Abstract
Lacticaseibacillus paracasei strain Shirota (LcS), promotes intestinal homeostasis, modulates immune cells, and provides anti-stress benefits. Four years of BLP (Biolactis powder: LcS preparation) administration is known to suppress the recurrence of highly dysmorphic polyps in participants who underwent colorectal adenoma resection. Furthermore, adenoma development tended to be suppressed in patients undergoing colorectal adenoma resection who consumed BLP for >20 years, accompanied by a reduction in aging weight loss. However, the underlying mechanisms and effects of prolonged BLP intake on gut mucosa and microbiota remain unclear. Hence, we aimed to analyse the gut microbiota and host gene expression in endoscopically obtained colonic mucosal tissue from participants who had been voluntarily consuming BLP for more than 20 years, as well as from non-consumers of BLP. The faecal and mucosal microbiota of the BLP group revealed a high detection rate and abundance of Coprococcus genus and a rich population of butyrate-producing bacteria. Conversely, the mucosa of the control group was enriched in opportunistic pathogens and environmental bacteria, including those from the families Pseudomonadaceae and Brachyspiraceae. RNA-seq of the colon mucosa of BLP-consuming patients revealed high expression of genes related to the oxidative phosphorylation (OXPHOS) pathway, including those of the mitochondrial electron transfer system. Additionally, T cell- and G-protein-coupled receptor-related genes were overexpressed in BLP-consuming patients. These findings indicate that prolonged BLP intake increases the abundance of butyrate-producing bacteria and activates the OXPHOS pathway in colonic mucosal tissue, which alters the enteroenvironment and limits colonisation by opportunistic pathogens. These findings may contribute to the prevention of colorectal cancer development and have implications for promoting healthy longevity.
Clinical Trial Registry number: 000025389.
1 Introduction
Cancer has been the second leading cause of global deaths in the last decade, and is the leading cause of death in Japan, with colorectal cancer being the second most prevalent form of cancer (Katanoda et al., 2021; Kocarnik et al., 2022). Cancer is generally associated with aging, and its incidence increases with age; therefore, cancer is considered an age-related disease (Berben et al., 2021). Cancer treatments, such as chemotherapy, radiotherapy, immunotherapy, and surgery, are promising but impose significant physical burdens and are potentially life-threatening for older patients (Qu et al., 2023). Additionally, non-responders to immune checkpoint inhibitors are more prevalent among older adults than those in the younger population (Ding et al., 2024). Endoscopic procedures also pose considerable risks, such as bleeding and perforation, in older patients (Iqbal et al., 2022; Noble et al., 2022). Thus, preventing the onset of cancer is a crucial combating strategy in this demographic (Kvakova et al., 2022). Many colorectal cancers are initially observed as adenomas with moderate or greater dysmorphism before progressing to malignancy (He et al., 2020). Prevention requires periodic medical check-ups and control of adenoma development (Schroeder and Tremaine, 1987; Shureiqi, 2022). Regarding cancer prevention, while the regular intake of functional food components, such as folic acid, calcium, vitamin D, n-3 polyunsaturated fatty acids, curcumin, resveratrol, and yoghurt, is effective, no long-term prospective studies have evaluated the intake of specific foods for ten years or more (Hull, 2021; Kvakova et al., 2022).
Intestinal microbiota is intricately linked to host health and disease (Manos, 2022). Disturbances in the gut microbiota, termed dysbiosis, are associated with various ailments, including gastrointestinal diseases (Hou et al., 2022). Mucosa-associated bacteria play crucial roles in the pathogenesis of colorectal cancer and inflammatory bowel disease. Mucosal homeostasis is maintained by preventing the invasion of epithelial cells by Escherichia coli and Proteus bacteria through their flagella with the aid of a glycoprotein called Lypd8, which is expressed on the mucosal epithelium (Okumura et al., 2016). Certain bacteria, including Proteobacteria, Fusobacteria, Bacteroides fragilis, Lachnoclostridium, and Clostridium hathway are detected at high levels in colorectal adenoma, and are potential risk factors for inducing colorectal cancer (Galeano Niño et al., 2022; Liang et al., 2020; Lu et al., 2016; Qu et al., 2023). Therefore, elucidating the state of gut microbiota and tissues in the normal mucosa of older patients is important.
The International Scientific association for Probiotics and Prebiotics in 2013 defined probiotics as ‘probiotics are live microorganisms that, when administrated in adequate amounts confer a health benefit on the host’ (Hill et al., 2014). Probiotics exert various beneficial effects on gut homeostasis, physiology, immune function, and overall health of the host (Gill et al., 2001; Gleeson et al., 2011; Shida et al., 2017). Lacticaseibacillus paracasei strain Shirota (LcS; formerly Lactobacillus casei strain Shirota) is a well-known probiotic which not only confers these benefits but also exhibits anti-tumour properties (S. Matsumoto et al., 2009). Our previous studies demonstrated that the occurrence rate of tumours with a grade of moderate atypia or higher was significantly lower in the group administered Biolactis powder (BLP) containing viable LcS preparation for 4 years (relative risk of 0.65 [95% CI 0.43-0.98]) (Ishikawa et al., 2005). However, the underlying mechanism remains unclear. To investigate these mechanisms, we conducted a follow-up study in which participants who had consumed BLP for 4 years continued to take BLP, for approximately 15 years. The objective of this study was to assess the long-term effects of BLP on adenoma development. Although no statistically significant difference was observed, the recurrence rate of intermediate- and high-grade colorectal adenomas was slightly lower in the group that consumed BLP for 15 years than in the group that did not consume BLP (non-BLP). (cancer development; 24 of 128 (18.8%) in the BLP group, 24 of 109 (22.0%) in the non-BLP group, risk ratio 0.88 [95% CI 0.53-1.47]). Additionally, it also found that BLP intake mitigated weight loss among older participants. (Ishikawa et al., 2020). Recent studies have indicated that elderly Japanese males exhibiting symptoms of sarcopenia demonstrate reduced intestinal microbial diversity and a decline in specific butyrate-producing bacteria compared to their non-frail counterparts (Asaoka et al., 2025). These findings hypothesise that prolonged consumption of BLP may induce favourable modulation in the intestinal milieu, encompassing both the gut microbiota and intestinal mucosal tissue, thereby exerting beneficial effects on subsequent host physiological functions, such as immune response and body weight regulation. Therefore, in this study, we aimed to elucidate the mechanisms of adenoma prevention by examining the effects of long-term probiotic intake on the intestinal microbiota and intestinal tissues, including immune cells.
2 Materials and methods
Study participants and design
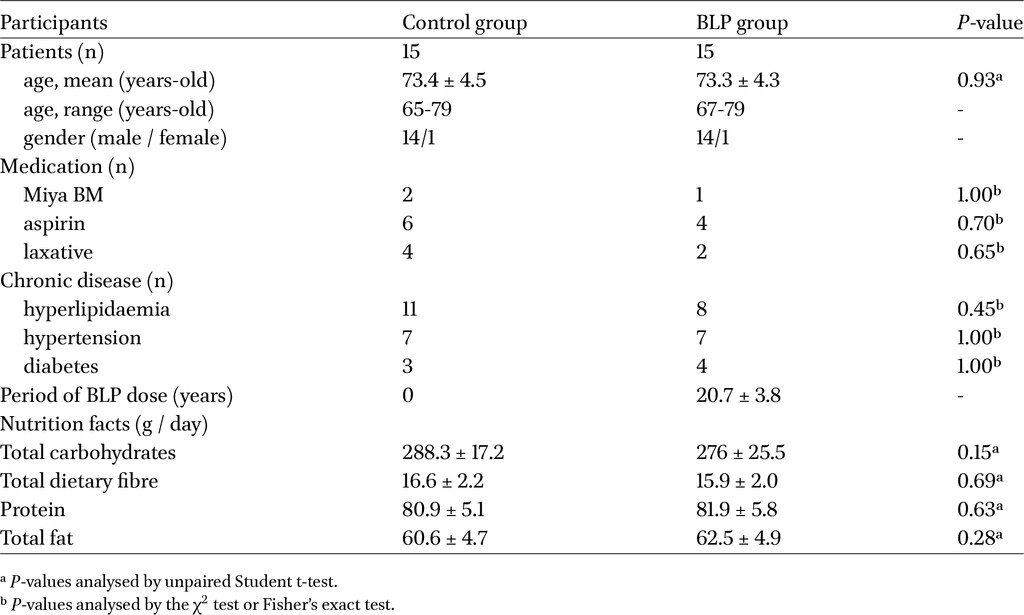
Participants who had previously enrolled in clinical study designed to evaluate the effects of wheat bran and BLP in preventing colorectal cancer (wheat bran biscuit (WBB)/BLP study) (Ishikawa et al., 2005) were included in this study. Approximately 200 individuals voluntarily continued taking BLP after the (WBB)/BLP study concluded. The BLP intake group for this trial was composed of participants selected from among these 200 individuals. They were asked to continue taking the BLP regardless of whether they had been in the BLP group. Participants who desired to continue received BLP. BLP was recommended to be consumed at a dosage of one packet, which contained one gram of LcS preparation, per three times daily following meals. In terms of intake monitoring, participants visited the principal physician’s clinic every three months for health evaluations and to verify their adherence to the intake regimen. Most participants adhered to a regimen of three packets per day. In order to avoid rendering the trial infeasible due to overly restrictive exclusion criteria, the exclusion criteria were established to include individuals who had taken antibiotics within one month prior to specimen collection. Prior to enrolment, participants provided both oral and written informed consent. After obtaining informed consent prior to the trial, nutritional assessments were conducted on the participants using a questionnaire regarding nutritional status over the past one year using Food Frequency Questionnaire (FFQ short version: commissioned from Kenpakusha Co., Ltd.) (Yokoyama et al., 2016). For uniform age and sex distribution, 15 participants who had been taking BLP for more than 20 years were recruited as the intake group, and 15 participants in the non-intake group were recruited as the control group. The principal physician confirmed through medical interviews that, although some participants had chronic conditions, these did not impede their involvement in this trial.
The participants collected stool samples 4 weeks prior to the endoscopy visit. On the day of the examination, 15 ml of blood was collected from each participant, followed by colonoscopy, wherein normal tissue was collected from the ascending colon (ac) and sigmoid colon (sc). The study was conducted in accordance with the guiding principles of the Declaration of Helsinki and was approved by the ethics committee of Kyoto Prefectural University of Medicine (Approval No. ERB-C-1247). Clinical Trial Registry number: 000025389.
Study products
BLP consisted of approximately 1.5 × 109 to 2.1 × 1010 viable LcS cells per gram. BLP was supplied by Yakult Honsha (Minato-ku, Tokyo, Japan). It was transported to the Ishikawa Gastroenterological Clinic (Yodoyabashi, Osaka, Japan) for distribution.
Blood sample
Blood samples were sent to BML (BML, Inc.. Kawagoe-shi, Saitama, Japan) to measure natural killer (NK) cell activity. RNA was extracted from peripheral blood and sent to Yakult Central Institute (Kunitachi-shi, Tokyo, Japan) for T- and B- cell receptor repertoire analysis.
Faecal sample
To evaluate the gut microbiota, participants were provided with comprehensive stool collection kits, which included microbially uncontaminated collection containers. Participants were instructed to collect a stool sample prior to undergoing colonoscopy, ensuring no antibiotic use in the preceding four weeks, and to promptly send the refrigerated samples to the clinic. Upon receipt at the clinic, a nurse assigned a specimen ID to the samples and stored them at −80 °C until use. Once an adequate number of samples was accumulated, the faecal samples were shipped frozen using container with dry ice to the Yakult Central Institute (Kunitachi-shi, Tokyo, Japan), where DNA was extracted and short chain fatty acids (SCFAs) were analysed. The 16S rRNA gene was subjected to gene amplicon analysis using a next-generation sequencer, MiSeq (Illumina, San Diego, CA, USA).
Measurement of total bacterial counts with flow cytometry
Colonic mucosal sample
Fresh, healthy, ascending and sigmoid colonic mucosal tissues were collected via endoscopy and were immediately frozen on dry ice. The specimen IDs were assigned by the attending nurse during endoscopy, and the samples were stored at −80 °C until use. Tissue samples, approximately 3-5 mm3 in size were transferred to the Yakult Central Institute (Kunitachi, Tokyo, Japan). The tissues were then placed in RLT+ buffer containing β-mercaptoethanol and sonicated with ø2 mm zirconia and alumina beads under medium-power conditions (10 cycles of 30 s sonication followed by 30 s rest). The lysates were processed using the AllPrep DNA/RNA Micro Kit (Qiagen, Hilden, Germany) to extract DNA and RNA according to the manufacturer’s protocol. The extracted DNA was used for 16S rRNA gene amplicon analysis to examine mucosa-associated bacteria, and RNA was used for RNA-seq analysis.
16S rRNA gene sequencing and microbial community analysis
Fresh faecal samples were diluted tenfold with PBS and vigorously shaken with sterile glass beads (2 mm in diameter), and 200 μl of the faecal suspension were used for DNA extraction. The faecal suspension was mixed with 500 μl extraction buffer [100 mM Tris-HCl (pH 9.0), 40 mM EDTA, 1% SDS (v/v)], 300 mg of sterile glass beads (0.1 mm in diameter, TOMY), and 500 μl of buffer-saturated phenol (Nippon Gene, Chiyoda-ku, Tokyo, Japan). The mixture was mechanically disrupted using a FastPrep MP (MP, Chuo-ku, Tokyo, Japan) at a power level of 5.0 for 30 s. The mixtures were then centrifuged at
RNA sequencing analysis
The RNA aliquots were sent on dry ice to Macrogen (Seoul, South Korea) for comprehensive RNA sequencing (RNAseq) analysis, which culminated in the generation of fastq.gz files.

Baseline characteristics of the participants
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Library preparation and sequence data acquisition
Since the RNA samples with DV200 values, indicating the percentage of RNA fragments above 200 nucleotides, exceeded the threshold of 50%, we prepared libraries using the TruSeq RNA Exome kit (Illumina). After preparation, a few samples did not meet the criteria for NGS. Ultimately, 12 samples from ac mucosal tissue and 12 samples from sc mucosal tissue were sequenced using the NovaSeq6000 platform to produce fastq.gz files.
Bioinformatic and statistical analysis
Fastq.gz files were trimmed and quality checked using Fastp (Chen, 2023). Read counts for each gene in the filtered FASTQ files were quantified against GENCODE human annotation version 41 using Salmon (version 1.4.0) (Patro et al., 2017). Technical replicates were averaged for each biologically independent sample. Gene counts were normalised to library size, and differential expression analysis was conducted using DESeq2 (v.1.30.1) (Love et al., 2014). Wald tested P-values were subjected to Benjamini-Hochberg correction for multiple testing, and gene sets with corrected P-values <0.05 were deemed significantly differentially expressed. These gene sets were further analysed using over-representation analysis (ORA), which was performed using ClusterProfiler (v 4.4.4) (Yu et al., 2012) with hallmark gene sets in the MsigDB database (7.5.1) (Liberzon et al., 2015), CellMarker_Augmented_2021, and Reactome_2022 in enrichR (Kuleshov et al., 2016).
Quantification and statistical analyses
Data were analysed using GraphPad Prism 8 software (GraphPad Software, San Diego, CA, USA). To compare the two groups, P-values were determined using the unpaired two-tailed Student’s t test, Mann-Whitney U test, the
3 Results
Characteristics of participants
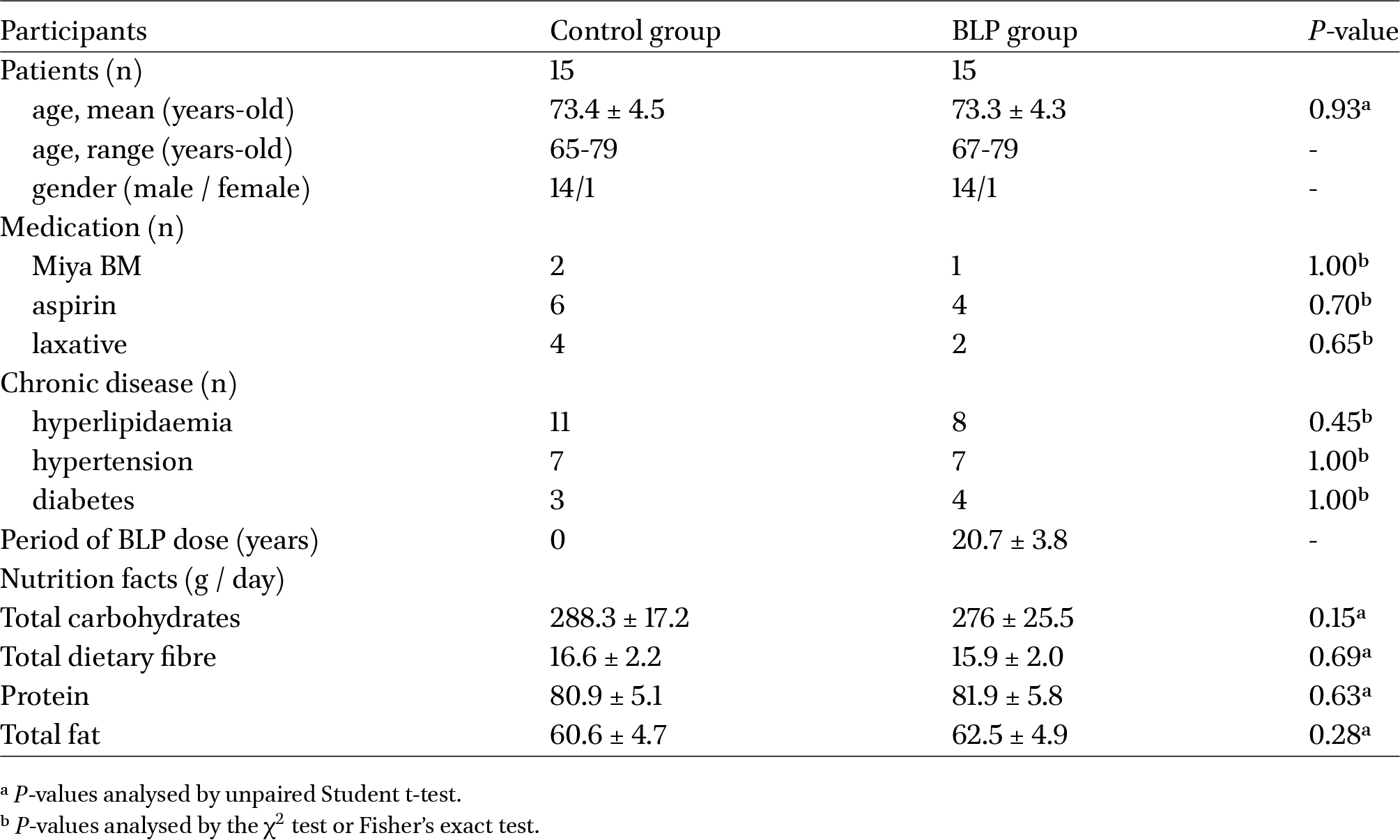
The BLP and control groups each was comprised of 14 males and one female, with mean age of 73.4 ± 4.5 years and 73.3 ± 4.3 years, respectively (Table 1). Participants in both the groups had undergone colonic adenoma resection approximately 15-20 years prior. In the BLP group, 14 patients had incessantly consumed 3 g of LcS preparation (1.5-21.0 × 109 cfu / g) per day, and one patient in the BLP group had consumed 2 g of BLP per day. There were no specific dietary restrictions for either group. Although no participant met the exclusion criteria, the principal physician determined that one individual was unsuitable for an intestinal biopsy. Consequently, neither a biopsy nor peripheral blood collection was performed for this participant. Regarding additional medications, one participant in the BLP group and two in the control group regularly consumed MiyaBM (Miyarisan Pharmaceutical, Tokyo, Japan). Furthermore, four participants in the BLP group and six in the control group regularly took aspirin, whereas two participants in the BLP group and four in the control group regularly used laxatives (Table 1). Regarding chronic disease among participants, eight participants in the BLP group and eleven in the control group had hyperlipidaemia, seven participants in the BLP group and seven in the control group had hypertension, and four participants in the BLP group and three in the control group had diabetes (Table 1). There were no statistically significant differences in medication usage and current medical history between the groups, and these medications and medical history were not excluded in our analyses. The findings from the FFQ-based nutritional survey revealed no statistically significant differences in the intake of carbohydrates, proteins, or total fat between the BLP group and the control group, suggesting an absence of variation in nutritional intake status (Table 1).
The faecal samples of BLP intake participants were abundant in Coprococcus genus and Lachnospiraceae family
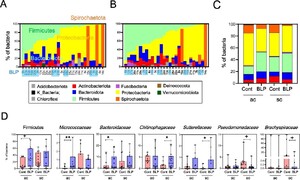
The total bacterial counts and alpha diversity (Shannon index) were similar between the two groups. However, beta diversity (Jaccard distance) varied significantly between the groups (Figures 1A-C). Although both the groups were similar at the microbial phylum level, a detailed analysis at the bacterial genus level revealed a higher detection rate, abundance, and counts of L. paracasei and Coprococcus genus in the BLP group than those in the control group (Figures 1D-G, Supplementary Figure S1). LDA at the genus level indicated that, in addition to Lacticaseibacillus and Coprococcus, Lachnospiraceae family, including Dorea and NK4A214, Oscillospiraceae family UCG-002, Christensenellaceae family Chiristensenella were enriched in the BLP group (Figures 1H-I). Furthermore, a higher prevalence of Eubacterium coprostanoligenes and Erysipelotrichaceae was observed in the BLP group, compared to that in the control group (Supplementary Figure S2). These findings suggest that long-term BLP intake may increase the abundance of the Lachnospiraceae family, particularly that of genus Coprococcus, in the faecal bacteria of older adults following colonic adenoma resection.
Comparison of faecal microbiota composition between the Biolactis powder (BLP) containing Lacticaseibacillus paracasei strain Shirota (LcS) and control (Cont) groups. (A) Total bacterial counts. (B) Alpha diversity (Shannon index). (C) Beta diversity (Jaccard index). Statistical analyses were performed using PERMANOVA test between the groups. (*
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
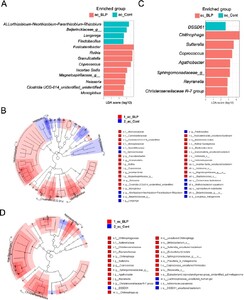
Colonic mucosa-associated bacteria in the BLP group: Enrichment of Lachnospiraceae and Bacteroidaceae, reduced abundance of Proteobacteria and Spirochaetota
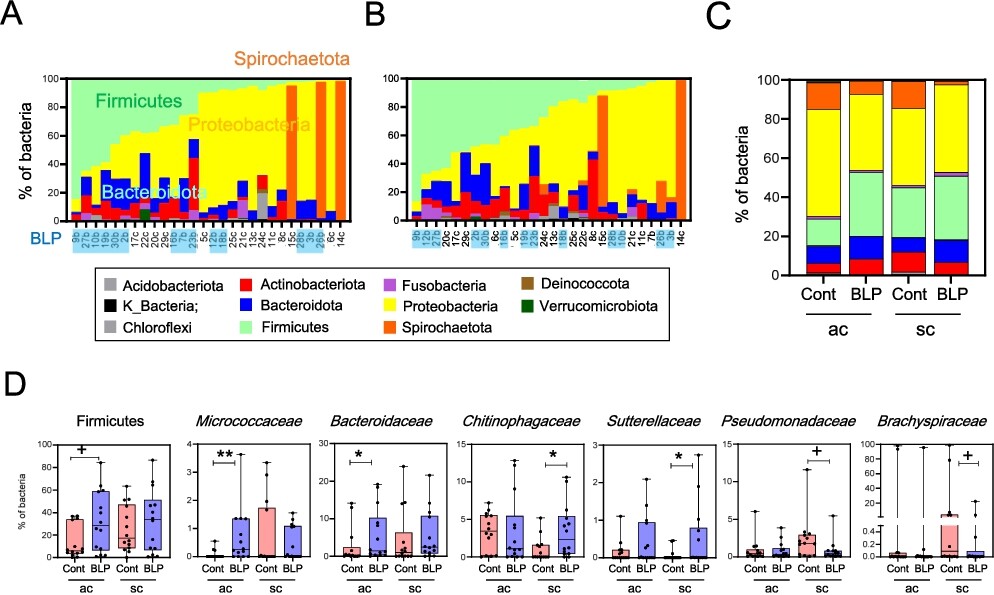
Subsequent to the colonic mucosa-associated bacterial analysis, diversity (observed features) tended to increase in the ac of the BLP group compared to that in the control group (Supplementary Figure S3A). Beta diversity (Jaccard distance) in both the ac and sc tended to differ between the two groups (Supplementary Figure S3B). At the phylum level, Proteobacteria and alternately Spirochaetota were the most prevalent in, both, the ac and sc mucosa (Figure 2A). Notably, the ac and sc mucosa in the BLP group exhibited a higher abundance of Firmicutes, Bacteroidota, and Actinobacteria, whereas the control group displayed a higher abundance of Proteobacteria (Figures 2A and B). In particular, the ac of the BLP group exhibited a higher abundance of Firmicutes, Micrococcaceae, and Bacteroidaceae than that in the control group (Figures 2C and D). Meanwhile, in the sc, the BLP group displayed a higher abundance of Chitinophagaceae and Sutterellaceae and lower abundances of Pseudomonadaceae and Brachyspiraceae (Figure 2D). LEfSe further revealed enrichment of the genus Rothia from the Micrococcaceae family, and those of Fusicatenibacter and Coprococcus from the Lachnospiraceae family in the ac of the BLP group (Figures 3A and B). In contrast, the ac of the control group exhibited enrichment of the alpha proteobacteria classes Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium genera and Beijerickiaceae genera, Sporichthyaceae family Longivirga genera and Chitinopahagaceae family Flectobacillus genera (Figures 3A and B). Additionally, in the sc of the control group, DSSD61 genera of the Burkholdelriaceae family exhibited a lower abundance, whereas Chitinophaga genera from the Chitinophagaceae family, Sutterella from the Sutterellaceae family, Reyranella genera from the Reyranellaceae family, Sphingomonadaceae family, Coprococcus and Agathobacter genera from the Lachnospiraceae family, and Chiristensenellaceae family were abundant in the BLP group (Figures 3C and D).
Comparison of mucosa-associated bacteria (MAB) in the ascending colon (ac) and sigmoid colon (sc) between the Biolactis powder (BLP) containing LcS intake group and the non-intake control (Cont) group. (A-B) Stacked bar graph of MAB abundance at the phylum level. Individual data of MAB abundance are presented with the horizontal axis showing subject ID, and the BLP group highlighted in light blue. MAB abundance in ac (A) and sc (B). (C) Stacked bar graph of averaged MAB abundance at the phylum level. (D) Analysis of Firmicutes and intestinal bacterial abundance at the family level. Statistical analysis was performed using the Mann–Whitney U test between the BLP and Cont groups (+
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Different structures of mucosa-associated bacteria (MAB) in the ascending colon (ac) and sigmoid colon (sc) between the Biolactis powder (BLP) containing LcS intake group and the non-intake control (Cont) group determined using LEfSe analysis. (A-B) Linear discriminant analysis (LDA) of genus-level abundance in 16S of ac (A) and sc (B). (C-D) Cladogram indicating the taxonomic differences between the BLP and Cont groups in the MAB of ac (C) and sc (D). Red denotes taxa enriched in the BLP group, while blue denotes taxa enriched in the Cont group. The diameter of each circle is proportional to the abundance.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Elevated expressions of genes involved in oxidative phosphorylation pathways in the colonic mucosal tissue following prolonged BLP intake
RNA-seq analysis was performed on mucosal tissue samples from the ac and sc. Initially, principal component analysis was conducted, which revealed remarkable differences in gene expression in the ac, but not sc, of the BLP and control groups (Supplementary Figures S4A and B). Subsequently, ORA of the upregulated genes in the ac mucosal tissue using the Hallmark database in MsigDB demonstrated higher levels of oxidative phosphorylation (OXPHOS) pathway related genes in the BLP group compared to those in the control group. These genes included NADH dehydrogenase (NDUF), Ubiquinol-Cytochrome C Reductase (UQCR), cytochrome C oxidase subunit (COX), ATP synthase membrane subunit c (ATP5), and members of the mitochondrial carrier family (SLC25) (Figure 4A). Furthermore, the gene expressions of carnitine palmitoyltransferase (CPT1A) and 2.4-dienoyl CoA reductase (DECR1) in the mucosa of the BLP group was higher than that in the control group (Figure 4B).
(A-B) Over-representation analysis (ORA) of mucosal tissue in the ascending colon (A) and sigmoid colon (B). (A-B) ORA was conducted using the Hallmark gene sets in MsigDB (7.5.1), focusing on genes associated with the oxidative phosphorylation pathway. The results are presented in a heatmap format. Key genes related to this pathway are colour-highlighted. NDUF (NADH oxidoreductase subunit), UQCR (ubiquinol-cytochrome c reductase), COX (cytochrome c oxidase), ATP5 (ATP synthase), and SLC25 (mitochondrial solute carrier).
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Upregulated expressions of genes associated with T cells and G protein-coupled receptor in colonic mucosal tissue following prolonged BLP intake
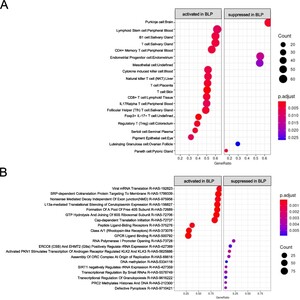
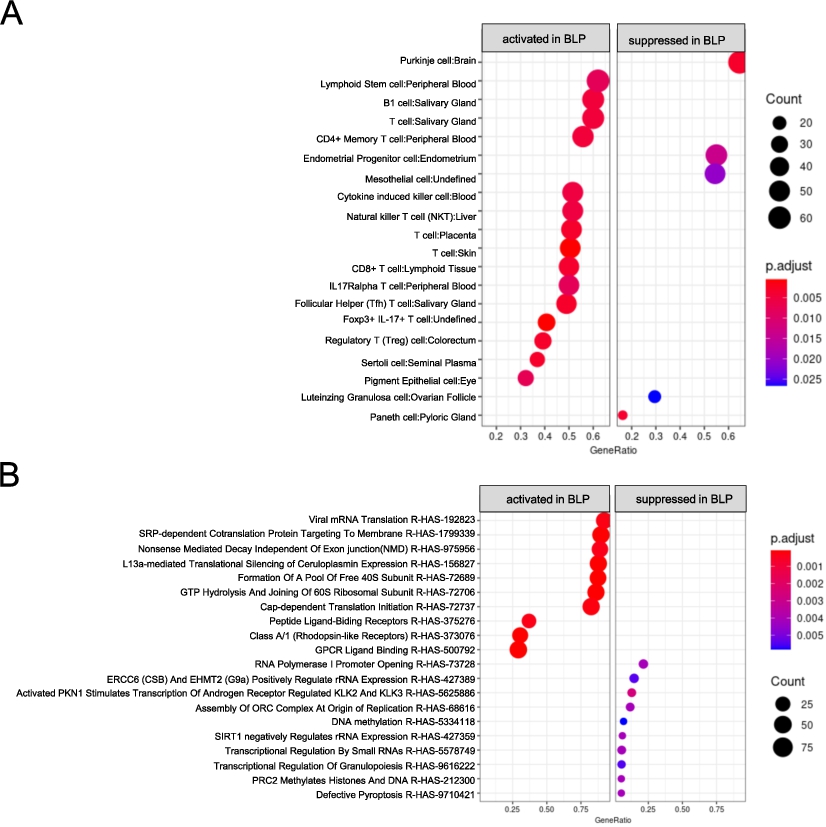
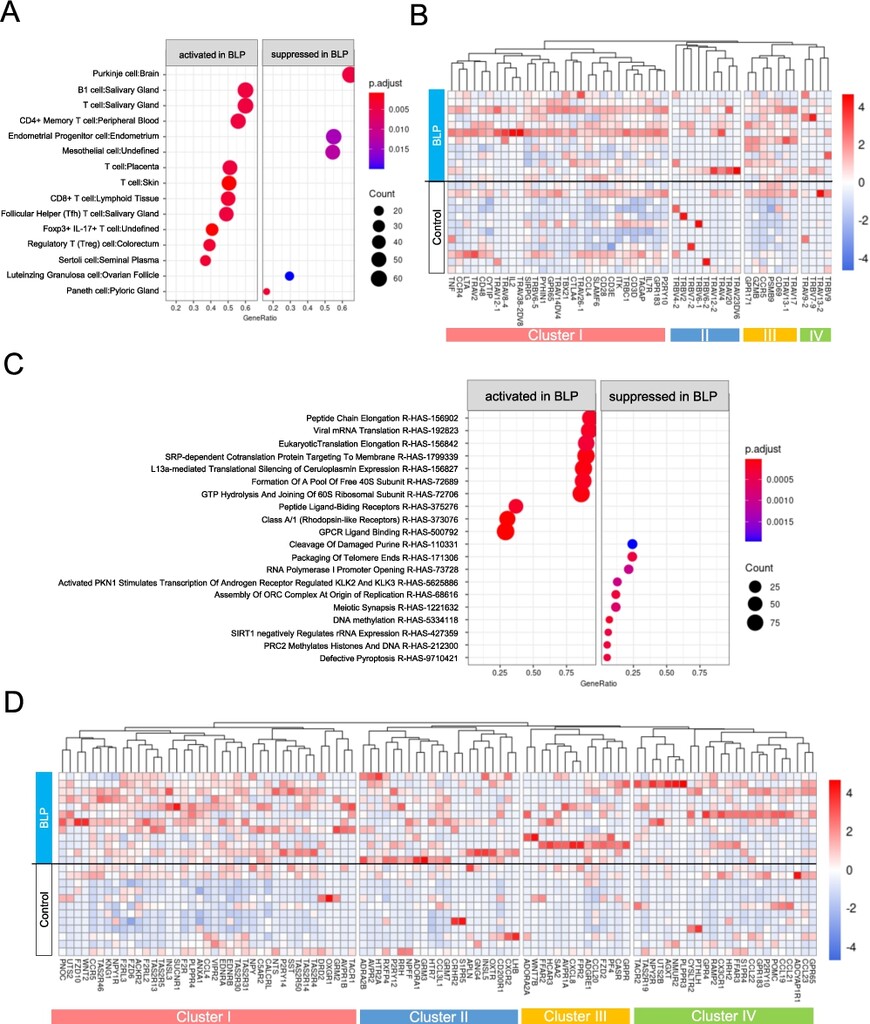
Subsequently, ORA was conducted using the CellMarker database, to assess enriched cell types in the BLP group. The ac mucosal tissue in the BLP group was enriched in B1 cells and numerous T cell subsets, including CD4+ Memory, CD8+, Follicular Helper, Foxp3+IL-17+, and Regulatory (Treg) T cells (Figure 5A). A detailed heatmap examination of T cells and salivary glands revealed that the expression of genes included in Cluster III (CD69, PSMB9, CCR5, GZMB, and GPR171) was high in all participants (Figure 5B).
Over representation analysis (ORA) in ac mucosal tissue. (A-B) ORA was performed using CellMarker_Augmented 2021 gene sets from enrichr. (A) Dot plot analysis was performed, where ‘activated’ indicates gene sets enriched in the BLP group, ‘suppressed’ indicate sets enriched in the Cont group. (B) Heat map analysis of genes related to T cells was performed. Hierarchical clustering was conducted using the group average method based on gene expression patterns, resulting in clusters I to IV. (C-D) ORA was performed using the Reactome gene sets from enrichr. (C) The results are presented in a dot plot. (D) Heat map analysis of genes related to G protein-coupled receptors (GPCRs) was conducted. Hierarchical clustering was performed using the group average method based on gene expression patterns, resulting in classification into cluster I to IV.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Further, ORA using the Reactome database, focusing on biological signalling pathways, revealed the upregulation of transcription/translation-related pathways, such as Peptide Chain Elongation and Viral mRNA translation, as well as G Protein-Coupled Receptor (GPCR)-related signalling pathways, such as Class A/1 (Rhodopsin-like Receptors) and GPCR Ligand Binding in the ac of the BLP group. Heatmap analysis of genes contributing to GPCR Ligand Binding suggested the upregulation of genes in cluster I in the ac BLP group (Figures 5C and D). The expression of various GPCRs such as TAS2R, P2RY14, NPY, and SUCNR1 was elevated (Figure 5D). A total of 97 GPCR-related genes, including 13 chemokines, three SCFA receptors, 15 lipid receptors, nine taste receptors, five adenosine receptors, 15 neuropeptide receptors, 17 hormone receptors, and eight angiotensin receptors, were upregulated in the BLP group (Table 2).
Upregulated G-protein-coupled receptor-related gene list from ascending colonic mucosa in BLP group
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
Network analysis revealed that GPR65, GPR174, S1PR4, and P2RY10 were present in both the T-cell network analysis and GPCRs table (Table 2, Figure S5). In the sc of the BLP group, a similar ORA was performed using the CellMarker and Reactome databases, yielding results comparable to those observed in the ac of the BLP group (Figures 6A and B, Supplementary Figures S6 and S7).
Over-representation analysis (ORA) in the sc mucosal tissue. (A-B) ORA was performed using the CellMarker_Augmented 2021 gene sets (A) and Reactome gene sets (B) from enrichr. Dot plot analysis was performed to illustrate the results. ‘activated’ indicates gene sets enriched in the BLP group, while ‘suppressed’ indicates gene sets enriched in the Cont group.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00107
The impact of prolonged BLP intake on peripheral immune cells
No significant difference was observed in the cytotoxic activity of peripheral NK cells between the BLP and control groups (Supplementary Figure S8). B cell receptor (BCR) repertoire analysis of peripheral blood demonstrated that use of the IgHV7 clone in IgA2+ B cells was significantly higher in the BLP group than that in the control group. Additionally, the use of IgHV5 and IgHV7 in IgM+ B cells in the BLP group tended to be higher than that in the control group (Supplementary Figure S9A). Furthermore, the usage of TRBV3 in the T cell receptor (TCR) β chain in the BLP group was significantly lower than those in the control group (Supplementary Figure S9B).
4 Discussion
Over 20 years intake of BLP affected intestinal microbiota following colonic adenoma resection. In the faecal samples of participants with prolonged BLP intake, an increased abundance of Lacticaseibacillus and Coprococcus genera was observed. Furthermore, Eubacterium coprostanoligenes and the Erysipelotrichaceae family also tended to be more abundant (Figure 1C, 1E-G, Supplementary Figure S3). LDA indicated that Coprococcus, Dorea and NK4A214 from the Lachnospiraceae family, as well as UCG-002 and Christensenella from the Oscilloscpiraceae family were enriched in the faeces of the BLP group (Figure 1H-I). Notably, except for Lacticaseibacillus, all remaining bacteria were butyrate-producing bacteria (BPB) (Tian et al., 2023). BPB and butyrate production are closely associated with gastrointestinal health. In colorectal carcinogenesis, butyrate and BPB level decrease, and the administration of butyrate-resistant starch potentially increases the abundance of BPB and suppresses the increase in oncogenic miRNAs (Hodgkinson et al., 2023). Unfortunately, in the present study, no differences in faecal butyric acid concentrations were observed between the two groups (Supplementary Figure S10). Regarding this point, it is plausible that butyrate produced in the intestine serves as an energy source for colonic epithelial cells through its absorption (Salvi et al., 2021). Additionally, considering the upregulation of GPCR gene expression, we propose that immune cells expressing butyrate receptors such as FFAR3 may utilise butyrate (Table 2). Alternatively, drinking fermented milk containing LcS in healthy adults with soft stool symptoms increases the levels of total organic acids, including butyric acid, and improves stool condition (K. Matsumoto et al., 2010). It is unclear whether butyric acid detection differed among participants who consumed LcS; however, the possibility of butyric acid production due to LcS consumption has been demonstrated.
Prolonged BLP intake impacted on the gene expression profile of colonic mucosal tissues. The expression of genes related to the OXPHOS pathway was higher in both the ac and sc of the BLP group than that in the control group (Figure 4). NDUF, UQCR, and COX are components of the respiratory chain complex I, III, and IV, respectively, whereas ATP5 and SLC25 function as ADP/ATP carriers; all the genes are associated with mitochondrial function. The upregulation of CPT1A, the rate-limiting enzyme for β-oxidation, and DECR1, which reduces unsaturated fatty acid content, indicates activation of the fatty acid metabolic pathway. These findings suggest an increase in mitochondrial activity in the colonic mucosal tissue in the BLP group. However, it remains unclear which specific cellular OXPHOS pathway is activated, representing a limitation of bulk RNA-seq analysis of the whole tissue. Further investigations are required to elucidate address this gap.
Long-term intake of BLP modulated the balance between anaerobic and opportunistic bacteria in colonic mucosa. The colonic mucosa and faecal samples of the BLP group revealed a high abundance of anaerobic Firmicutes and Bacteroidaceae, particularly BPB, such as Coprococcus, Dorea, Oscillospiraceae_UCG-002, Lachnospiraceae_NK4A214, Christensenella, Fusicatenibacter and Lachnospiraceae_Agathobacter (Figures 1E-I, 2A-D, 3A-D). In contrast, the phyla Proteobacteria and Spirochaetota, which include many facultative anaerobes and opportunistic pathobionts, were enriched in the control group (Figure 2D) (Freedberg et al., 2018; Tanahashi et al., 2008). In addition, LDA of ac and sc mucosa in the control group indicated the enrichment of aerobic and nitrogen-reducing bacteria, such as Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium and Beijerinckiaceae, environmental bacteria Flexibacteriaceae family Flexibacteria, Sporichthyaceae family Longivirga and Nitrosomonadaceae family DSSD61. The observed variation in this phenomenon is attributed to differences in the anaerobic state of the intestinal mucosa. Yael Litvac and colleagues, as well as Kendra Hodkinson and her team, have discussed the physiological role of OXPHOS activation in epithelial cells (Hodgkinson et al., 2023; Litvak et al., 2018). They suggested that fermented products, such as butyrate, produced by obligate anaerobes, maintain epithelial cells in a state of high oxygen consumption via the OXPHOS pathway. The epithelial cells are defined as C2 cells. This polarisation sustains anaerobic conditions by ensuring hypoxia in the epithelium and restricting oxygen diffusion into the lumen. In contrast, C1 cells transition to a low-oxygen-consuming state reliant on glycolysis, thereby increasing O2 and NO3− concentrations in the lumen. Based on this, the BLP group appeared to have a higher prevalence of C2 cells, supporting the proliferation of obligate anaerobic bacteria. Therefore, prolonged BLP intake presumably activates the OXPHOS pathway in the colonic mucosal epithelium, thus promoting anaerobic conditions, and inducing the colonisation of obligate anaerobes and mucosal commensal bacteria (Supplementary Figure S11). This may inhibit the colonisation of opportunistic bacteria.
The prolonged consumption of BLP influenced the composition of immune cells in the colonic mucosal tissue. ORA revealed the enrichment of various T cell subsets and B cells in the BLP group (Figure 5A). Immune cells exhibiting effector functions predominantly rely on glycolysis, whereas immune tolerance, memory, and wound healing processes are dependent on OXPHOS, including fatty acid beta-oxidation. The former category includes effector CD8+ T cells, Th1, Th2, Th17, and M1 macrophages, whereas the latter category encompasses naïve T cells, central memory T cells, Tregs, M2 macrophages, and bone marrow-derived immunosuppressive cells (Ashton et al., 2018; Ghosh et al., 2020; Maciver et al., 2013). Based on these findings, it is plausible that CD4+ memory T cells and Tregs were enriched in the BLP group. Considering that the upregulation of T cell-related chemokine (CCR4 and CCR5) gene expression was observed in the BLP group, prolonged BLP intake was postulated to facilitate T cell migration into the colonic mucosa (Figure 5B, Supplementary Figure S5).
Furthermore, the expression of GPCR-related genes was notably higher in the BLP group than that in the control group. Notably, GPR65, GPR174, S1PR4, and P2RY10 were expressed at higher levels in T cells and GPCRs in the BLP group (Figure 5C and 5D, Supplementary Figure S5, Table 2). GPR65, an extracellular proton-sensing GPCR, plays a role in mucosal homeostasis by regulating the expression of antimicrobial peptides on colonic epithelial cells (Li et al., 2023; Marie et al., 2022). GPR174, S1PR4, and P2RY10 are receptors for lysophosphatidylserine, a lysophospholipid subtype known to inhibit T cell proliferation and promote cell migration and macrophage phagocytosis (Makide et al., 2014). Additionally, the BLP group exhibited higher expression of lipid-related receptors, such as oxysterol receptors (GPR183), sphingosine 1-phosphate receptors (S1PR4 and S1PR5), and SCFA receptors (FFAR2, FFAR3, and SUCNR1) (Brown et al., 2023; Emgård et al., 2018; Kinjo et al., 2008; Tan et al., 2014). These findings suggest that prolonged BLP intake may be associated with lipid metabolism and cell signalling in the gut.
The long-term consumption of BLP led to modifications in the peripheral immune system. Drinking fermented milk containing LcS is known to enhance NK cell activity in healthy adults (Shida et al., 2017). However, the NK cell activity was similar between the BLP and control groups (Supplementary Figure S8). In contrast, BCR repertoire analysis revealed a higher usage of IgH7 clones in IgA2 and IgM in the BLP group (Supplementary Figure S9). This suggests that prolonged BLP intake significantly influences the adaptive immune system in the periphery.
From an anti-cancer perspective, opportunistic bacteria colonisation represents a significant risk factor for cancer development and could serve as a target for preventive strategies. In the present study, the BLP group showed low abundances of Pseudomonadaceae and Brahyspiraceae. The Family Pseudomonadaceae, which includes pathobionts such as P. aeruginosa, is prevalent in mucosal colorectal adenomas, suggesting a potential association with adenoma development (Lu et al., 2016; Thomas et al., 2016). Although many individuals harbouring Brachyspiraceae are asymptomatic, some have been linked to other complications or intestinal diseases, indicating that the presence of this bacterial family may trigger additional infections and diseases (Tanahashi et al., 2008). Thus, prolonged BLP intake might be an important factor in cancer development as shown in Supplementary Figure S11; however, further investigations using more pronounced molecular biological methods and culture techniques are warranted.
Over 15 years of BLP intake demonstrated a weight loss suppression effect in older participants following colonic adenoma resection, with this analysis indicating an increase in butyrate-producing bacteria in the gut. Additionally, a reduction in gut microbial diversity and a decrease in butyrate-producing bacteria have been documented in older males exhibiting symptoms of sarcopenia (Asaoka et al., 2025). Butyrate serves as an energy source for colonic epithelial cells and plays a role in maintaining intestinal homeostasis (Salvi et al., 2021). Based on these observations, we hypothesised that the maintenance of intestinal homeostasis by butyrate may enhance nutrient absorption efficiency, thereby contributing to the suppression of weight loss. Although further research is warranted, this finding is considered noteworthy from the perspective of geriatric nutrition.
The long-term consumption of BLP is suggested to modulate the gut microbiota and colonic mucosal gene expression profiles in participants with colonic adenoma resection. While the findings of this long-term study are both rare and valuable, quite a few limitations were noted. Dietary interventions involving a fermented milk drink containing with lactic acid bacteria, prebiotics, postbiotics, and other supplements pose ethical and logistical challenges in the context of long-term studies. Although we confirmed that the participants did not regularly or habitually consume fermented milk drinks containing lactic acid bacteria, it remains possible that they ingested other functional foods. Additionally, considering the recent advances in research on the interaction between chronic disease, medication, and gut microbiota, the limitation of medication intake and the medical history of the older participants present a significant challenge (Takagi et al., 2020; Vich Vila et al., 2020). Furthermore, RNA-seq analysis of whole tissues limits the ability to identify specific cells which activate the OXPHOS pathway. Future studies should employ detailed analyses using single-cell RNA-sequencing, spatial transcriptome analysis, or in vitro validation tests with organoids. Although this study observed a high abundance of BPB, no increase in butyrate levels was observed. The role of butyrate for this hypothesis needs to be established using alternative experimental systems. These considerations are essential to understand the long-term effects of BLP intake on host health. We are committed to pursuing the aforementioned analyses to elucidate the beneficial role of BLP intake in cancer prevention, which will ultimately contribute to healthy longevity.
Corresponding authors; e-mail: tomoaki-naito@yakult.co.jp; tetsuji-hori@yakult.co.jp
these authors contributed equally to this study
Supplementary Materials and methods
Figure S1. Detect of L. paracasei feature.
Figure S2. Number of faecal bacterial at family level
Figure S3. Mucosa-associated bacterial diversity and taxonomic analysis
Figure S4. Principal component analysis based on gene expression estimates for the BLP and Control group
Figure S5. Network analysis of ORA analysis with Cell Marker in ac mucosal tissue
Figure S6. Network analysis of ORA analysis with Cell Marker database in sc mucosal tissue
Figure S7. Network analysis of ORA analysis with Reactome database in sc mucosal tissue
Figure S8. NK cell activity in peripheral blood mononuclear cells
Figure S9. B cell receptor and T cell receptor clonal usage analysis in peripheral blood mononuclear cells
Figure S10. Result of faecal pH and various organic acid measurement
Figure S11. Gut mucosal niches in older adults following adenoma resection (hypothesis)
Table S1. Primer information for TCRβ repertoire analysis
Acknowledgements
We would like to thank the participants of this study. Finally, we would also like to thank Editage (www.editage.com) for the English language editing.
Authors’ contribution
Conceptualisation, TN, TH, SM and HI; methodology, TN, TH, SM and HI; sample collection, TH, HT, SM and HI; formal analysis, TN, TH, AM, TS, NN and TS; writing – original draft, TN; writing – review and editing, TN, TH, KO, SM and HI; resources, TH, HT, SM and HI; supervision, TH, SM and HI. All authors have approved the final version of the manuscript.
Conflict of interest
TN, TH, ST, AM, KO, NN, TS, HT and SM were employed by Yakult Honsha, who prepared the LcS. HI was founded by Yakult Honsha. Yakult Honsha Co., Ltd. funded the datacentre consignment fee. The funders had no role in the study design, data collection and analysis, decision to publish, or manuscript preparation.
Data availability
16S rRNA-gene amplicon data and RNA-sequencing data were deposited into NCBI sequence Read Archive (SRA) database under the BioProject ID number PRJNA1270739. Any additional information required to reanalyse the data reported in this paper is available on request to the authors.
Lead contact
Further information and requests for reagents may be directed to and fulfilled by the lead contacts, Tomoaki Naito (tomoaki-naito@yakult.co.jp) and Tetsuji Hori (tetsuji-hori@yakult.co.jp).
References
Asaoka, D., Toda, K., Yoshimoto, S., Katsumoto, N., Odamaki, T., Iwabuchi, N., Tanaka, M., Xiao, J.Z., Nishikawa, Y., Nomura, O., Takeda, T., Nagahara, A., Koido, S., Ohkusa, T. and Sato, N., 2025. Sex-specific associations of gut microbiota composition with sarcopenia defined by Asian working group for sarcopenia 2019 consensus in older outpatients: Prospective cross-sectional study in Japan. Nutrients 17: 1746. https://doi.org/10.3390/nu17101746
Ashton, T.M., Gillies McKenna, W., Kunz-Schughart, L.A. and Higgins, G.S., 2018. Oxidative phosphorylation as an emerging target in cancer therapy. Clinical Cancer Research 24: 2482-2490. https://doi.org/10.1158/1078-0432.CCR-17-3070
Berben, L., Floris, G., Wildiers, H. and Hatse, S., 2021. Cancer and aging: Two tightly interconnected biological processes. Cancers 13: 1-20. https://doi.org/10.3390/cancers13061400
Brown, E.M., Clardy, J. and Xavier, R.J., 2023. Gut microbiome lipid metabolism and its impact on host physiology. Cell Host and Microbe 31: 173-186. https://doi.org/10.1016/j.chom.2023.01.009
Chen, S., 2023. Ultrafast one-pass FASTQ data preprocessing, quality control, and deduplication using fastp. IMeta 2: 1-5. https://doi.org/10.1002/imt2.107
Ding, Y., Lei, S., Wang, L., Tang, L., Zhang, Y., Liao, Y., Deng, X., Li, Y., Gong, Y. and Li, Y., 2024. Age-related efficacy of immunotherapies in advanced non-small cell lung cancer: a comprehensive meta-analysis. Lung Cancer 195: 107925. https://doi.org/10.1016/j.lungcan.2024.107925
Emgård, J., Kammoun, H., Garcı́a-Cassani, B., Chesné, J., Parigi, S.M., Jacob, J.M., Cheng, H.W., Evren, E., Das, S., Czarnewski, P., Sleiers, N., Melo-Gonzalez, F., Kvedaraite, E., Svensson, M., Scandella, E., Hepworth, M.R., Huber, S., Ludewig, B., Peduto, L., Villablanca, E.J., Veiga-Fernandes, H., Pereira, J.P., Flavell, R.A. and Willinger, T., 2018. Oxysterol sensing through the receptor GPR183 promotes the lymphoid-tissue-inducing function of innate lymphoid cells and colonic inflammation. Immunity 48: 120-132.e8. https://doi.org/10.1016/j.immuni.2017.11.020
Freedberg, D.E., Zhou, M.J., Cohen, M.E., Annavajhala, M.K., Khan, S., Moscoso, D.I., Brooks, C., Whittier, S., Chong, D.H., Uhlemann, A.C. and Abrams, J.A., 2018. Pathogen colonization of the gastrointestinal microbiome at intensive care unit admission and risk for subsequent death or infection. Intensive Care Medicine 44: 1203-1211. https://doi.org/10.1007/s00134-018-5268-8
Galeano Niño, J.L., Wu, H., LaCourse, K.D., Kempchinsky, A.G., Baryiames, A., Barber, B., Futran, N., Houlton, J., Sather, C., Sicinska, E., Taylor, A., Minot, S.S., Johnston, C.D. and Bullman, S., 2022. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature 611: 810-817. https://doi.org/10.1038/s41586-022-05435-0
Ghosh, P., Vidal, C., Dey, S. and Zhang, L., 2020. Mitochondria targeting as an effective strategy for cancer therapy. International Journal of Molecular Sciences 21: 1-19. https://doi.org/10.3390/ijms21093363
Gill, H.S., Rutherfurd, K.J. and Cross, M.L., 2001. Dietary probiotic supplementation enhances natural killer cell activity in the elderly: An investigation of age-related immunological changes. Journal of Clinical Immunology 21: 264-271. https://doi.org/10.1023/A:1010979225018
Gleeson, M., Bishop, N.C., Oliveira, M. and Tauler, P., 2011. Daily probiotic’s (Lactobacillus casei Shirota) reduction of infection incidence in athletes. International journal of sport nutrition and exercise metabolism 21: 55-64.
He, X., Hang, D., Wu, K., Nayor, J., Drew, D.A., Giovannucci, E.L., Ogino, S., Chan, A.T. and Song, M., 2020. Long-term risk of colorectal cancer after removal of conventional adenomas and serrated polyps. Gastroenterology 158: 852-861.e4. https://doi.org/10.1053/j.gastro.2019.06.039
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology and Hepatology 11: 506-514. https://doi.org/10.1038/nrgastro.2014.66
Hodgkinson, K., El Abbar, F., Dobranowski, P., Manoogian, J., Butcher, J., Figeys, D., Mack, D. and Stintzi, A., 2023. Butyrate’s role in human health and the current progress towards its clinical application to treat gastrointestinal disease. Clinical Nutrition 42: 61-75. https://doi.org/10.1016/j.clnu.2022.10.024
Hou, K., Wu, Z.X., Chen, X.Y., Wang, J.Q., Zhang, D., Xiao, C., Zhu, D., Koya, J.B., Wei, L., Li, J. and Chen, Z.S., 2022. Microbiota in health and diseases. Signal Transduction and Targeted Therapy 7: 135. https://doi.org/10.1038/s41392-022-00974-4
Hull, M.A., 2021. Conference on diet and digestive disease symposium 4: Gi cancers, the role of nutrition in prevention, pathology and management: Nutritional prevention of colorectal cancer. Proceedings of the Nutrition Society 80: 59-64. https://doi.org/10.1017/S0029665120000051
Iqbal, U., Nawaz, A., Ahmed, Z., Kamal, F., Lee-Smith, W., Khan, M.A., Alastal, Y., Confer, B.D. and Khara, H.S., 2022. Safety of endoscopic mucosal resection of large colonic polyps in elderly patients: a systematic review and meta-analysis. Annals of Gastroenterology. 35: 420-426. https://doi.org/10.20524/aog.2022.0727
Ishikawa, H., Akedo, I., Otani, T., Suzuki, T., Nakamura, T., Takeyama, I., Ishiguro, S., Miyaoka, E., Sobue, T. and Kakizoe, T., 2005. Randomized trial of dietary fiber and Lactobacillus casei administration for prevention of colorectal tumors. International Journal of Cancer 116: 762-767. https://doi.org/10.1002/ijc.2115
Ishikawa, H., Mutoh, M., Yoshimura, K., Fujii, G., Nakamura, T., Takeshita, T., Wakabayashi, K. and Sakai, T., 2020. Very long-term treatment with a lactobacillus probiotic preparation, Lactobacillus casei strain Shirota, suppresses weight loss in the elderly. Nutrients 12: 1599 https://doi.org/10.3390/nu12061599
Katanoda, K., Hori, M., Saito, E., Shibata, A., Ito, Y., Minami, T., Ikeda, S., Suzuki, T. and Matsuda, T., 2021. Updated trends in cancer in Japan: Incidence in 1985–2015 and mortality in 1958–2018 – a sign of decrease in cancer incidence. Journal of Epidemiology 31: 426-450. https://doi.org/10.2188/jea.JE20200416
Kinjo, Y., Pei, B., Bufali, S., Raju, R., Richardson, S.K., Imamura, M., Fujio, M., Wu, D., Khurana, A., Kawahara, K., Wong, C.H., Howell, A.R., Seeberger, P.H. and Kronenberg, M., 2008. Natural Sphingomonas glycolipids vary greatly in their ability to activate natural killer T cells. Chemistry and Biology 15: 654-664. https://doi.org/10.1016/j.chembiol.2008.05.012
Kocarnik, J.M., Compton, K., Dean, F.E., Fu, W., Gaw, B.L., Harvey, J.D., Henrikson, H.J., Lu, D., Pennini, A., Xu, R., Ababneh, E., Abbasi-Kangevari, M., Abbastabar, H., Abd-Elsalam, S.M., Abdoli, A., Abedi, A., Abidi, H., Abolhassani, H., Adedeji, I.A. and Force, L.M., 2022. Cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life years for 29 cancer groups from 2010 to 2019 a systematic analysis for the Global Burden of Disease Study 2019. JAMA Oncology 8: 420-444. https://doi.org/10.1001/jamaoncol.2021.6987
Kuleshov, M.V., Jones, M.R., Rouillard, A.D., Fernandez, N.F., Duan, Q., Wang, Z., Koplev, S., Jenkins, S.L., Jagodnik, K.M., Lachmann, A., McDermott, M.G., Monteiro, C.D., Gundersen, G.W. and Maayan, A., 2016. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Research 44: W90-W97. https://doi.org/10.1093/nar/gkw377
Kvakova, M., Kamlarova, A., Stofilova, J., Benetinova, V. and Bertkova, I., 2022. Probiotics and postbiotics in colorectal cancer: Prevention and complementary therapy. World Journal of Gastroenterology 28: 3370-3382. https://doi.org/10.3748/wjg.v28.i27.3370
Li, G., Lin, J., Gao, X., Su, H., Lin, R., Gao, H., Feng, Z., Wu, H., Feng, B., Zuo, K., Li, Y., Wu, W., Fang, L. and Liu, Z., 2023. Intestinal epithelial pH-sensing receptor GPR65 maintains mucosal homeostasis via regulating antimicrobial defense and restrains gut inflammation in inflammatory bowel disease. Gut Microbes 15: 2257269. https://doi.org/10.1080/19490976.2023.2257269
Liang, J.Q., Li, T., Nakatsu, G., Chen, Y.X., Yau, T.O., Chu, E., Wong, S., Szeto, C.H., Ng, S.C., Chan, F.K.L., Fang, J.Y., Sung, J.J.Y. and Yu, J., 2020. A novel faecal Lachnoclostridium marker for the non-invasive diagnosis of colorectal adenoma and cancer. Gut 69: 1248-1257. https://doi.org/10.1136/gutjnl-2019-318532
Liberzon, A., Birger, C., Thorvaldsdóttir, H., Ghandi, M., Mesirov, J.P. and Tamayo, P., 2015. The molecular signatures database hallmark gene set collection. Cell Systems 1: 417-425. https://doi.org/10.1016/j.cels.2015.12.004
Litvak, Y., Byndloss, M.X. and Bäumler, A.J., 2018. Colonocyte metabolism shapes the gut microbiota. Science 362: eaat9076. https://doi.org/10.1126/science.aat9076
Love, M.I., Huber, W. and Anders, S., 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology 15: 550. https://doi.org/10.1186/s13059-014-0550-8
Lu, Y., Chen, J., Zheng, J., Hu, G., Wang, J., Huang, C., Lou, L., Wang, X. and Zeng, Y., 2016. Mucosal adherent bacterial dysbiosis in patients with colorectal adenomas. Scientific Reports 6: 26337. https://doi.org/10.1038/srep26337
Maciver, N.J., Michalek, R.D. and Rathmell, J.C., 2013. Metabolic regulation of T lymphocytes. Annual Review of Immunology 31: 259-283. https://doi.org/10.1146/annurev-immunol-032712-095956
Makide, K., Uwamizu, A., Shinjo, Y., Ishiguro, J., Okutani, M., Inoue, A. and Aoki, J., 2014. Novel lysophosphoplipid receptors: their structure and function. Journal of Lipid Research 55: 1986-1995. https://doi.org/10.1194/jlr.R046920
Manos, J., 2022. The human microbiome in disease and pathology. Apmis 130: 690-705. https://doi.org/10.1111/apm.13225
Marie, M.A., Sanderlin, E.J., Satturwar, S., Hong, H., Lertpiriyapong, K., Donthi, D. and Yang, L.V., 2022. GPR65 (TDAG8) inhibits intestinal inflammation and colitis-associated colorectal cancer development in experimental mouse models. Biochimica et Biophysica Acta – Molecular Basis of Disease 1868: 166288. https://doi.org/10.1016/j.bbadis.2021.166288
Matsuki, T., Yahagi, K., Mori, H., Matsumoto, H., Hara, T., Tajima, S., Ogawa, E., Kodama, H., Yamamoto, K., Yamada, T., Matsumoto, S. and Kurokawa, K., 2016. A key genetic factor for fucosyllactose utilization affects infant gut microbiota development. Nature Communications 7: 1-12. https://doi.org/10.1038/ncomms11939
Matsumoto, K., Takada, T., Shimizu, K., Moriyama, K., Kawakami, K., Hirano, K., Kajimoto, O. and Nomoto, K., 2010. Effects of a probiotic fermented milk beverage containing Lactobacillus casei strain Shirota on defecation frequency, intestinal microbiota, and the intestinal environment of healthy individuals with soft stools. Journal of Bioscience and Bioengineering 110: 547-552 https://doi.org/10.1016/j.jbiosc.2010.05.016
Matsumoto, S., Hara, T., Nagaoka, M., Mike, A., Mitsuyama, K., Sako, T., Yamamoto, M., Kado, S. and Takada, T., 2009. A component of polysaccharide peptidoglycan complex on Lactobacillus induced an improvement of murine model of inflammatory bowel disease and colitis-associated cancer. Immunology 128: e170-e180. https://doi.org/10.1111/j.1365-2567.2008.02942.x
Noble, A., Durant, L., Dilke, S.M., Man, R., Martin, I., Patel, R., Hoyles, L., Pring, E.T., Latchford, A., Clark, S.K., Carding, S.R. and Knight, S.C., 2022. Altered mucosal immune-microbiota interactions in familial adenomatous polyposis. Clinical and Translational Gastroenterology 13: e00428. https://doi.org/10.14309/ctg.0000000000000428
Okumura, R., Kurakawa, T., Nakano, T., Kayama, H., Kinoshita, M., Motooka, D., Gotoh, K., Kimura, T., Kamiyama, N., Kusu, T., Ueda, Y., Wu, H., Iijima, H., Barman, S., Osawa, H., Matsuno, H., Nishimura, J., Ohba, Y., Nakamura, S., Iida, T., Yamamoto, M., Umemoto, E., Sano, K. and Takeda, K., 2016. Lypd8 promotes the segregation of flagellated microbiota and colonic epithelia. Nature 532: 117-121. https://doi.org/10.1038/nature17406
Patro, R., Duggal, G., Love, M.I., Irizarry, R.A. and Kingsford, C., 2017. Salmon provides fast and bias-aware quantification of transcript expression. Nature Methods 14: 417-419. https://doi.org/10.1038/nmeth.4197
Qu, R., Zhang, Y., Ma, Y., Zhou, X., Sun, L., Jiang, C., Zhang, Z. and Fu, W., 2023. Role of the gut microbiota and its metabolites in tumorigenesis or development of colorectal cancer. Advanced Science 10: 1-21. https://doi.org/10.1002/advs.202205563
Salvi, P.S. and Cowles, R.A., 2021. Butyrate and the intestinal epithelium: Modulation of proliferation and inflammation in homeostasis and disease. Cells 10: 1775. https://www.mdpi.com/2073-4409/10/7/1775
Schroeder, K.W., Tremaine, W.J. and Ilstrup, D.M., 1987. Coated oral 5-aminosalicylic acid therapy for mildly to moderately active ulcerative colitis. A randomized study. The New England Journal of Medicine 317: 1625-1629.
Shida, K., Sato, T., Iizuka, R., Hoshi, R., Watanabe, O., Igarashi, T., Miyazaki, K., Nanno, M. and Ishikawa, F., 2017. Daily intake of fermented milk with Lactobacillus casei strain Shirota reduces the incidence and duration of upper respiratory tract infections in healthy middle-aged office workers. European Journal of Nutrition 56: 45-53. https://doi.org/10.1007/s00394-015-1056-1
Shureiqi, I., 2022. Aspirin for colorectal cancer prevention: age matters. Cancer Prevention Research 15: 565-568. https://doi.org/10.1158/1940-6207.CAPR-22-0176
Takagi, T., Naito, Y., Kashiwagi, S., Uchiyama, K., Mizushima, K., Kamada, K., Ishikawa, T., Inoue, R., Okuda, K., Tsujimoto, Y., Ohnogi, H. and Itoh, Y., 2020. Changes in the gut microbiota are associated with hypertension, hyperlipidemia, and type 2 diabetes mellitus in Japanese subjects. Nutrients 12: 1-13. https://doi.org/10.3390/nu12102996
Tan, J., McKenzie, C., Potamitis, M., Thorburn, A.N., Mackay, C.R. and Macia, L., 2014. The role of short-chain fatty acids in health and disease. Advances in Immunology 121: 91-119. https://doi.org/10.1016/B978-0-12-800100-4.00003-9
Tanahashi, J., Daa, T., Gamachi, A., Kashima, K., Kondoh, Y., Yada, N. and Yokoyama, S., 2008. Human intestinal spirochetosis in Japan; its incidence, clinicopathologic features, and genotypic identification. Modern Pathology 21: 76-84. https://doi.org/10.1038/modpathol.3800987
Thomas, A.M., Jesus, E.C., Lopes, A., Aguiar, S., Begnami, M.D., Rocha, R.M., Carpinetti, P.A., Camargo, A.A., Hoffmann, C., Freitas, H.C., Silva, I.T., Nunes, D.N., Setubal, J.C. and Dias-Neto, E., 2016. Tissue-associated bacterial alterations in rectal carcinoma patients revealed by 16S rRNA community profiling. Frontiers in Cellular and Infection Microbiology 6: 179. https://doi.org/10.3389/fcimb.2016.00179
Tian, H., Cui, J., Ye, C., Zhao, J., Yang, B., Xu, Y., Ji, S., Wang, L., Lv, X., Ma, C., Zhou, S., Li, N., Wang, X., Qin, H. and Chen, Q., 2023. Depletion of butyrate-producing microbes of the Firmicutes predicts nonresponse to FMT therapy in patients with recurrent Clostridium difficile infection. Gut Microbes 15: 2236362. https://doi.org/10.1080/19490976.2023.2236362
Vich Vila, A., Collij, V., Sanna, S., Sinha, T., Imhann, F., Bourgonje, A.R., Mujagic, Z., Jonkers, D.M.A.E., Masclee, A.A.M., Fu, J., Kurilshikov, A., Wijmenga, C., Zhernakova, A. and Weersma, R.K., 2020. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nature Communications 11: 362. https://doi.org/10.1038/s41467-019-14177-z
Yokoyama, Y., Takachi, R., Ishihara, J., Ishii, Y., Sasazuki, S., Sawada, N., Shinozawa, Y., Tanaka, J., Kato, E., Kitamura, K., Nakamura, K. and Tsugane, S., 2016. Validity of short and long self-administered food frequency questionnaires in ranking dietary intake in middle-aged and elderly Japanese in the Japan Public Health Center-Based Prospective Study for the Next Generation (JPHC-NEXT) protocol area. Journal of Epidemiology 26: 420-432. https://doi.org/10.2188/jea.JE20150064
Yu, G., Wang, L.G., Han, Y. and He, Q.Y., 2012. ClusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 16: 284-287. https://doi.org/10.1089/omi.2011.0118






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}