Abstract
Obesity is a growing public health concern in the Middle East and North Africa (MENA) region, yet limited research has explored how gut microbiota varies between Arab populations. This study compared the gut microbiota composition and diversity of Emirati and Lebanese adults with obesity and assessed the role of age and nationality in shaping microbial variation. A total of 43 Emirati and 30 Lebanese individuals with obesity (body mass index (BMI) ≥35 kg/m2) were recruited. Participants provided anthropometric and biochemical data, dietary records, and stool samples for 16S rRNA sequencing. The analysis revealed significantly higher BMI, weight, and fat mass in Emirati participants, while Lebanese individuals reported higher fibre intake. Taxonomic profiling showed higher relative abundances of Pseudomonadota, Mycoplasmatota, Cyanobacteriota, and Lentisphaerota in the Lebanese group, whereas Bacteroidota was more abundant among Emiratis. Lebanese participants also exhibited significantly greater microbial alpha-diversity. Beta-diversity analysis confirmed clear distinctions in microbial community structure between the two groups. Linear Discriminant Analysis Effect Size (LefSe) (LDA score >10log2) and regression models (
1 Introduction
Obesity has emerged as one of the most pressing global health challenges of the 21st century. According to the World Health Organization (2024), more than one billion people – approximately one in eight worldwide – are currently living with obesity (WHO, 2024). The World Obesity Federation (2025) projects that by 2030, adult obesity prevalence will have increased by over 115% compared with 2010, reaching roughly 1.13 billion individuals globally. Furthermore, long-term forecasts from the Global Burden of Disease (GBD) BMI Collaborators (2025) suggest that by 2050, around 60% of adults (nearly 3.8 billion people) and one-third of children and adolescents (≈ 746 million) will be overweight or obese worldwide (IHME, 2025). These trends are particularly concerning in the Middle East and North Africa, where obesity rates are among the fastest rising. Obesity rates have been steadily rising in Gulf countries, with prevalence increasing from 6.5% in 1975 to 20% in 2016 (Ritchie, 2017; WHO, 2016). According to the United Arab Emirates (UAE)’s 2017–2018 National Health Survey, obesity affected 27.8% of the UAE population, while in Lebanon, the prevalence was 30.8% as reported by the World Health Organization in 2016 (WHO, 2016). Without effective prevention and policy interventions, obesity-related diseases – including type 2 diabetes, cardiovascular disease, and other metabolic complications – are expected to increase substantially, especially in low- and middle-income settings (GBD 2021 Adult BMI Collaborators, 2025; IHME, 2025; WHO, 2024; World Obesity Federation, 2025).
Genetic predisposition, lifestyle and environmental factors contribute to obesity, whilst emerging evidence highlights the gut microbiota as a key regulator of in particular, environmental and diet-related effects on metabolism and energy homeostasis (Patrone et al., 2016; Turnbaugh et al., 2006). The gut microbiota comprises not only bacterial phyla but also fungi, archaea, viruses, and protozoa, with its bacterial component being primarily composed of Bacillota, Bacteroidota, Actinomycetota, Pseudomonadota, Fusobacteriota, and Verrucomicrobiota (Oren and Garrity, 2021), with Bacillota and Bacteroidota accounting for nearly 90% of microbial communities (Arumugam et al., 2011; Rinninella et al., 2019).
An imbalance in gut microbiota, commonly referred to as dysbiosis, remains a debated concept, as microbial composition varies considerably among individuals, making it difficult to establish universal criteria for distinguishing a dysbiotic state from a healthy profile. Nevertheless, variations in gut microbiota have been linked to several metabolic and inflammatory disorders, including obesity, diabetes, and allergies (Gupta et al., 2017). Early animal studies suggested a strong relationship between the Bacillota/Bacteroidota ratio and obesity (Turnbaugh et al., 2006), but subsequent findings in humans have been inconsistent (Magne et al., 2020; Sze and Schloss, 2016). Beyond this ratio, specific bacterial phyla and genera have also been associated with obesity (OB) (Crovesy et al., 2020). Individuals with obesity, insulin resistance (IR), and type 2 diabetes mellitus (T2DM) frequently exhibit altered gut microbiota compared to lean counterparts, characterised by reduced microbial richness and diversity as well as lower levels of butyrate-producing bacteria, changes that have been linked to metabolic impairments (Jardon et al., 2022). Increasingly, researchers emphasise that definitions of dysbiosis should account not only for taxonomic shifts but also for functional consequences on host metabolism and health.
The composition of gut microbiota varies among individuals and across populations, influenced by lifestyle, genetics, age, diet, medication use, geography, ethnicity, and environment (Crovesy et al., 2020; Deschasaux et al., 2018; Gupta et al., 2017; Jardon et al., 2022; Pinart et al., 2021). Advances in high-throughput sequencing technology have enhanced our understanding of population-specific differences in gut microbiota (Yasir et al., 2015).
While the Lebanese and Emirati populations share some environmental and cultural similarities, variations in dietary patterns, lifestyle habits, and genetic backgrounds may shape distinct gut microbiota profiles. These differences raise concerns about the universality of microbiome-based therapies, emphasising the need for geographically tailored, community-specific approaches to microbiome research (Gupta et al., 2017).
Although multiple studies have explored the relationship between gut microbiota and obesity in different populations (Escobar et al., 2014; Jain et al., 2018), comparative research in Middle Eastern populations remains scarce (Yasir et al., 2015). Furthermore, data on ethnicity-driven gut microbiota differences in Arab populations remain inconsistent (Brooks et al., 2018).
This study aims to bridge this research gap by analysing gut microbiota composition and diversity in Lebanese and Emirati individuals with OB using high throughput 16S ribosomal RNA (rRNA) gene amplicon sequencing and assessing whether age or other factors contribute to these observed variations. Identifying population-specific microbiota signatures associated with OB will provide valuable insights for developing targeted interventions to manage and prevent obesity-related complications in the Middle East, where research remains limited.
2 Materials and methods
Study design
We conducted a cross-sectional design among Lebanese and Emirati subjects with OB living in the UAE and Lebanon, respectively. The present study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Middle East Institute of Health University Hospital (MEIH-UH) in Lebanon (AL-04-02/2018-IRB/MEIH) and by the Research Ethics Committee of the Ministry of Health and Prevention of UAE (MOHP/DXB-REC-52/2018), the Dubai Health Care Regulatory Research Ethics Committee (DHCR-REC), and the Zayed University Ethical Committee Board (ZU19_51_F) in UAE.
Study population
Recruitment took place from October 2019 until April 2021. Sample size was determined by following standard calculations based on normal distributions. The current study utilised data from a mother study designed to investigate gut microbiota profiles in Lebanese and Emirati participants, both with OB and of normal weight, as well as changes following bariatric surgery. The present analysis was restricted to obese participants from these larger cross-sectional studies that included both obese and normal-weight adults from Lebanon and the UAE. The current manuscript focuses solely on the obese subgroup to assess ethnicity-related microbial differences within obesity. In the mother study, the primary outcome was the change in the Bacillota /Bacteriodota ratio. Sample size calculation was based on expected changes in this ratio before and after bariatric surgery, referencing findings from Damms-Machado et al. (2015), which reported a significant increase in the faecal Bacillota /Bacteriodota ratio from 5.9 (SD 2.1) to 10.4 (SD 1.4) three months post-surgery. Based on these values, a minimum of two participants was required to achieve 80% power at a two-sided 5% significance level (Ali Ahmad et al., 2025). To account for protocol deviations and anticipated reductions in effect size (∼30%), researchers conservatively multiplied the required sample by a factor of 15. This adjustment yielded a final target of 30 participants per group. Accordingly, we aimed to recruit 30 Emirati and 30 Lebanese adults with OB, including both sexes, residing in the UAE and Lebanon, respectively.
Inclusion and exclusion criteria
Participants were eligible if they were adults between 18 and 60 years of age, had a body mass index (BMI) ≥ 35 kg/m2, and had maintained stable body weight (no fluctuation greater than 5%) during the three months prior to recruitment. Eligibility was restricted to Emirati and Lebanese nationals with both parents of the same nationality. Additional requirements included absence of antibiotic use in the previous three months and willingness to complete a 3-day 24-hour food diary (two weekdays and one weekend) to assess habitual energy and fibre intake. For Emirati participants, dietary intake was analysed using the Middle Eastern food composition database available in the Nutritics software, whereas for Lebanese participants, food reference tables from Understanding Nutrition (Whitney and Rolfes, 2002) were used. All participants provided written informed consent before enrolment. Eligibility was assessed through a structured screening questionnaire (Supplementary Materials and methods S1), which captured information on general health, weight history, alcohol intake, physical activity, smoking, medication use, and supplement intake (including probiotics, fibre, and prebiotics).
Participants were excluded if they had experienced weight loss ≥5% within the last three months, were pregnant, reported excessive alcohol consumption (>1/2 ounce/day for females; >1 ounce/day for males) (Snetselaar et al., 2021), or were unwilling or unable to provide informed consent.
Recruitment
Lebanese participants with obesity were recruited from the weight-loss program at the Middle East Institute of Health University Hospital (MEIH-UH), while Emirati participants with obesity were recruited from hospitals in Dubai and Sharjah. Recruitment took place between October 2019 and April 2021.
Anthropometric measurements
Eligible subjects were asked to arrive at the testing site after a 12 h fast and were asked to avoid alcohol and caffeine consumption, as well as any unusual strenuous exercise 24 h prior to the screening. While subjects wore light clothes and no shoes, height and weight were measured using the ‘SECA 700’ balance and BMI was calculated as the ratio of weight (kg) to height (m) squared (kg/m2). Waist circumference (WC) (cm) was measured to the nearest 0.1 cm using a flexible, non-stretchable measuring tape at the umbilicus level midway between the lower ribs and the upper iliac crest while standing (Alberti et al., 2006). WC was defined as normal or ‘at risk’ according to the International Diabetes Federation (IDF) using Europids cutoff values for normal of <94 cm for males and <80 cm for females (Alberti et al., 2006). Waist-to-height ratio (WtHR) was calculated as the ratio of the waist circumference divided by height (both in cm). A cut-off of 0.5 was used to define abdominal obesity, whereby a WtHR <0.5 was considered normal (Gibson and Ashwell, 2020).
The percentage of body fat (PBF) was measured using a bioelectrical impedance analyzer 200 (BC-420 MA, Tanita Corporation, Tokyo, Japan) for Emiratis and an InBody composition analyzer 720 (InBody Co., Ltd., Seoul, Korea) for Lebanese participants, according to the manufacturer’s instructions. Cutoffs of ≥25% and ≥35% were used to define elevated PBF among males and females, respectively (Nuttall, 2015).
Biochemical analyses
After a 12 h fast, on-site, venous blood samples were collected by a registered nurse for the analyses of fasting blood glucose (FBG, mg/dl), total cholesterol (TC, mg/dl), low-density lipoprotein cholesterol (LDL-C, mg/dl), high-density lipoprotein cholesterol (HDL-C, mg/dl), and triglyceride (TG, mg/dl) that were analysed via an enzymatic colorimetric method using the Roche/Hitachi Cobas® 6000 analyzer (Roche, Basel, Switzerland) and insulin via the Enzyme-Linked Immunosorbent Assay (ELISA) principle using the Elecsys insulin kit (Roche) for Lebanese participants. As for Emirates, the former were measured via the portable Lux Meter Blood Test (Biochemical Systems International, S.p.A; Arezzo, Italy) following the manufacturer’s instructions. As for insulin: one 5 ml venous blood sample was drawn into an ethylenediaminetetraacetic acid (EDTA) tube and analysed by using ELISA kits (Diametra Millipore, St. Louis, MO, USA).
Insulin resistance was calculated using the Homeostatic Model Assessment for Insulin Resistance (HOMA-IR), defined as (fasting immunoreactive insulin (μU/ml) × fasting plasma glucose (mg/dL))/ 405 (Vaccaro et al., 2004). HOMA-IR values exceeding 1.8 for Emirati participants (Esteghamati et al., 2009) and 2.32 for Lebanese participants (Chedid et al., 2009) were considered indicative of insulin resistance.
Stool collection
Stool was collected using a faecal specimen collector along with the Zymo DNA/RNA Shield™ Faecal Collection tube which was pre-filled with DNA/RNA Shield™ (9 ml) (Zymo Research Corp., Irvine, CA, USA). Subjects were asked to follow the instructions by the developer of the tubes, where two samples were collected from each participant, one of which was analysed, and the second was held for accurate verification. Collection tubes had a spoon attached to the cap for collecting 1 g of faeces. The nucleic acids (DNA & RNA) in the samples were preserved by DNA/RNA Shield™ at room temperature (DNA > 1 year, RNA up to 1 month).
DNA isolation and sequencing of the V3–V4 region of the 16S rRNA gene
DNA isolation and sequencing of the barcoded amplicons of the V3–V4 region of the 16S rRNA gene were performed according to the Illumina protocols (Illumina, Eindhoven, the Netherlands) which involved PCR amplification using region-specific primers, followed by indexing, purification, and paired-end library preparation (Surono et al., 2022). Sequencing was conducted on the Illumina MiSeq platform (San Diego, CA, USA) using barcodes and the paired end 2 × 300 bp protocol. Raw reads were processed and demultiplexed using (QIIME 2, version 2023.2) (Bolyen et al., 2019), and taxonomic classification was performed using the SILVA 16S rRNA gene reference database, version 132 (Quast et al., 2013; Yilmaz et al., 2014).
Diversity analyses
The alpha-diversity metrics used in this study included Faith’s phylogenetic diversity (PD) (Faith, 1992), observed features (DeSantis et al., 2006), Chao1 index (Chao, 1984), Pielou’s evenness (Pielou, 1966) and Shannon’s indices (Shannon and Weaver, 1949). These indices were calculated using QIIME 2 and visualised in RStudio (version 4.2.1) using the phyloseq and ggplot2 packages (Wickham, 2016). Rarefaction curves were also generated using the ggplot2 package, with rarefication depth set at 3800. Beta-diversity was assessed using Bray–Curtis dissimilarity (Bray and Curtis, 1957), Jaccard similarity (Jaccard, 1908), unweighted UniFrac, and weighted UniFrac (Lozupone and Knight, 2005) and visualised through Principal Coordinate Analysis (PCoA) plots using the vegan and phyloseq packages in R.
Statistical analyses
Statistical analyses were performed using the Statistical Package for the Social Sciences software version 20 (SPSS Inc. Chicago, IL, USA) and software package R (4.2.1) (R Core Team, http://www.R-project.org) in RStudio. The normality of data using SPSS was tested using the skewness and kurtosis. Normally distributed data is expressed as mean (M) ± standard deviation (SD), and skewed data as median (Mdn) and interquartile range (IQR). Depending on the data distribution, differences between the groups were assessed using an independent samples t-test for normally distributed continuous variables or a nonparametric test using the Mann–Whitney U-test for skewed continuous variables and chi-square for categorical values. The value of P < 0.05 was used to denote statistical significance.
Comparisons of relative abundances at the phylum and genus levels were performed using the non-parametric Kruskal–Wallis H-test corrected with the Benjamini–Hochberg false discovery rate (FDR) for multiple comparisons by using the software package R in RStudio. Although only two groups were compared, the Kruskal–Wallis test was retained for the microbiota data to maintain consistency with the LEfSe workflow and because its distribution-free properties are well suited to highly skewed genus-level abundance data. In contrast, Mann–Whitney U-tests were used for clinical variables, as this is the standard non-parametric method for two-group comparisons. Spearman’s rank correlation coefficient was used to evaluate associations between microbial taxa and clinical variables. In addition, linear regression models were applied to examine the relationship between microbial composition and nationality, with age assessed as a potential confounder. Both unadjusted and age-adjusted models were reported. For each genus-level analysis, the relative abundance of the bacterial genus was used as the dependent variable in separate linear regression models, and nationality (Lebanese vs. Emirati) served as the main independent variable (coded 1 and 0, respectively). Relative abundances were analysed as proportional values without transformation (e.g. no log, arcsine-square-root, or CLR transformation), as exploratory inspection indicated that model residuals met acceptable assumptions for linear regression. Zero values were retained as observed without imputation. Age was added as a covariate in adjusted models to assess confounding, applying a ≥10% change-in-estimate criterion for identifying meaningful confounding effects (Skelly et al., 2012). Model assumptions – including linearity, homoscedasticity, multicollinearity, and normality of residuals – were evaluated using residual plots and diagnostic checks. No additional covariates met criteria for inclusion.
Taxonomic composition was summarised using stacked bar plots at the phylum and genus levels, illustrating the relative abundance of dominant microbial taxa across study groups.
The statistical significance of beta-diversity between the two groups with OB (Emirati vs. Lebanese) was determined using Permutational Multivariate Analysis of Variance (PERMANOVA) using the software package R in RStudio.
Linear discriminant analysis (LDA) effect size (LEfSe) was done using the R software in RStudio to explore microbial taxa with differential abundance between Lebanese and Emirati individuals with OB. Genera enriched in each group with an LDA score >10log2 at a value of
3 Results
Participant characteristics
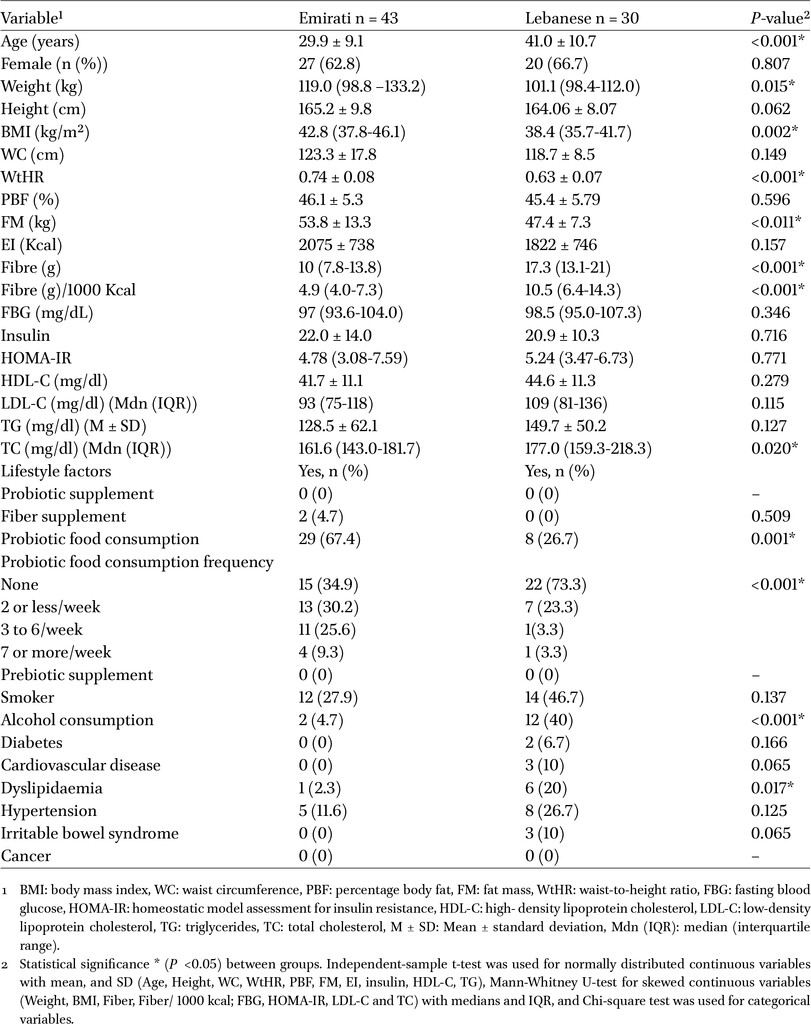
The participant characteristics are presented in Table 1. A total of 43 Emirati and 30 Lebanese individuals with OB (BMI ≥35 kg/m2) were enrolled in the study. The difference in sample sizes between the two groups was mainly due to recruitment challenges in Lebanon. While the broader study initially aimed to enrol participants undergoing bariatric surgery from both countries, enrolment in Lebanon was hindered by financial constraints and disruptions caused by the COVID-19 pandemic.

Demographic, anthropometric, and biochemical parameters between Lebanese and Emirati individuals with obesity
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Most of the subjects were females including 62.8 and 66.7% of the Emirati and Lebanese participants, respectively (P = 0.807). Anthropometric measurements showed that weight (
Alpha- and beta-diversity
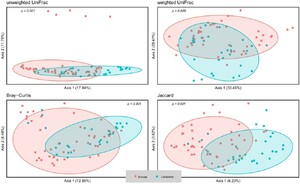
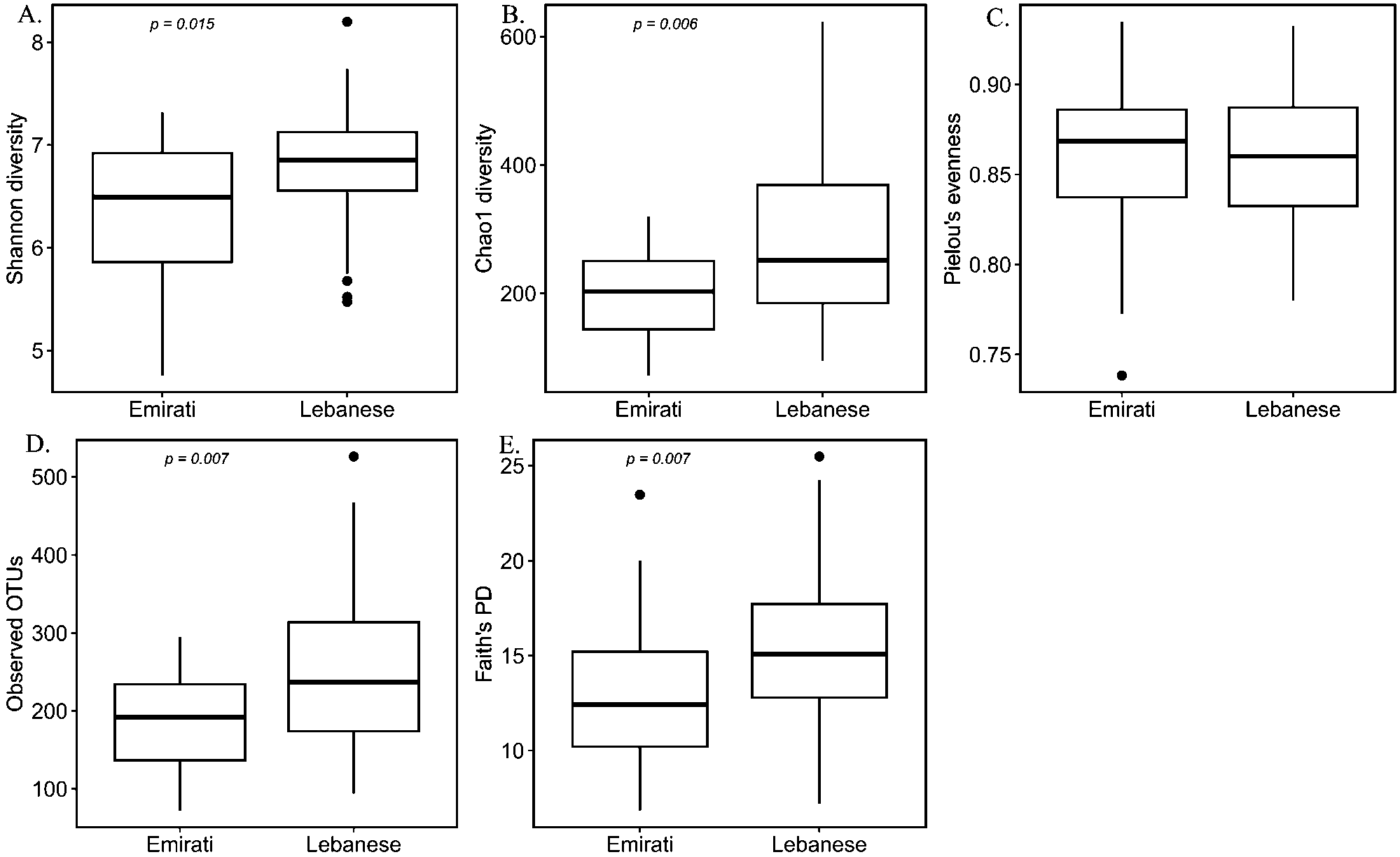
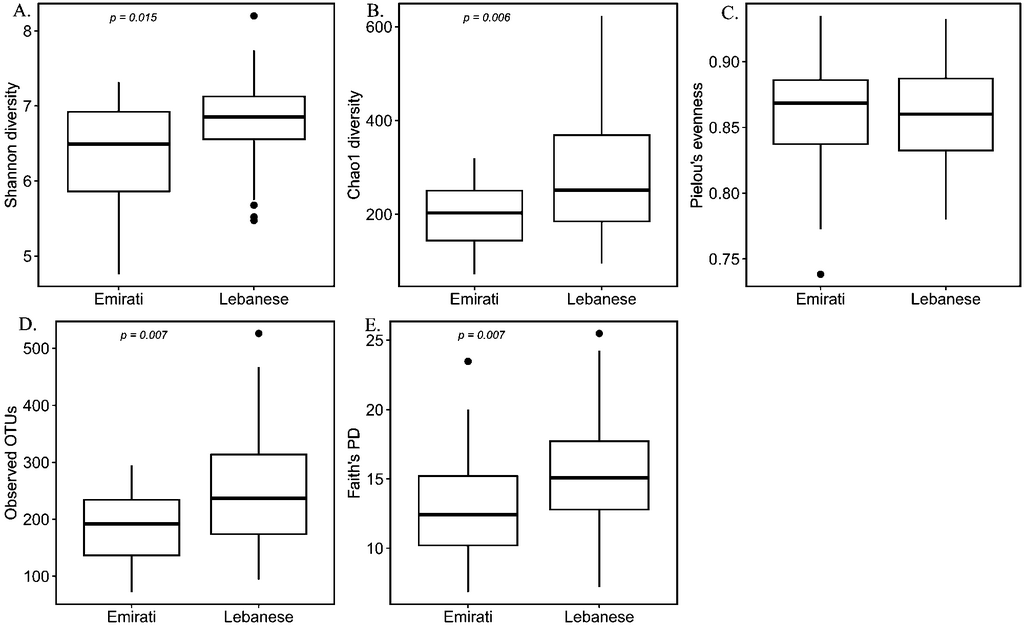
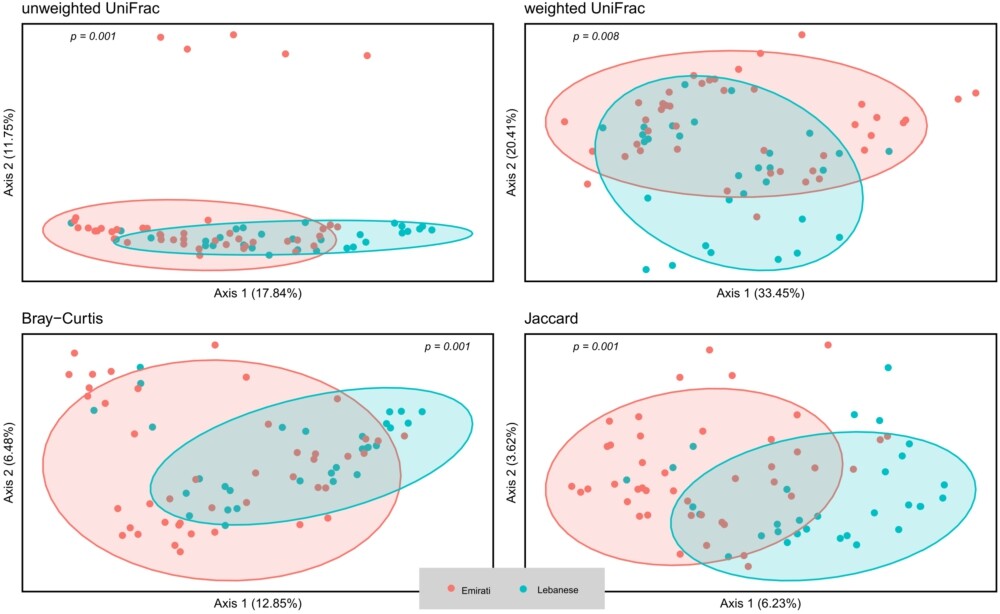
According to the alpha-diversity metrics (Figure 1), Lebanese adults with OB showed significantly higher diversity (Shannon index, P = 0.015), coinciding with the higher observed operational taxonomic unit (OTU) (P = 0.007), higher Chao1 richness (p = 0.006), and higher Faith’s phylogenetic diversity (PD) (P = 0.007) compared to the Emirati group, but no statistically significant difference in evenness (Pielou’s evenness, P = 0.964). Microbial composition as reflected by the beta-diversity measures including unweighted UniFrac, weighted UniFrac, Bray–Curtis dissimilarity, and Jaccard similarity showed a marked separation between the gut microbiota community of Lebanese and Emirati with OB. Pairwise PERMANOVA analysis indicated a significant difference in beta-diversity between the two groups (Figure 2); (P = 0.001; P = 0.008; P = 0.001; P = 0.001, respectively).
Alpha-diversity of Lebanese and Emirati individuals with obesity. Alpha-diversity boxplot showing Shannon diversity (A), Chao1 diversity (B), Pielou’s evenness (C), Observed operational taxonomic unit (OTU)(D), and Faith’s Phylogenetic Diversity (PD) (E) of Lebanese and Emirati individuals with obesity. For each sample, the alpha-diversity indices are shown on the y-axis, and the groups (Emirati or Lebanese) are on the x-axis. Boxes denote the interquartile range (IQR) between the first and third quartiles, and the horizontal line defines the median. Whiskers represent the smallest (ymin) and largest (ymax) values. Outliers are indicated by black circles. Kruskal Wallis test was performed. A P-value <0.05 denotes a significant difference between Lebanese and Emirati individuals with obesity.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Beta-diversity of Lebanese and Emirati individuals with obesity. Principal coordinate analysis (PCoA) plot of unweighted Unifrac, weighted Unifrac, Bray-Curtis dissimilarity, and Jaccard similarity index between Lebanese (blue) and Emirati individuals (red) with OB. A P-value <0.05 denotes a significant difference between the two groups.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Taxonomic analysis
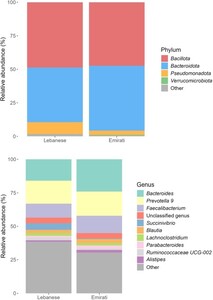
Taxonomic profiling of the gut microbiota was conducted and visualised using stacked bar plots. At the phylum level (Figure 3A), the microbial communities of both Lebanese and Emirati participants with obesity were dominated by Bacillota and Bacteroidota, which together accounted for nearly 90% of the total bacterial abundance. In the Lebanese group, Bacillota represented approximately 49% of the community, followed by Bacteroidota (41%), while Pseudomonadota (about 9%) and Verrucomicrobiota (<1%) were detected in lower proportions. Similarly, in the Emirati group, Bacillota (47%) and Bacteroidota (49%) were the most abundant phyla, with smaller contributions from Pseudomonadota (3%) and other phyla such as Actinomycetota, Mycoplasmatota, Cyanobacteriota, and Lentisphaerota, each present at very low levels.
Stacked bar plots. (Top) Stacked bar plots of phylum- level and (Bottom) at genus level composition detected in datasets of Lebanese vs. Emirati individuals with obesity. Relative abundances (percentages) of the phyla are displayed.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
At the genus level (Figure 3B), the gut microbiota composition included a range of taxa commonly reported in obese populations. The most represented genera included Bacteroides, Prevotella, Faecalibacterium, Ruminococcus, Lachnospira, Blautia, and Alloprevotella, reflecting their contribution to carbohydrate fermentation and short-chain fatty acid production. Additional taxa such as Collinsella, Paraprevotella, Dialister, Eubacterium ruminantium group, Intestinimonas, and Caproiciproducens were also present, alongside multiple low-abundance genera distributed across individuals.
It is noteworthy that several taxa were completely absent in one group but present in the other at phylum (Figure 4A) and genus level (data not shown). For example, Allisonella, Howardella, Intestinimonas, and Caproiciproducens were detected in the Lebanese cohort but absent in Emiratis, whereas other genera such as Bacteroides and Parabacteroides were enriched in Emiratis but nearly absent in Lebanese participants.
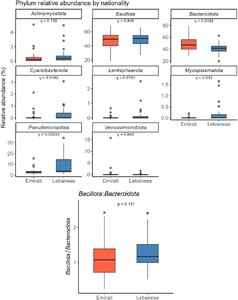
Comparative relative abundance of selected bacterial phyla and Bacillota/Bacteroidota ratio in Lebanese and Emirate participants with obesity. (Top panels) Boxplots showing relative abundance (%) of selected phyla, stratified by nationality with q-values. (Bottom panel) Boxplot of Bacillota/Bacteroidota ratio stratified by nationality. Differences between groups were assessed using the Kruskal-Wallis test followed by false discovery rate (FDR). Adjusted q-values are indicated where applicable. A q-value of ≤ 0.05 was considered statistically significant, as determined by the Kruskal-Wallis test with FDR correction.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Taken together, these findings indicate that the gut microbiota in both Lebanese and Emirati individuals with obesity was shaped by a limited number of dominant phyla and genera, complemented by a broad range of low-abundance taxa that added to inter-individual variability within each population.
Differences in gut microbial genera between Lebanese and Emirati individuals with obesity
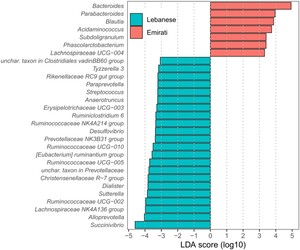
Based on the relative abundances of the gut microbiota in the Lebanese vs. Emirati individuals with OB, LEfSe identified several genera that differed significantly between Lebanese and Emirati individuals, with only taxa exceeding the LDA threshold (>10 log2) and P < 0.05 reported.
In the Emirati group, Bacteroides, Parabacteroides (both of the Bacteroidota phylum), Blautia, Acidaminococcus, Subdoligranulum, Phascolarctobacterium, and Lachnospiraceae UCG-004 (all Bacillota phylum) were enriched. In contrast, Lebanese participants showed higher abundances of 22 bacterial genera: Unchar.taxon Clostridiales vadinBB60 group 1, Tyzzerella, Streptococcus, Anaerotruncus, Erysipelotrichaceae UCG-003, Ruminiclostridium 6, Ruminococcaceae NK4A214 group, Ruminococcaceae UCG-010, Eubacterium ruminantium group, Ruminococcaceae UCG-005, Christensenellaceae R-7 group, Dialister, Ruminococcaceae UCG-002, Lachnospiraceae NK4A 136 group (all Bacillota phylum); Rikenellaceae RC9 gut group, Paraprrevotella, Prevotellaceae NK3B31 group, Unchar.taxon in Prevotellaceae, and Alloprevotella (all Bacteroidota phylum); Desulfovibrio, Sutterella, and Succinivibrio (all Pseudomonadota phylum) (Figure 5).
Linear discriminant analysis effect size (LEfSe) plot. The genera enriched in Lebanese and Emirati individuals with OB are coloured by blue and red, respectively. Histogram of the LDA scores computed for features differentially abundant among Lebanese and Emirati individuals with obesity. Only class meetings with an LDA significant threshold >10log2 and
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
According to the Kruskal-Wallis analysis with Benjamini–Hochberg correction at the genus level, as seen in Figure 6, there is an overlap with some of the results from the LEfSe. For the Lebanese group with OB, in accordance with the LEfSe results, there is an increased relative abundance of 14 out of the 22 microbial taxa, which are Alloprevotella (q <0.001), Lachnospiraceae NK4A136 group (q <0.001), Ruminococcaceae UCG-005 (
Relative abundance of selected gut bacterial genera by nationality. Boxplots depict the relative abundance (%) of selected bacterial genera in Lebanese and Emirati participants with obesity. Group comparisons were performed using the Kruskal–Wallis test, with false discovery rate (FDR) correction applied to account for multiple testing. Adjusted q-values are displayed for each genus.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Spearman correlation between gut microbiota and age in Lebanese and Emirati individuals with obesity
There was on average more than 10-year difference between Lebanese and Emirati participants (Table 1). Spearman correlation analysis with FDR correction (Figure 7 and Table 2) identified several bacteria that were significantly positively associated with age (q < 0.01). The most pronounced correlation was observed for Ruminococcaceae NK4A214 (rho = 0.397, q = 0.0014), followed by Prevotellaceae D_5 uncultured (rho = 0.386, q = 0.002), Ruminococcaceae UCG-005 (rho = 0.364, q = 0.004), and Anaerotruncus (rho = 0.359, q = 0.005). Additional significant associations were found for Alloprevotella (rho = 0.358, q = 0.005), Ruminococcaceae UCG-002 (rho = 0.358, q = 0.005), Clostridiales vadin BB60 group (rho = 0.354, q = 0.005), and Ruminococcaceae UCG-010 (rho = 0.347, q = 0.006). Among all clinical and demographic variables assessed, age was the only factor significantly correlated with specific bacterial genera after FDR correction (Figure 7). Collectively, these findings indicate that age plays a substantial role in shaping gut microbiota composition, particularly influencing members of the Ruminococcaceae family and other bacteria such as Prevotellaceae D_5 uncultured and Anaerotruncus. The uniformity of positive associations suggests a gradual, age-related enrichment of these bacteria. None of the other clinical parameters were significantly correlated with any of the taxa.
Spearman correlation analysis between gut microbiota and clinical variables in Lebanese and Emirati individuals with obesity. In the heatmap, blue squares indicate significant positive correlations, and red squares indicate significant negative correlations. * Significant correlation at q <0.05 (FDR-correction). Only age showed significant correlations with gut microbial taxa after FDR correction. Abbreviations: BMI: Body mass index; WC: waist circumference; WtHR: waist to height ratio, PBF: percent body fat; FM: fat mass; HOMA-IR: homeostatic model assessment for insulin resistance LDL-C: low density lipoprotein cholesterol, HDL-C: high density lipoprotein cholesterol; TG: triglyceride.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Correlation analysis between gut microbiota and age in Lebanese and Emirati individuals with obesity
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
Linear regression analysis
The relationship between microbial composition and nationality was subsequently examined, comparing Lebanese and Emirati individuals with OB, while accounting for age as a potential confounder. Linear regression models were used (Table 3) to assess these associations, with both unadjusted and adjusted models reported. A change-in-estimate criterion with a cut-off of 10% was used to identify the confounder (Skelly et al., 2012).
Linear regression analysis between gut microbiota and age among Lebanese and Emirati individuals with obesity.1
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00111
In the unadjusted model, Lebanese individuals had a significantly higher relative abundance of Alloprevotella compared to Emiratis (β = 0.0109,
For Prevotellaceae
In Clostridiales vadin BB60, a strong association was observed in the unadjusted model (β = 0.0012,
For Anaerotruncus, Lebanese nationality was significantly associated with bacterial abundance in the unadjusted model (β = 0.0015, P = 0.036). However, after adjusting for age, the effect size declined by 35.3% (β = 0.00097) and was no longer significant (P = 0.229), further indicating confounding by age.
The unadjusted model for Ruminococcaceae
For Ruminococcaceae
No significant association was found for Ruminococcaceae
Finally, for Ruminococcaceae
Overall, the regression analyses revealed a consistent pattern across taxa: several nationality–microbiota associations observed in the unadjusted models were attenuated after controlling for age, indicating that age acted as a meaningful confounder for many genera. However, a subset of taxa – including Prevotellaceae D_5, Clostridiales vadinBB60, and Ruminococcaceae UCG-010 – remained significantly associated with nationality even after adjustment, suggesting a robust nationality-linked microbial signature. In addition, age itself showed positive associations with several genera, reinforcing its influential role in shaping gut microbial composition within this cohort.
4 Discussion
This study contributes to the growing body of research exploring how geography, culture, and ethnicity shape gut microbiota and its relationship with obesity. By comparing obese individuals from two Arab nationalities – Lebanese and Emirati residing in their respective countries, we highlight distinct microbial patterns potentially driven by regional dietary and lifestyle habits. Existing studies, such as comparisons between French and Saudi individuals, have shown that dietary and cultural factors can significantly influence microbial composition (Yasir et al., 2015). Despite global interest in ethnic differences in microbiota composition, data from Middle Eastern populations remain limited. Our findings emphasise the need for more inclusive studies in this region to better understand how cultural and environmental factors influence metabolic health and to inform tailored intervention strategies.
The comparison of Lebanese and Emirati individuals with OB provides a meaningful framework to explore how cultural norms shape gut microbiota and potentially affect metabolic outcomes. The predominance of female participants 62.8 to 66.7% reflects a broader regional trend of higher obesity prevalence among cis-women in the Middle East and North Africa (MENA), a phenomenon attributed to sociocultural perceptions associating larger body size with beauty and affluence (Mallat et al., 2016; Naja et al., 2011; Sibai et al., 2003). These findings suggest that public health strategies targeting obesity in MENA countries should consider the cultural contexts influencing body image and dietary behaviours, alongside biological factors such as the gut microbiota.
Despite overall dietary similarities, Lebanese participants reported significantly higher fibre intake. While no direct correlations between fibre intake and microbial taxa were observed after FDR correction, fibre’s known role in modulating gut microbiota may partly explain the distinct microbial profiles observed. In contrast, Emirati individuals exhibited higher anthropometric measures – including weight, BMI, WtHR, and fat mass – although these were not significantly associated with specific bacterial genera in our dataset. These differences align with previous studies linking higher fibre intake to greater gut microbial diversity and improved metabolic health (Nasreddine et al., 2012, 2014). Comparable patterns have been observed in other cross-cultural comparisons, such as between French and Saudi populations, where differences in fibre consumption and traditional versus Westernised diets influenced microbial composition (Yasir et al., 2015). Another important demographic difference in our cohort was age, with Lebanese participants being on average more than 10 years older than Emiratis (Table 1). Since age is a well-recognised determinant of gut microbiota composition, this imbalance needed to be considered when interpreting our findings.
Our findings revealed that Lebanese adults with OB exhibit significantly greater gut microbial diversity compared to their Emirati counterparts. This is supported by multiple alpha-diversity indices, including the Shannon index, observed features, Chao1 richness, and Faith’s PD. Greater microbial diversity may indicate a more complex and versatile ecosystem, potentially conferring resilience to metabolic diseases, as various studies have associated increased gut microbiota diversity with improved metabolic profiles in both humans and mice (Astbury et al., 2020; Cotillard et al., 2013; Le Chatelier et al., 2013; Meyer and Bennett, 2016). Additionally, although a meta-analysis found that obesity is linked to alterations in gut microbial composition, including changes in richness and diversity, significant discrepancies were observed across different cohorts (Chanda and De, 2024). For instance, high BMI has been associated with decreased gut microbial diversity, and certain taxa, such as Christensenellaceae, have been linked to BMI variations across ethnicities (Balakrishnan et al., 2021).
In terms of beta-diversity, PCoA was used to visualise differences between the Lebanese and Emirati populations, and these were confirmed as statistically significant by pairwise PERMANOVA analyses, indicating marked differentiation in microbial community composition. This suggests that nationality – and by extension, environmental and dietary exposures – may play a critical role in shaping gut microbiota profiles (Parizadeh and Arrieta, 2023; Yasir et al., 2015).
Taxonomic analysis of gut microbiota revealed both shared and distinct features. While both groups exhibited comparable proportions of the dominant phyla Bacillota and Bacteroidota – consistent with established gut microbiota compositions (Chen et al., 2021; Zhang et al., 2023) – notable differences emerged in less abundant but potentially influential taxa. The Lebanese group displayed significantly higher levels of Pseudomonadota (9% vs 3% in Emiratis), Mycoplasmatota (q = 0.003), Cyanobacteriota (q = 0.009), and Lentisphaerota (q = 0.049), whereas the Emirati group exhibited a greater abundance of Bacteroidota (q = 0.009). Elevated levels of Mycoplasmatota in the Lebanese group may reflect the influence of Western dietary components, such as high fat and sugar intake, which have been associated with increased obesity risk (Konstantinidis et al., 2020; Napoli et al., 2024). Although the Bacillota/Bacteroidota ratio has historically been linked to obesity-related microbiota changes (Boulange et al., 2016; Compare et al., 2016; John et al., 2018; Turnbaugh et al., 2008), our results underscore the limitations of using this ratio as a universal marker of obesity. In our study, Lebanese individuals with OB exhibited a higher, but not significant, Bacillota/Bacteroidota ratio than obese Emiratis, despite the latter presenting with higher overall obesity levels. This discrepancy aligns with recent research questioning the reliability of the Bacillota/Bacteroidota ratio across ethnically and geographically diverse populations (Magne et al., 2020). While some studies have observed elevated Bacillota/Bacteroidota ratios in obese individuals (Koliada et al., 2017), others report inconsistent or population-specific patterns (Kim et al., 2019). These findings highlight the need to move beyond broad phylum-level comparisons and instead investigate specific microbial genera and their functional contributions to host metabolism (Bauer et al., 2016).
Geographic, ethnic, and cultural factors are well-documented determinants of gut microbiota composition (Yatsunenko et al., 2012). As previous studies show, Americans, Japanese, Koreans, and Chinese individuals each harbour distinct microbial profiles, with varying proportions of Bacillota, Actinomycetota, and Bacteroidota, respectively (Brooks et al., 2018). At the genus level, variations are similarly pronounced, with Bacteroides enriched in Chinese populations, Prevotella in Indians, and Ruminococcus in the Dutch (Brooks et al., 2018). Our findings reflect these ethnically stratified patterns. Emiratis – characterised by higher BMI – had increased levels of Bacteroides and Parabacteroides, which are associated with high-fat, high-sugar diets and enhanced energy harvest. Conversely, Lebanese participants had higher abundances of Alloprevotella and Paraprevotella, linked to high-fibre intake and improved metabolic profiles (Christensen et al., 2018). These results bolster the existing evidence linking Bacteroides-dominant microbiota to increased adiposity, while leaner individuals often harbour Prevotella-rich communities (Lee et al., 2024). Supporting this, studies comparing French and Saudi populations (Yasir et al., 2015), as well as investigations among Emiratis (Ahmad et al., 2023) and Lebanese (Abou-Samra et al., 2025) have highlighted gut microbiota differences shaped by diet, lifestyle, and cultural practices.
LEfSe analysis further identified taxa that discriminate between the two populations. Emirati individuals showed enrichment in Bacteroides, while Lebanese participants had higher levels of Lachnospiraceae. Both genera have been previously implicated in metabolic functions related to energy harvest, lipid metabolism, and fat storage (Wu et al., 2024; Yang et al., 2022). However, interpretation of these differences must consider the significant age gap between the groups. Spearman correlation analysis confirmed that age significantly correlated with several taxa, and linear regression models showed that some between-group differences – particularly at the genus level – were attenuated or lost after adjusting for age. This underscores the role of age as a confounding factor in microbiota studies, as also reported in previous research (Chen et al., 2021; De la Cuesta-Zuluaga et al., 2019; Odamaki et al., 2016).
Age-related changes in the gut microbiome have been extensively documented across the human lifespan. Research spanning from infancy to old age has identified key transitions in microbial composition – particularly during early development and later life stages (Odamaki et al., 2016). Some studies report minimal age-related differences (Takagi et al., 2019), while others identify significant associations between age and microbial composition (De la Cuesta-Zuluaga et al., 2019). These inconsistencies indicate that age can act as a confounder in microbiome studies and must be carefully considered. In our cohort, although age explained part of the between-group variation, several taxa – including Prevotellaceae D_5, Clostridiales vadinBB60, and Ruminococcaceae UCG-010 – remained significantly associated with Lebanese nationality after adjustment. This suggests that nationality-related dietary and environmental factors continue to influence gut microbiota composition independently of age, consistent with evidence that distinct microbial signatures persist across populations even when controlling for demographic variables (Brooks et al., 2018; Yatsunenko et al., 2012).
Each of these taxa has known links to health-promoting microbial functions. Prevotellaceae D_5 is associated with fibre-rich diets and improved glycaemic control (Kovatcheva-Datchary et al., 2015); Clostridiales vadinBB60 has been found in metabolically healthy individuals and centenarians and is implicated in lipid metabolism and anti-inflammatory effects (Zhang et al., 2019). Ruminococcaceae UCG-010 plays a role in fibre degradation and short-chain fatty acid production, which support gut health and are linked to GLP-1 release, satiety, and protection against type 2 diabetes (Gryaznova et al., 2022; Mancabelli et al., 2017; Tims et al., 2013; Tsai et al., 2020; Wei et al., 2018). The persistence of these associations despite age adjustment highlights the key role of cultural and environmental influences tied to nationality in shaping the gut microbiota.
Although ethnicity was not found to have a significant correlation with microbiota composition in a Middle Eastern population, lifestyle and environmental influences exhibited a greater degree of homogeneity compared to the ethnic, sociocultural, economic, and dietary diversity observed within the United States (Brooks et al., 2018). Importantly, our study controlled for methodological consistency – using identical DNA extraction and sequencing protocols – yet still observed significant differences between groups. This reinforces the notion that factors such as diet (Asnicar et al., 2021), genetics (Goodrich et al., 2014), physical activity, and medication use (Heiman and Greenway, 2016) contribute meaningfully to microbiota variation.
Strengths and limitations
This study provides novel comparative insights into how regional and ethnic factors may shape gut microbiota composition in the context of obesity, focusing on two Arab populations living in the MENA region. The use of standardised microbiota sequencing protocols across sites minimised methodological variability, while comprehensive data collection – including anthropometric, biochemical, and selected dietary variables – facilitated robust analysis. The application of linear regression models adjusted for age, along with rigorous participant selection and harmonised methodologies, further strengthened the reliability and interpretability of the findings. Nonetheless, several limitations should be considered. The cross-sectional design limits the ability to infer causality, suggesting that there must be longitudinal research to examine microbiota dynamics over time. The relatively small sample size may constrain the generalisability of the results. This study also focused exclusively on obese participants from a larger cross-sectional dataset that also included normal-weight adults. As such, comparisons with normal-weight controls were beyond the scope of the current analysis but will be explored in a separate study using the same parent cohort. Another notable limitation is the significant age difference between the Lebanese and Emirati participants, which could independently influence gut microbiota composition. While we addressed this through correlation and regression analyses, which confirmed age as a confounding factor, residual confounding cannot be fully excluded. Although age differences between the 2 samples were accounted for, other relevant factors – such as physical activity, medication use, psychosocial stress, socioeconomic status, and genetic background – were not assessed and may have influenced gut microbiota composition. Additionally, dietary assessment was limited to fibre and probiotic intake, omitting other potentially significant dietary components (e.g. fat and protein intake). Variability in laboratory assays between countries may have introduced inconsistencies in biochemical measurements, and the lack of functional microbiota profiling restricted insights into microbial metabolic potential. Additionally, the use of different bioelectric impedance analysis devices across study sites (Tanita in the UAE and InBody in Lebanon) may introduce inter-device variability and measurement bias, which could affect the comparability of body composition data between the two populations.
5 Conclusions
This study highlights significant gut microbiota differences between Lebanese and Emirati adults with OB, shaped by cultural, dietary, and geographic factors. Emiratis exhibited a higher abundance of Bacteroides and Parabacteroides, bacteria commonly linked to high-fat, low-fibre diets and greater energy harvest. In contrast, Lebanese individuals showed greater microbial diversity and elevated levels of Alloprevotella, Paraprevotella, and Mycoplasmatota, reflecting higher fibre intake. Furthermore, this study reveals that dietary patterns rooted in cultural and geographic context strongly influence the composition of gut microbiota. The study also identified age as key confounder, which significantly influenced the associations between nationality and microbial profiles. These findings highlight the importance of incorporating both age – and ethnicity-specific microbiome signatures when designing targeted obesity interventions. Notably, microbial differences such as higher abundances of Prevotellaceae D_5, Clostridiales vadinBB60, and Ruminococcaceae UCG-010 in Lebanese individuals remained significant after adjusting for age, emphasising the potential for culturally driven, microbiota-informed strategies. Ultimately, the gut microbiota may serve as both a biomarker of lifestyle and a modifiable factor in the management of obesity and its metabolic consequences within Arab populations living in the MENA region. Future research should adopt longitudinal designs to elucidate the causal mechanisms linking gut microbiota to obesity and metabolic health, particularly in response to dietary and lifestyle interventions. Including non-obese control groups and diverse Arab populations will enhance generalisability and provide insights into ethnicity-specific microbial variations. Integrating functional metagenomics, metabolomic profiling, and detailed dietary, lifestyle, and genetic data will deepen our understanding of microbial activity and its metabolic impact. Additionally, culturally tailored dietary interventions and investigations into age-related microbial shifts across life stages are warranted to inform effective, personalised strategies for weight management in the MENA region.
Corresponding authors; e-mail: mirey.karavetian@utoronto.ca; koen.venema@wur.nl
These authors share first authorship
Acknowledgements
We thank the research assistants at the Middle East Institute of Health University Hospital in Lebanon for their assistance in data collection and collaboration and the research assistants and students from the College of Natural and Health Sciences department at Zayed University in Dubai campus for their assistance in data collection, Dr. Sulaiman al Habib Hospital (Dubai, UAE), Al Qassimi Hospital (Sharjah, UAE), and the Research Institute for Medical and Health Sciences (Sharjah, UAE) for their collaboration and involvement in the study.
Authors’ contribution
Conceptualization, M.A.S., M.A.A., M.K. and K.V.; methodology, M.A.S., M.A.A., M.K. and K.V.; software, M.A.S., M.A.A., and K.V.; validation, M.K. and K.V.; formal analysis, M.A.S. and M.A.A.; investigation, M.A.S. and M.A.A.; resources, M.K., K.V. and C.A.M.; data curation, M.A.S., M.A.A., M.K. and K.V.; writing – original draft preparation, M.A.S. and M.A.A.; writing – review and editing, M.K., K.V., E.B and C.A.M.; visualization, M.A.S., M.A.A., M.K. and K.V.; supervision, M.K., K.V., and E.B.; project administration, M.K. and K.V.; funding acquisition, C.A.M. and K.V. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
KV is editor in chief of Beneficial Microbes, but has not been involved in the review and decision process of this article. The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Funding
This research was funded by Zayed University, Dubai, UAE (grant number R19054) and was supported by the Dutch Province of Limburg with a grant to the Centre for Healthy Eating and Food Innovation (HEFI) of Maastricht University Campus Venlo (Grant number HEFI-1).
References
Abou-Samra, M., Venema, K., Ayoub Moubareck, C. and Karavetian, M., 2025. Gut microbiota composition of lean and obese Lebanese individuals. Beneficial Microbes. https://doi.org/10.1163/18762891-bja00062
Arumugam, M., Raes, J., Pelletier, E., Le Paslier, D., Yamada, T., Mende, D.R., Fernandes, G.R., Tap, J., Bruls, T., Batto, J.M., Bertalan, M., Borruel, N., Casellas, F., Fernandez, L., Gautier, L., Hansen, T., Hattori, M., Hayashi, T., Kleerebezem, M., Kurokawa, K., Leclerc, M., Levenez, F., Manichanh, C., Nielsen, H.B., Nielsen, T., Pons, N., Poulain, J., Qin, J., Sicheritz-Ponten, T., Tims, S., Torrents, D., Ugarte, E., Zoetendal, E.G., Wang, J., Guarner, F., Pedersen, O., de Vos, W.M., Brunak, S., Doré, J., Antolı́n, M., Artiguenave, F., Blottiere, H.M., Almeida, M., Brechot, C., Cara, C., Chervaux, C., Cultrone, A., Delorme, C., Denariaz, G., Dervyn, R., Foerstner, K.U., Friss, C., van de Guchte, M., Guedon, E., Haimet, F., Huber, W., van Hylckama-Vlieg, J., Jamet, A., Juste, C., Kaci, G., Knol, J., Lakhdari, O., Layec, S., Le Roux, K., Maguin, E., Mérieux, A., Melo Minardi, R., M’rini, C., Muller, J., Oozeer, R., Parkhill, J., Renault, P., Rescigno, M., Sanchez, N., Sunagawa, S., Torrejon, A., Turner, K., Vandemeulebrouck, G., Varela, E., Winogradsky, Y., Zeller, G., Weissenbach, J., Ehrlich, S.D. and Bork, P., MetaHIT Consortium, 2011. Enterotypes of the human gut microbiome. Nature 473: 174-180. https://doi.org/10.1038/nature09944
Astbury, S., Atallah, E., Vijay, A., Aithal, G.P., Grove, J.I. and Valdes, A.M., 2020. Lower gut microbiome diversity and higher abundance of proinflammatory genus Collinsella are associated with biopsy-proven nonalcoholic steatohepatitis. Gut Microbes 11(3): 569-580. https://doi.org/10.1080/19490976.2019.1681861
Bolyen, E., Rideout, J.R., Dillon, M.R., Bokulich, N.A., Abnet, C.C., Al-Ghalith, G.A., Alexander, H., Alm, E.J., Arumugam, M., Asnicar, F., Bai, Y., Bisanz, J.E., Bittinger, K., Brejnrod, A., Brislawn, C.J., Brown, C.T., Callahan, B.J., Caraballo-Rodriguez, A.M., Chase, J., Cope, E.K., Da Silva, R., Diener, C., Dorrestein, P.C., Douglas, G.M., Durall, D.M., Duvallet, C., Edwardson, C.F., Ernst, M., Estaki, M., Fouquier, J., Gauglitz, J.M., Gibbons, S.M., Gibson, D.L., Gonzalez, A., Gorlick, K., Guo, J., Hillmann, B., Holmes, S., Holste, H., Huttenhower, C., Huttley, G.A., Janssen, S., Jarmusch, A.K., Jiang, L., Kaehler, B.D., Kang, K.B., Keefe, C.R., Keim, P., Kelley, S.T., Knights, D., Koester, I., Kosciolek, T., Kreps, J., Langille, M.G.I., Lee, J., Ley, R., Liu, Y.X., Loftfield, E., Lozupone, C., Maher, M., Marotz, C., Martin, B.D., McDonald, D., McIver, L.J., Melnik, A.V., Metcalf, J.L., Morgan, S.C., Morton, J.T., Naimey, A.T., Navas-Molina, J.A., Nothias, L.F., Orchanian, S.B., Pearson, T., Peoples, S.L., Petras, D., Preuss, M.L., Pruesse, E., Rasmussen, L.B., Rivers, A., Robeson, M.S. 2nd, Rosenthal, P., Segata, N., Shaffer, M., Shiffer, A., Sinha, R., Song, S.J., Spear, J.R., Swafford, A.D., Thompson, L.R., Torres, P.J., Trinh, P., Tripathi, A., Turnbaugh, P.J., Ul-Hasan, S., van der Hooft, J.J.J., Vargas, F., Vázquez-Baeza, Y., Vogtmann, E., von Hippel, M., Walters, W., Wan, Y., Wang, M., Warren, J., Weber, K.C., Williamson, C.H.D., Willis, A.D., Xu, Z.Z., Zaneveld, J.R., Zhang, Y., Zhu, Q., Knight, R. and Caporaso, J.G., 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnology 37(8): 852-857. https://doi.org/10.1038/s41587-019-0209-9
Damms-Machado, A., Mitra, S., Schollenberger, A.E., Kramer, K.M., Meile, T., Konigsrainer, A., Huson, D.H. and Bischoff, S.C., 2015. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. BioMed Research International 2015: 806248. https://doi.org/10.1155/2015/806248
De la Cuesta-Zuluaga, J., Kelley, S.T., Chen, Y., Escobar, J.S., Mueller, N.T., Ley, R.E., McDonald, D., Huang, S., Swafford, A.D., Knight, R. and Thackray, V.G., 2019. Age- and sex-dependent patterns of gut microbial diversity in human adults. mSystems 4(4). https://doi.org/10.1128/msystems.00261-19
DeSantis, T.Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E.L., Keller, K., Huber, T., Dalevi, D., Hu, P. and Andersen, G.L., 2006. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Applied and Environmental Microbiology 72(7): 5069-5072. https://doi.org/10.1128/aem.03006-05
Deschasaux, M., Bouter, K.E., Prodan, A., Levin, E., Groen, A.K., Herrema, H., Tremaroli, V., Bakker, G.J., Attaye, I., Pinto-Sietsma, S.-J., van Raalte, D.H., Snijder, M.B., Nicolaou, M., Peters, R., Zwinderman, A.H., Bäckhed, F. and Nieuwdorp, M., 2018. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nature Medicine 24(10): 1526-1531. https://doi.org/10.1038/s41591-018-0160-1
Esteghamati, A., Ashraf, H., Esteghamati, A.-R., Meysamie, A., Khalilzadeh, O., Nakhjavani, M. and Abbasi, M., 2009. Optimal threshold of homeostasis model assessment for insulin resistance in an Iranian population: the implication of metabolic syndrome to detect insulin resistance. Diabetes Research and Clinical Practice 84(3): 279-287.
GBD 2021 Adult BMI Collaborators, 2025. Global, regional, and national prevalence of adult overweight and obesity, 1990-2021, with forecasts to 2050: a forecasting study for the Global Burden of Disease Study 2021. The Lancet 405(10481): 813-838. https://doi.org/10.1016/S0140-6736(25)00355-1
Gryaznova, M., Dvoretskaya, Y., Burakova, I., Syromyatnikov, M., Popov, E., Kokina, A., Mikhaylov, E. and Popov, V., 2022. Dynamics of changes in the gut microbiota of healthy mice fed with lactic acid bacteria and bifidobacteria. Microorganisms 10(5): 1020. https://doi.org/10.3390/microorganisms10051020
Harakeh, S., Angelakis, E., Karamitros, T., Bachar, D., Bahijri, S., Ajabnoor, G., Alfadul, S.M., Farraj, S.A., Al Amri, T., Al-Hejin, A., Ahmed, A., Mirza, A.A., Didier, R. and Azhar, E.I., 2020. Impact of smoking cessation, coffee and bread consumption on the intestinal microbial composition among Saudis: A cross-sectional study. PLoS ONE 15(4): e0230895. https://doi.org/10.1371/journal.pone.0230895
Institute for Health Metrics and Evaluation (IHME), 2025. The Lancet: More than half of adults and a third of children and adolescents predicted to have overweight or obesity by 2050. Institute for Health Metrics and Evaluation, Seattle, WA, USA. Available at: https://www.healthdata.org/news-events/newsroom/news-releases/lancet-more-half-adults-and-third-children-and-adolescents
Koliada, A., Syzenko, G., Moseiko, V., Budovska, L., Puchkov, K., Perederiy, V., Gavalko, Y., Dorofeyev, A., Romanenko, M., Tkach, S., Sineok, L., Lushchak, O. and Vaiserman, A., 2017. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiology 17(1): 120. https://doi.org/10.1186/s12866-017-1027-1
Lee, H.K., Kim, N.E., Shin, C.M., Oh, T.J., Yoon, H., Park, Y.S., Kim, N., Won, S. and Lee, D.H., 2024. Gut microbiome signature of metabolically healthy obese individuals according to anthropometric, metabolic and inflammatory parameters. Science Reports 14(1): 3449. https://doi.org/10.1038/s41598-024-53837-z
Mancabelli, L., Milani, C., Lugli, G.A., Turroni, F., Cocconi, D., van Sinderen, D. and Ventura, M., 2017. Identification of universal gut microbial biomarkers of common human intestinal diseases by meta-analysis. FEMS Microbiology and Ecology 93(12): fix153. https://doi.org/10.1093/femsec/fix153
Naja, F., Nasreddine, L., Itani, L., Chamieh, M.C., Adra, N., Sibai, A.M. and Hwalla, N., 2011. Dietary patterns and their association with obesity and sociodemographic factors in a national sample of Lebanese adults. Public Health Nutrition 14(9): 1570-1578. https://doi.org/10.1017/S136898001100070X
Napoli, T.F., Cortez, R.V., Sparvoli, L.G., Taddei, C.R. and Salles, J.E.N., 2024. Unveiling contrasts in microbiota response: A1c control improves dysbiosis in low-A1c T2DM, but fails in high-A1c cases-a key to metabolic memory? BMJ Open Diabetes Research and Care 12(3): e003964. https://doi.org/10.1136/bmjdrc-2023-003964
Patrone, V., Vajana, E., Minuti, A., Callegari, M.L., Federico, A., Loguercio, C., Dallio, M., Tolone, S., Docimo, L. and Morelli, L., 2016. Postoperative changes in fecal bacterial communities and fermentation products in obese patients undergoing bilio-intestinal bypass. Frontiers in Microbiology 7: 200. https://doi.org/10.3389/fmicb.2016.00200
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J. and Glöckner, F.O., 2013. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research 41(Database issue): D590-596. https://doi.org/10.1093/nar/gks1219
Rinninella, E., Raoul, P., Cintoni, M., Franceschi, F., Miggiano, G.A.D., Gasbarrini, A. and Mele, M.C., 2019. What is the healthy gut microbiota composition? a changing ecosystem across age, environment, diet, and diseases. Microorganisms 7(1): 14. https://doi.org/10.3390/microorganisms7010014
Ritchie, H., 2017. Obesity. Available at: https://ourworldindata.org/obesity
Surono, I.S., Simatupang, A., Kusumo, P.D., Waspodo, P., Verbruggen, S., Verhoeven, J. and Venema, K., 2022. Effect of different functional food supplements on the gut microbiota of prediabetic indonesian individuals during weight loss. Nutrients 14(4): 781. https://doi.org/10.3390/nu14040781
Sze, M.A. and Schloss, P.D., 2016. Looking for a signal in the noise: revisiting obesity and the microbiome. mBio 7(4). https://doi.org/10.1128/mbio.01018-16
Takagi, T., Naito, Y., Inoue, R., Kashiwagi, S., Uchiyama, K., Mizushima, K., Tsuchiya, S., Dohi, O., Yoshida, N., Kamada, K., Ishikawa, T., Handa, O., Konishi, H., Okuda, K., Tsujimoto, Y., Ohnogi, H. and Itoh, Y., 2019. Differences in gut microbiota associated with age, sex, and stool consistency in healthy Japanese subjects. Journal of Gastroenterology 54(1): 53-63. https://doi.org/10.1007/s00535-018-1488-5
Tsai, M.C., Liu, Y.Y., Lin, C.C., Wang, C.C., Wu, Y.J., Yong, C.C., Chen, K.D., Chuah, S.K., Yao, C.C., Huang, P.Y., Chen, C.H., Hu, T.H. and Chen, C.L., 2020. Gut microbiota dysbiosis in patients with biopsy-proven nonalcoholic fatty liver disease: a cross-sectional study in Taiwan. Nutrients 12(3): 820. https://doi.org/10.3390/nu12030820
World Health Organization (WHO), 2016. Diabetes United Arab Emirates 2016 country profile. Available at: https://www.who.int/publications/m/item/diabetes-are-country-profile-united-arab-emirates-2016
World Health Organization (WHO), 2024. One in eight people are now living with obesity. Available at: https://www.who.int/news/item/01-03-2024-one-in-eight-people-are-now-living-with-obesity
World Obesity Federation, 2025. World Obesity Atlas 2025. Available at: https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2025
Yasir, M., Angelakis, E., Bibi, F., Azhar, E.I., Bachar, D., Lagier, J.C., Gaborit, B., Hassan, A.M., Jiman-Fatani, A.A., Alshali, K.Z., Robert, C., Dutour, A. and Raoult, D., 2015. Comparison of the gut microbiota of people in France and Saudi Arabia. Nutrition and Diabetes 5: e153. https://doi.org/10.1038/nutd.2015.3
Yatsunenko, T., Rey, F.E., Manary, M.J., Trehan, I., Dominguez-Bello, M.G., Contreras, M., Magris, M., Hidalgo, G., Baldassano, R.N., Anokhin, A.P., Heath, A.C., Warner, B., Reeder, J., Kuczynski, J., Caporaso, J.G., Lozupone, C.A., Lauber, C., Clemente, J.C., Knights, D., Knight, R. and Gordon, J.I., 2012. Human gut microbiome viewed across age and geography. Nature 486(7402): 222-227. https://doi.org/10.1038/nature11053
Yilmaz, P., Parfrey, L.W., Yarza, P., Gerken, J., Pruesse, E., Quast, C., Schweer, T., Peplies, J., Ludwig, W. and Glöckner, F.O., 2014. The SILVA and ‘all-species living tree project (LTP)’ taxonomic frameworks. Nucleic Acids Research 42 (Database issue): D643-648. https://doi.org/10.1093/nar/gkt1209
Zhang, W., Li, J., Lu, S., Han, N., Miao, J., Zhang, T., Qiang, Y., Kong, Y., Wang, H., Gao, T., Liu, Y., Li, X., Peng, X., Chen, X., Zhao, X., Che, J., Zhang, L., Chen, X., Zhang, Q., Hu, M., Li, Q. and Kan, B., 2019. Gut microbiota community characteristics and disease-related microorganism pattern in a population of healthy Chinese people. Scientific Reports 9(1): 1594. https://doi.org/10.1038/s41598-018-36318-y







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}