hosseini-Fard, S.R., Keyvani, H., Teimoori, A. and Karampoor, S., 2022. Dual role of microbiota-derived short-chain fatty acids on host and pathogen. Biomedicine and Pharmacotherapy 145: 112352. https://doi.org/10.1016/j.biopha.2021.112352
Newton, D.F., Cummings, J.H., Macfarlane, S. and Macfarlane, G.T., 1998. Growth of a human intestinal Desulfovibrio desulfuricans in continuous cultures containing defined populations of saccharolytic and amino acid fermenting bacteria. Journal of Applied Microbiology 85: 372-380. https://doi.org/10.1046/j.1365-2672.1998.00522.x
Paparo, L., Picariello, G., Bruno, C., Pisapia, L., Canale, V., Sarracino, A., Nocerino, R., Carucci, L., Cosenza, L., Cozzolino, T. and Berni Canani, R., 2021. Tolerogenic effect elicited by protein fraction derived from different formulas for dietary treatment of cow’s milk allergy in human cells. Frontiers in Immunology 11: 604075. https://doi.org/10.3389/fimmu.2020.604075
Qiu, J., Villa, M., Sanin, D.E., Buck, M.D., O’Sullivan, D., Ching, R., Matsushita, M., Grzes, K.M., Winkler, F., Chang, C.-H., Curtis, J.D., Kyle, R.L., Van Teijlingen Bakker, N., Corrado, M., Haessler, F., Alfei, F., Edwards-Hicks, J., Maggi, L.B., Zehn, D., Egawa, T., Bengsch, B., Klein Geltink, R.I., Jenuwein, T., Pearce, E.J. and Pearce, E.L., 2019. Acetate promotes T cell effector function during glucose restriction. Cell Reports 27: 2063-2074. https://doi.org/10.1016/j.celrep.2019.04.022
Seekatz, A.M., Schnizlein, M.K., Koenigsknecht, M.J., Baker, J.R., Hasler, W.L., Bleske, B.E., Young, V.B. and Sun, D., 2019. Spatial and temporal analysis of the stomach and small-intestinal microbiota in fasted healthy humans. mSphere 4. https://doi.org/10.1128/mSphere.00126-19
Thompson-Chagoyan, O.C., Fallani, M., Maldonado, J., Vieites, J.M., Khanna, S., Edwards, C., Doré, J. and Gil, A., 2011. Faecal microbiota and short-chain fatty acid levels in faeces from infants with cow’s milk protein allergy. International Archives of Allergy and Immunology 156: 325-332. https://doi.org/10.1159/000323893
Tulini, F.L., Hymery, N., Haertle, T., Le Blay, G. and De Martinis, E.C., 2016. Screening for antimicrobial and proteolytic activities of lactic acid bacteria isolated from cow, buffalo and goat milk and cheeses marketed in the southeast region of Brazil. Journal of Dairy Research 83: 115-124. https://doi.org/10.1017/S0022029915000606
Von Berg, A., Filipiak-Pittroff, B., Schulz, H., Hoffmann, U., Link, E., Sussmann, M., Schnappinger, M., Bruske, I., Standl, M., Kramer, U., Hoffmann, B., Heinrich, J., Bauer, C.P., Koletzko, S. and Berdel, D., 2016. Allergic manifestation 15 years after early intervention with hydrolyzed formulas–the GINI Study. Allergy 71: 210-219. https://doi.org/10.1111/all.12790
Wang, M., Li, M., Wu, S., Lebrilla, C.B., Chapkin, R.S., Ivanov, I. and Donovan, S.M., 2015. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. Journal of Pediatric Gastroenterology and Nutrition 60: 825-833. https://doi.org/10.1097/mpg.0000000000000752
Wickham, H., 2016. ggplot2: elegant graphics for data analysis. Springer-Verlag, New York, NY, USA. Available at: https://ggplot2.tidyverse.org.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}