Abstract
Abnormal colonisation of the ileal mucosa by adherent-invasive Escherichia coli (AIEC) is a key feature of Crohnâs disease. To date, no curative treatment for this disease exists, highlighting the need to develop new therapies targeting the origin of the inflammation, in particular the intestinal microbiota and more specifically AIEC. This study investigated the anti-virulence properties of 17 bacterial strains (lactobacilli and bifidobacteria) and three plant extracts (walnut and green tea leaves and liquorice roots) against AIEC. In vitro, six lactobacilli strains and one bifidobacterium strain reduced AIEC LF82 adhesion to Caco-2/TC7 cells and/or suppressed IL-8 secretion induced by AIEC. Although plant extracts did not prevent adhesion or inflammation, they inhibited AIEC growth. In a murine model of dextran sulfate sodium-induced colitis exacerbated by LF82 infection, two Lacticaseibacillus strains, one Bifidobacterium strain, and walnut and green tea extracts efficiently alleviated colitis and reduced faecal lipocalin-2 levels. For the green tea extract and one Lacticaseibacillus strain, beneficial effects were correlated with a decreased number of AIEC associated with the colonic mucosa. Building on these findings, bacteria and plant extract combinations were tested in the same model. A formulation combining two Lacticaseibacillus strains (Lbs. casei and Lbs. rhamnosus) with the walnut extract demonstrated the greatest efficacy, markedly reducing colitis score and preserving intestinal mucosa integrity. While untreated mice remained heavily colonised, the combination promoted AIEC elimination from the gut of half the mice, contributing to the alleviation of colitis symptoms. These results highlight the ability of combinations of specific bacteria/plant extracts to limit the presence of AIEC in the ileal mucosa of Crohnâs disease patients, presenting a promising approach for disease management.
1 Introduction
Crohnâs disease (CD) is a chronic inflammatory bowel disease (IBD) resulting from an aberrant immune response to detrimental environmental stimuli, including intestinal microbiota, in genetically predisposed individuals (Dolinger et al., 2024). Currently, no curative treatment exists in CD which remains a substantial economic and social burden on public health (Floyd et al., 2015). Therapeutic strategies, including anti-inflammatory agents, immunomodulators, biologics, and surgical resection of inflamed regions, are implemented to manage symptoms and reduce recurrences, rather than achieving a definitive cure (de Mattos et al., 2015).
The gut microbiota plays a critical role in the pathogenesis of CD. Numerous studies have demonstrated dysbiosis in the intestinal microbiota of CD patients compared to healthy individuals, notably with an increased prevalence in adherent-invasive Escherichia coli (AIEC) (Darfeuille-Michaud et al., 2004). AIEC are considered as a âCD pathobiontâ due to their ability to initiate and/or sustain the chronic inflammatory state of the disease in a susceptible host. These bacteria strongly adhere to intestinal epithelial cells (IEC) enabling their invasion (Boudeau et al., 1999; Eaves-Pyles et al., 2008) and are capable of surviving and replicating within macrophages, where they induce the secretion of tumour necrosis factor-alpha (TNF-α) (Bringer et al., 2006; Glasser et al., 2001). In a recent clinical study on CD patients, an association was observed between AIEC and early ileal lesions, with the presence of these bacteria identified as a predictive marker for endoscopic post-operative recurrence (POR) at six months (Buisson et al., 2023). This finding underscores the role of AIEC in the initial stages of ileal CD and the necessity of developing AIEC-targeted therapies as a strategy to prevent disease progression. Various strategies have been investigated to eliminate AIEC from the gut, including an anti-adhesive approach using heptylmannoside derivatives, such as FimH antagonists (Sivignon et al., 2015b) and a targeted elimination approach with bacteriophages (Galtier et al., 2017). Furthermore, probiotic interventions present a promising strategy for disrupting AIEC colonisation in the gut.
Probiotics are defined as âlive microorganisms which, when administered in adequate amounts, confer a health benefit on the hostâ (FAO/WHO, 2001). They can positively affect the host by strengthening the intestinal barrier and modulating the immune response, or they can act on pathogenic bacteria by inhibiting their adhesion to the epithelium or by creating an environment unfavourable to their survival (Alard et al., 2018; HrdyÌ et al., 2020; Ni et al., 2023; Plaza-Diaz et al., 2019). Studies targeting AIEC bacteria have shown the positive effects of Escherichia coli (E. coli) Nissle 1917, along with lactobacilli and bifidobacteria strains and certain yeasts, particularly the strain Saccharomyces cerevisiae CNCM I-3856, by significantly reducing AIEC adhesion to IEC and modulating inflammation (Boudeau et al., 2003; Ingrassia et al., 2005; Leccese et al., 2020; Sivignon et al., 2015a). In vivo, S. cerevisiae CNCM I-3856, and particularly heteropolysaccharides extracted from its cell wall, decreased AIEC gut colonisation and prevented signs of colitis in a mouse model of CD (Sivignon et al., 2021, 2015a). However, meta-analyses of clinical trials assessing the efficacy of probiotics in CD patients have not shown significant benefits (Limketkai et al., 2020). This lack of effectiveness may be due to various factors, including the choice of probiotic strains, doses, formulations, treatment duration, cohort heterogeneity, and, in particular, insufficient consideration of patientsâ specific microbial dysbiosis profiles, such as the presence of AIEC.
Alongside traditional anti-inflammatory medications, an increasing number of IBD patients are exploring alternative strategies, such as plant extracts, which represent natural products, to help alleviate symptoms. Plant extracts can modulate the intestinal microbiota by increasing the number of beneficial bacterial species producing short-chain fatty acids (SCFA) (Wu et al., 2021). Inversely, they can exert effects against pathogenic bacteria, such as E. coli strains responsible for urinary tract infections or diarrhea, primarily through their antibacterial activity and ability to inhibit biofilm formation (Lagha et al., 2019; Olivas-Quintero et al., 2022). An anhydrous ethanol extract from walnut husks showed antibacterial activity against Staphylococcus aureus, Bacillus subtilis, and E. coli via cell wall damage and impaired energy metabolism, alteration of bacterial growth, and proliferation (Wang et al., 2023). To date, few studies have demonstrated the ability of plant extracts to reduce adhesion and invasion levels of AIEC (Szandruk-Bender et al., 2020; Zhang et al., 2021).

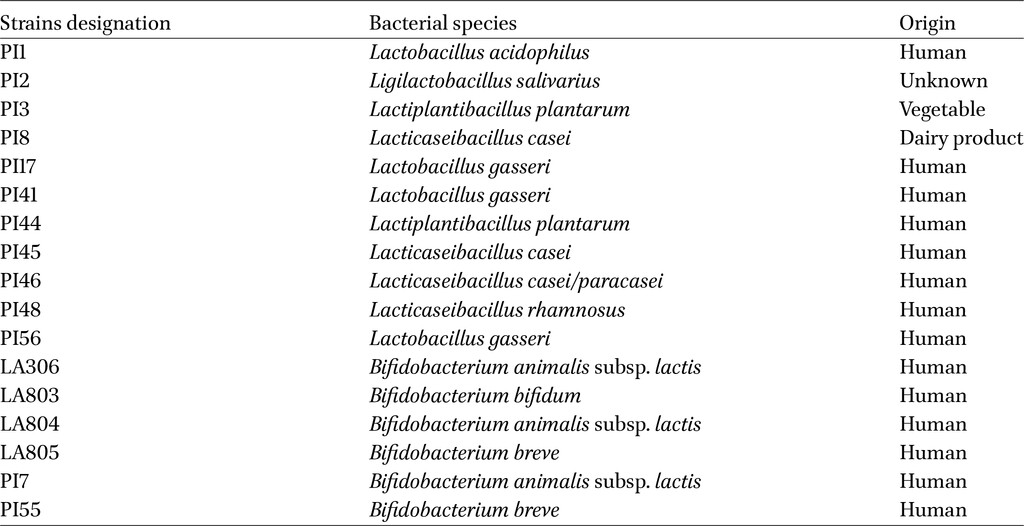
Bacterial strains studied
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
Our study aimed to perform an in vitro screening of the potential of probiotic strains and plant extracts for their anti-inflammatory and anti-virulence properties against AIEC. Bacteria and plant extracts of interest were subsequently evaluated in vivo, using a murine model of dextran sulfate sodium (DSS)-induced colitis infected with AIEC, both independently and in combination, to assess their ability to prevent AIEC gut colonisation and alleviate associated colitis symptoms. Our findings led to the development of a bacteria-plant extract-based formulation capable of limiting AIEC gut colonisation and mitigating associated colitis symptoms.
2 Material and methods
Bacterial strains and plant extracts
The Adherent-Invasive E. coli (AIEC) strain LF82, resistant to ampicillin and erythromycin, was isolated from an ileal biopsy of a Crohnâs disease (CD) patient. The LF82-
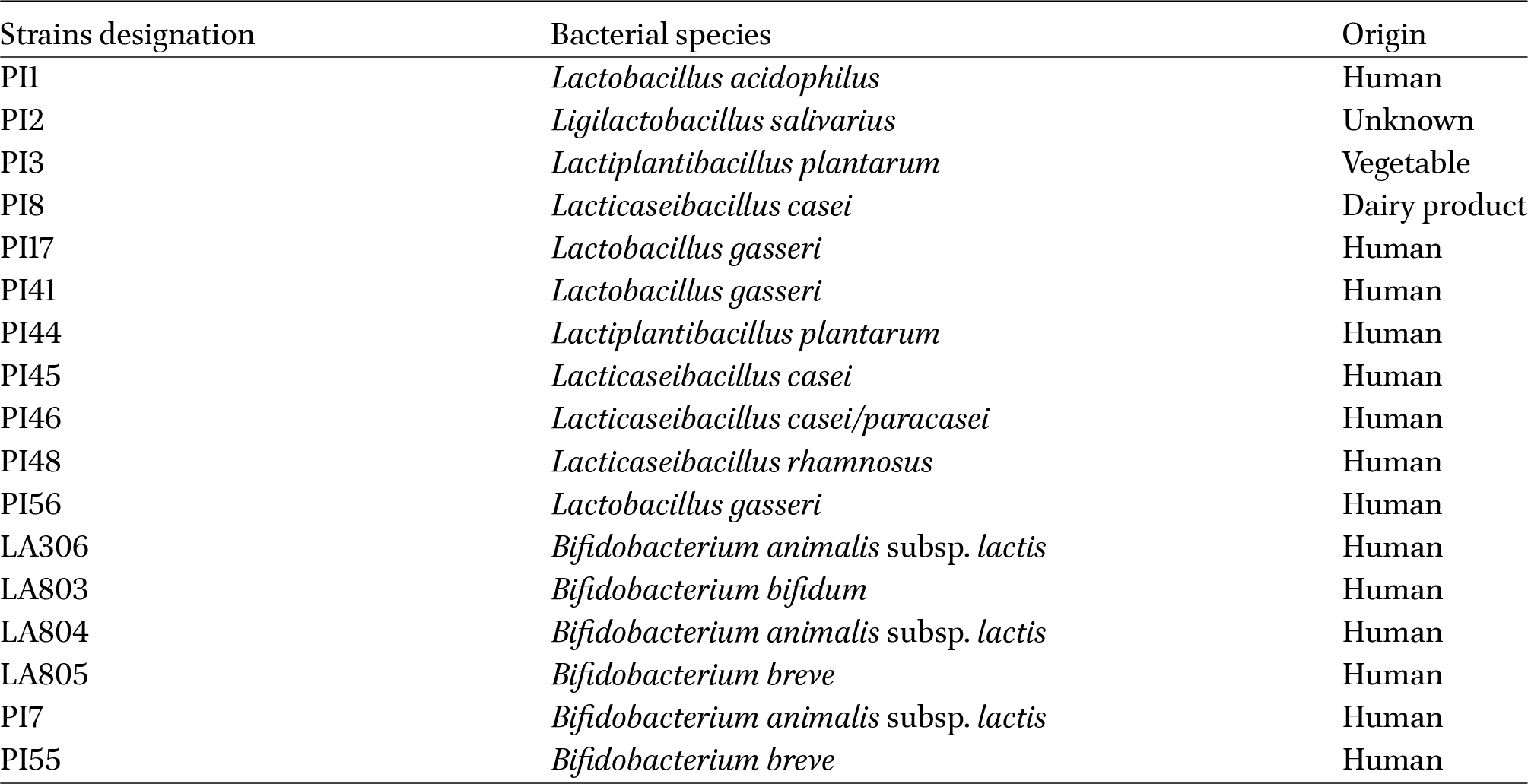
Eleven lactobacilli and six bifidobacteria strains were provided by PiLeJe Laboratoire to assess their properties (Table 1). Lactobacilli were cultured for 24 h and bifidobacteria for 48 h at 37 °C under anaerobic conditions (GENbag anaer, BiomeÌrieux, Marcy lâEÌtoile, France) in De Man, Rogosa and Sharpe broth (MRS, Biokar Diagnostics, Beauvais, France), supplemented with 0.1% L-cysteine hydrochloride (Thermo Fisher Scientific, Waltham, MA, USA) for bifidobacteria. For experimental studies, bacterial cultures were centrifuged (3,500 g, 10 min, 4 °C) and resuspended in Phosphate Buffer Sodium (PBS). For combination experiment, bacterial strains in lyophilized form (supplied by PiLeJe Laboratoire) were directly resuspended in PBS.
Plant extracts from walnut (Juglans regia L.) leaves, liquorice (Glycyrrhiza glabra L.) roots, and green tea (Camellia sinensis L.) leaves were provided by PiLeJe Laboratory in lyophilized form. The extract of C. sinensis tested (Green tea CPSP-01, named green tea extract subsequently) is obtained by extraction of 1 kg of C. sinensis dry leaves in 10 l of water at 85 °C for 30 min. After filtration, the resulting extract is concentrated under vacuum (native extract ratio [NER] about 4:1), then fixed and dried on 0.835 kg of C. sinensis dry leaves (impregnation support) under reduced pressure (drug extract ratio [DER] about 2:1). The resulting product is crushed to form the green tea extract. Dry hydroalcoholic extracts of walnut leaves and liquorice roots were prepared as described above with 20 to 70% (v/v) ethanolic leaching (Authier et al., 2022). Extracts were reconstituted in PBS and sonicated using a Bioruptor device (Diagenode, Seraing, France) for three 15-second cycles. The same batches and preparation methods were used across all experiments included in this study.
Cell culture
The human intestinal epithelial cell line Caco-2/TC7 (ATCC HTB-37) was maintained in high-glucose Dulbeccoâs Modified Eagle Medium (DMEM) supplemented with L-glutamine (Gibco, Grand Island, NY, USA), 10% fetal bovine serum (FBS) (Dutscher, Bernolsheim, France), 1% antibiotic/antimycotic solution (Cytiva, Marlborough, MA, USA), 1% non-essential amino acids (Fisher), and 1% vitamins (Dutscher). Cells were cultured at 37 °C in a humidified atmosphere containing 5% CO2. TC7 cells were seeded in 96-well plates (104 cells/well) for viability assays, in 24-well plates (4 à 105 cells/well) for adhesion assays and cytokine measurements, before incubation for an additional 24 h.
Bacterial growth
Increasing concentrations of plant extracts were incubated with bacteria in LB broth for AIEC or MRS medium (± 0.1% cysteine) for lactobacilli or bifidobacteria, and growth was monitored by measuring optical density (OD) at 600 nm every 30 min for 12 or 16 h using the Flexstation 3 instrument (Molecular Devices, San Jose, CA, USA).
Functional assessment of type 1 pili and flagella
Functionality of type 1 pili and flagella were assessed by yeast agglutination and mobility test, respectively. AIEC LF82 or LF82-
Biofilm formation assay
Biofilm formation assays were adapted from a previously described method (Martinez-Medina et al., 2009b). Briefly, strains were grown overnight with plant extracts (100 μg/ml) in LB broth supplemented with 5 g/l of glucose (Sigma-Aldrich, St. Louis, MO, USA). Then, 130 μl of a standardised LF82 suspension (3 à 107 CFU/ml) in M63 medium (0.8% glucose, 1 mM MgSO4) (Bio Basic, Toronto, Canada) were placed into microplates and grown for 24 h at 30 °C under shaking. Growth OD was read at 630 nm. Wells were washed once in PBS and adhered bacteria were stained with 1% crystal violet (Sigma-Aldrich). After five washes, crystal violet stain â bound biofilm was solubilized in ethanol and the OD was read at 570 nm. Biofilm formation index was calculated using the formula: (OD570 attached bacteria â OD570 control) / (OD630 of bacterial growth in broth â OD630 control).
Viability assay on TC7 cells
Serial dilutions of plant extracts were added to Caco-2/TC7 cells for 24Â h. After PBS washes, cell viability was assessed using the XTT assay (Biotium, Fremont, CA, USA) according to the manufacturerâs protocol. Absorbance was measured at 450Â nm and 650Â nm after 4Â h of XTT incubation.
Adhesion assays on Caco-2/TC7 cells
Caco-2/TC7 cells were infected with AIEC or DAEC bacterial strains at a multiplicity of infection (MOI) of 10 bacteria per cell for 3Â h. Candidate probiotics were tested at MOI comprised between 1 and 200 per cell. For co-incubation experiments, candidates and AIEC were added simultaneously on cells. For pre-incubation assays, cells were incubated with candidates 1-h prior to infection with AIEC or DAEC bacteria. Candidate strains were also incubated alone for 4Â h on cells to determine their adhesive capacity. Plant extracts were incubated overnight either with TC7 cells or during LF82 growth for 18Â h before cellular infection. Cells were washed with PBS, lysed with 1% Triton X-100 (Sigma-Aldrich), and spread on LB agar plates using an easySpiral (Interscience, Saint Nom la BreteÌche, France) with the appropriate antibiotics to quantify E. coli CFU or on MRS agar for lactobacilli or bifidobacteria enumeration. The number of CFU was quantified with the Scan 4000 (Interscience).
Cytokine quantification
TC7 cells were infected with AIEC strains at MOI 10 for 5Â h. Cells were incubated with candidate strains (MOI 100) for 1-h prior to AIEC infection. For plant extract studies, cells were pre-incubated overnight before LF82 infection. IL-8 and CCL-2 levels in cell culture supernatants were quantified by ELISA (#DY208 and #DY279, Duoset, R&D systems, Minneapolis, MN, USA).
Mice and ethics statement
Six-week-old BALB/cByJ male mice purchased from Charles River Laboratories (Lâarbresle, France) were housed at the University Clermont Auvergne (accredited establishment no. D63 113 16) under standard laboratory conditions (22 ± 1 °C, 12-12-h light/dark cycle). They were acclimated for one week with ad libitum access to food and water. All animal experiments were performed in accordance with the Committee for Research and Ethical Issues C2E2A and the French ministry of Education & Research (Approval No. 38082-2022071217268046). This study was carried out in accordance with relevant guidelines and regulations.
Mouse infection experiments
Mice were randomly assigned to 7 or 8 groups (n = 6 or 8 per group). To induce colitis, mice were given 2% (w/v) of dextran sulfate sodium (DSS, MW 36-50 kDa, MP Biomedicals, Eschwege, Germany) in drinking water for 8 days (D0-D8), and were orally challenged with 108 AIEC LF82 once a day during this period. Probiotics (109 CFU), plant extracts (1 mg), or combinations were administered daily by gavage in 0.2 ml PBS for one week before (D-7 to D-1) and during the DSS-LF82 challenge. Body mass was recorded daily. Disease Activity Index (DAI) score which is a composite score determined by the relative body weight loss, stool softness, and faecal blood (Hemocare test, Care Diagnostica, Voerde, Germany), was assessed to evaluate the progression of colitis (Supplementary Table S1). Faecal lipocalin-2 was measured by ELISA (#DY1857, Duoset, R&D systems). Intestinal permeability was evaluated on D7 by the fluorescence measurement (Flexstation 3, Molecular Devices) of FITC-dextran 4 kDa (FD4, TdB labs, Uppsale, Sweden) in the serum of mice 5 h after its administration by gavage of 15 mg of FD4 diluted in PBS. On D8, mice were euthanized and colons were collected for histological analyses, LF82 quantification, and cytokine measurements. Concerning histomorphological evaluation, Swiss rolls were made from longitudinally opened-colonic tissues, fixed in PBS-buffered 10% formalin, embedded in paraffin, and then sectioned into 5-μm thick slices for haematoxylin and eosin (H&E) staining. The colitis severity was scored (Supplementary Tables S2 and S3) in a blinded manner by professional pathologists using light microscopy. The final score reflects signs of inflammation, cell infiltration, fibrosis, and epithelial and mucosal damage. Quantification of LF82 associated with the colonic mucosa was carried out on a 1 cm segment of proximal colon. The pro-inflammatory cytokine KC released by the colonic tissue in culture medium for 24 h was quantified by ELISA according to the manufacturerâs instructions (#DY453, Duoset, R&D systems).
Statistical analysis
Data are presented as mean ± SEM or median. Statistical comparisons were performed using the non-parametric Kruskal-Wallis test with Dunnâs multiple comparison. Differences were considered significant at
3 Results
Lactobacilli strains decrease AIEC-induced inflammation in intestinal epithelial cells
The ability of the candidate bacterial strains to reduce cytokine release by Caco-2/TC7 cells upon AIEC infection was investigated. First, cells were pre-incubated for 1 h with bacterial strains before being infected with the AIEC reference strain LF82 for five additional hours. As shown in Figure 1A, infection with AIEC LF82 induced high IL-8 secretion by TC7 cells (300 pg/ml). Among the 17 strains tested, five significantly prevented IL-8 secretion: PI8, PI17 (
Efficacy of 17 strains of lactobacilli or bifidobacteria in reducing the inflammatory response of AIEC-infected intestinal epithelial TC7 cells. (A-D) Caco-2/TC7 cells were incubated with bacterial strains (MOI 100; lactobacilli in red; bifidobacteria in blue) for 1 h, followed by a 5-h infection with the AIEC LF82 strain (A, B) or other AIEC strains (C, D) at MOI 10. IL-8 (A, C) and CCL-2 (B, D) cytokine releases were measured in cell media by ELISA. Data are presented as mean ± SEM (n = 3). *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
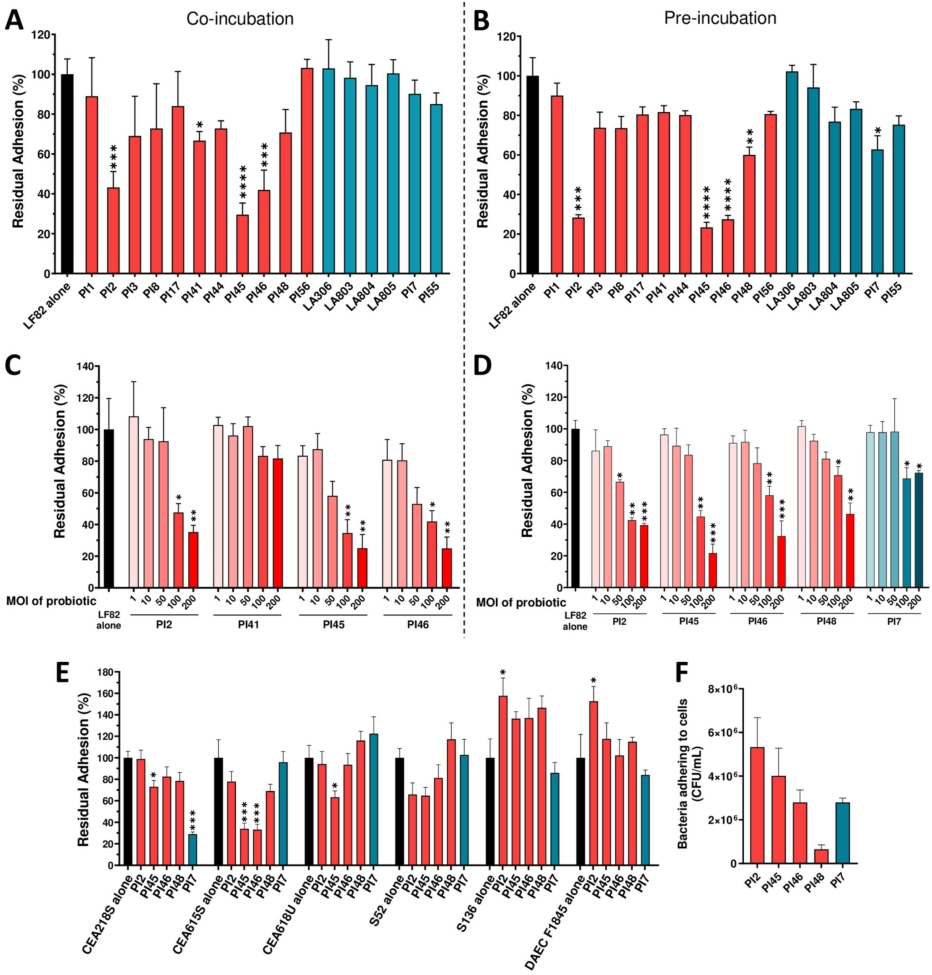
Lactobacilli strains inhibit AIEC-adhesion to intestinal epithelial cells
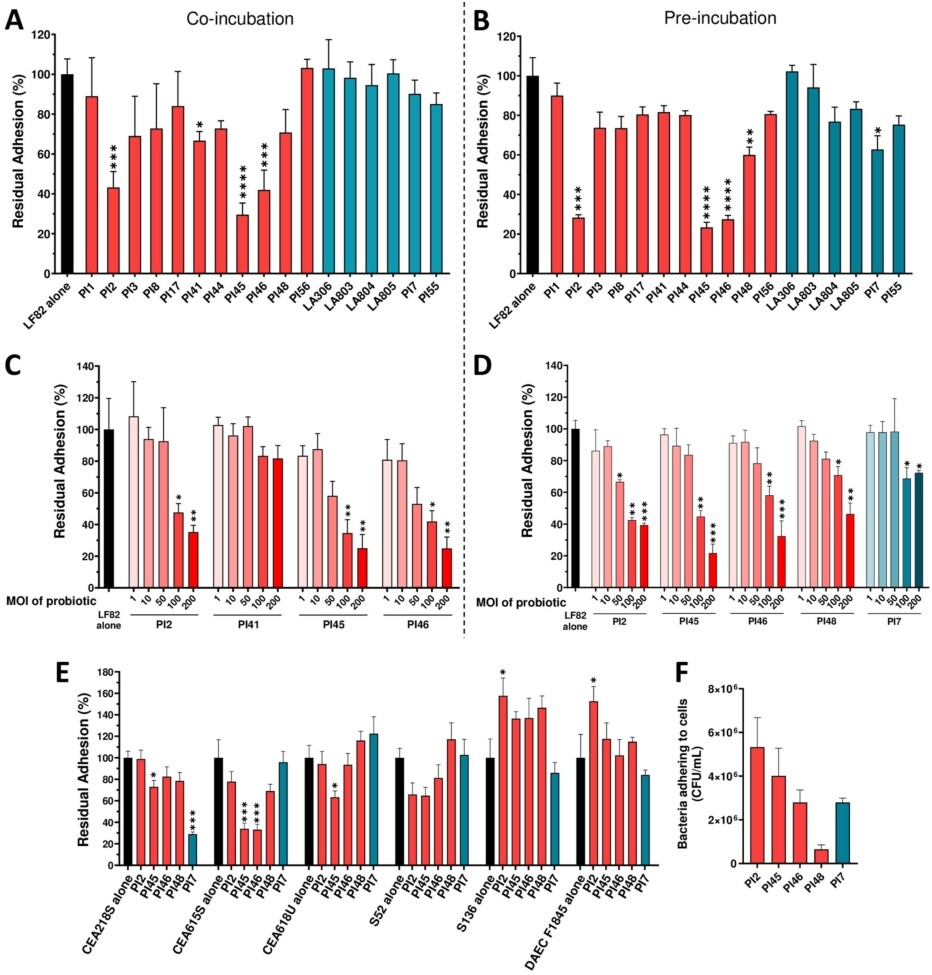
The ability of the 17 bacterial strains to inhibit adhesion of AIEC to IEC, the main virulence feature of this pathobiont, was evaluated. Four lactobacilli strains, PI41, PI2, PI46, and PI45, significantly reduced AIEC LF82 adhesion to cells (66.7% ± 4.6%;
Efficacy of 17 strains of lactobacilli and bifidobacteria in reducing AIEC strains adhesion to intestinal epithelial TC7 cells. (A-D) Caco-2/TC7 cells were infected with AIEC LF82 at MOI 10 for 3 h. (A) co-incubation experiment: TC7 cells were simultaneously incubated with probiotic strains (MOI 100) and AIEC LF82 for 3 h. (B) pre-incubation experiment: TC7 cells were first incubated with probiotic strains (MOI 100) for 1 h, followed by a 3-h infection with AIEC LF82 in the presence of probiotics. (C-D) dose-effect assays with candidate strains in either a co-incubation (C) or pre-incubation (D) setup. (E) percentages of residual adhesion of AIEC strains and one DAEC strain to intestinal epithelial cells in the presence of five candidate strains using a pre-incubation protocol. (F) TC7 cells were incubated with probiotic strains at MOI 100 for 4 h. Bacteria adhering to the cells were quantified on MRS agar plates. Results are expressed as percentages of adherent bacteria, with adhesion in the absence bacteria set at 100% (A-E), or as number of adherent bacteria per ml (F). Data are presented as mean ± SEM (n = 3-4). *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
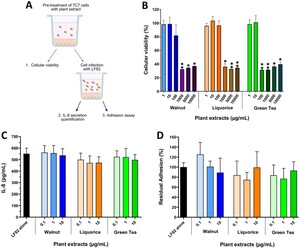
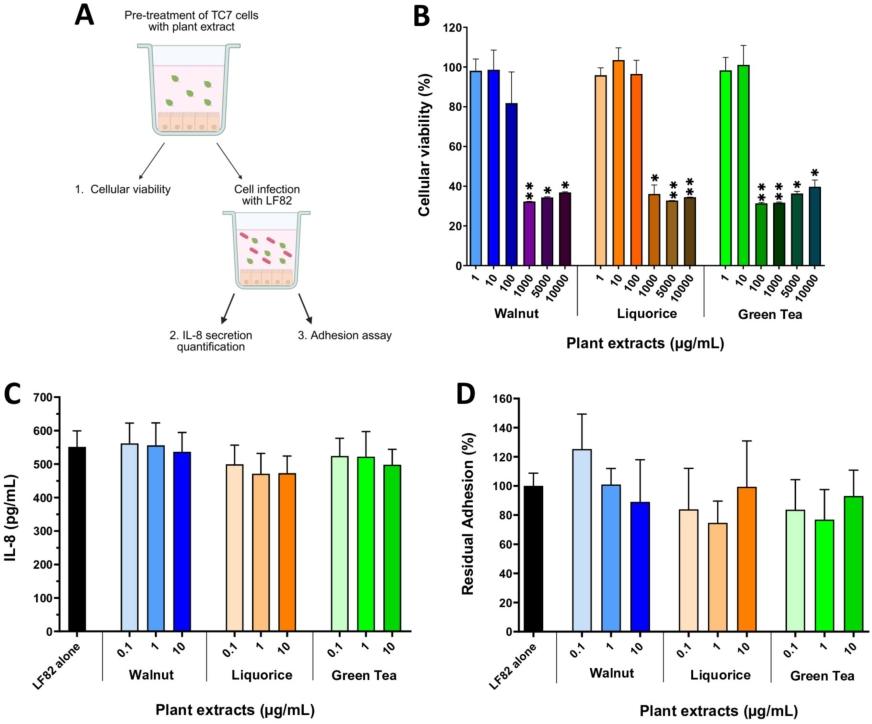
Effects of a pre-treatment of TC7 cells with plant extracts on cellular viability, AIEC-induced inflammatory response and AIEC adhesion. (A) experimental design. (B) TC7 cells were pre-incubated for 24 h with increasing concentrations of plant extracts and cellular viability was assessed by XTT assay. (C) TC7 cells were pre-incubated overnight with plant extracts and then infected with AIEC LF82 (MOI 10) for 5 h. IL-8 secretion was measured in the culture medium by ELISA. (D) cells were pre-incubated overnight with plant extracts and then infected with AIEC LF82 (MOI 10) for 3 h. The number of cell-adherent AIEC LF82 was determined by counting on LB agar plates. Results are expressed as percentages of adherent bacteria, with AIEC LF82 infection in the absence of plant extracts set as 100%. Data are presented as mean values ± SEM (n = 3-4). *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
Plant extracts do not reduce AIEC adhesion to IEC nor inflammatory response but affect AIEC growth and type-1 pili function
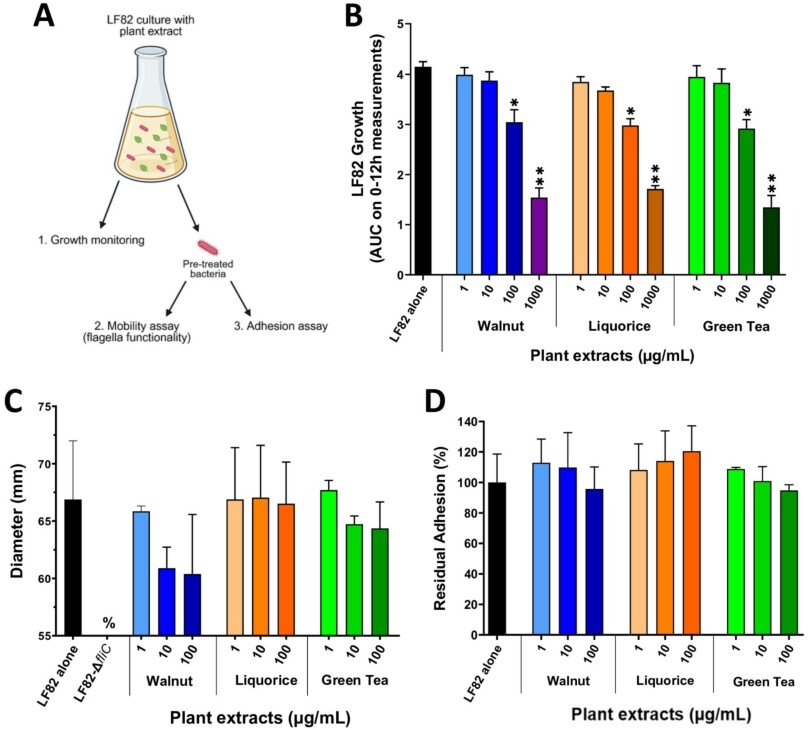
Three plant extracts â walnut (WAL), liquorice (LIQ) and green tea (GT) â were studied for their ability to interfere in the AIEC-IEC interaction. First, TC7 cells were pre-treated with increasing concentrations of extracts (Figure 3A). After 24 h of incubation, a significant reduction (â¼70%) in cell viability was observed from 1,000 μg/ml of WAL and LIQ, and from 100 μg/ml of GT (Figure 3B). As such, concentrations below 100 μg/ml of plant extracts were selected to evaluate their effects on the AIEC/cell interaction. An overnight pre-incubation of TC7 cells with plant extracts did not succeed in preventing IL-8 secretion induced by AIEC LF82 (Figure 3C) and did not modify LF82 adhesion levels to cells (Figure 3D).
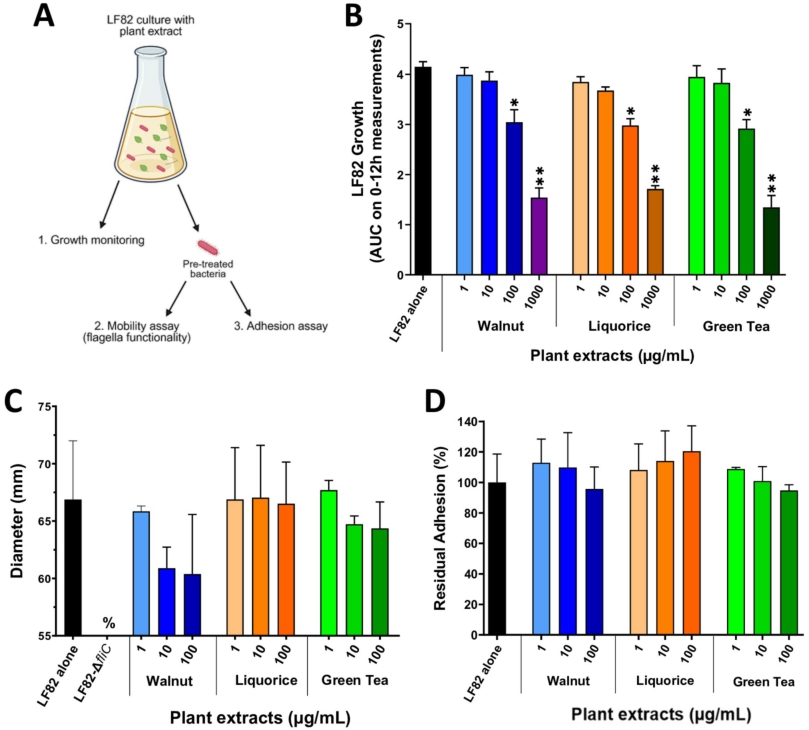
Next, LF82 bacteria were incubated with plant extracts (Figure 4A). Increasing extract concentrations (1-1,000 μg/ml) were added to a culture of AIEC LF82 in LB medium and bacterial growth was followed for 12 h. LF82 growth was significantly reduced for the three extracts starting at 100 μg/ml (Figure 4B,
Impact of a pre-treatment of AIEC LF82 with plant extracts on bacterial growth, mobility and adhesion ability. (A) experimental design. (B) growth of AIEC LF82 in LB medium alone or with varying concentrations of plant extracts was measured by OD at 600Â nm over 12Â h (area under the curve, AUC). (C) mobility assay in LB-agar 0.3% after an overnight culture of AIEC LF82 with or without plant extracts. The LF82-
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
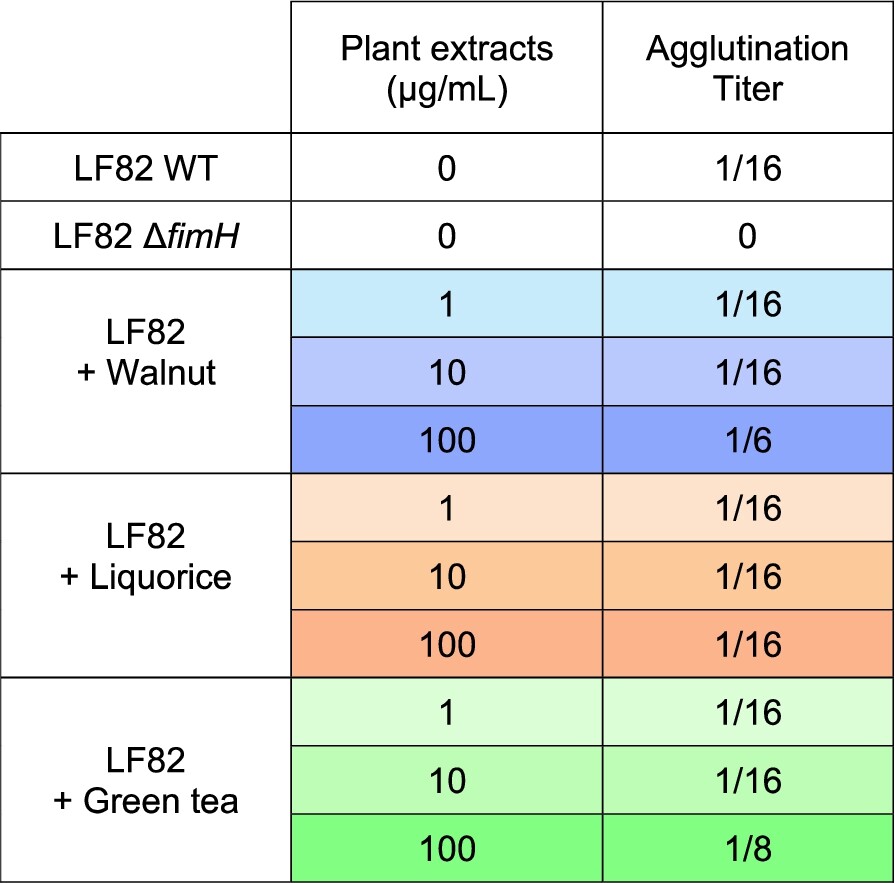
Impact of plant extracts on the ability of AIEC LF82 to produce functional type 1 pili. An agglutination assay with yeasts was performed after an overnight culture of AIEC LF82 in the presence or absence of plant extracts. The AIEC LF82-
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
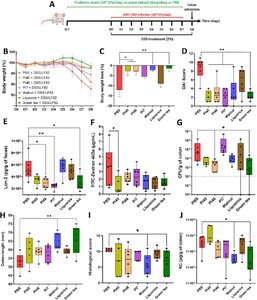
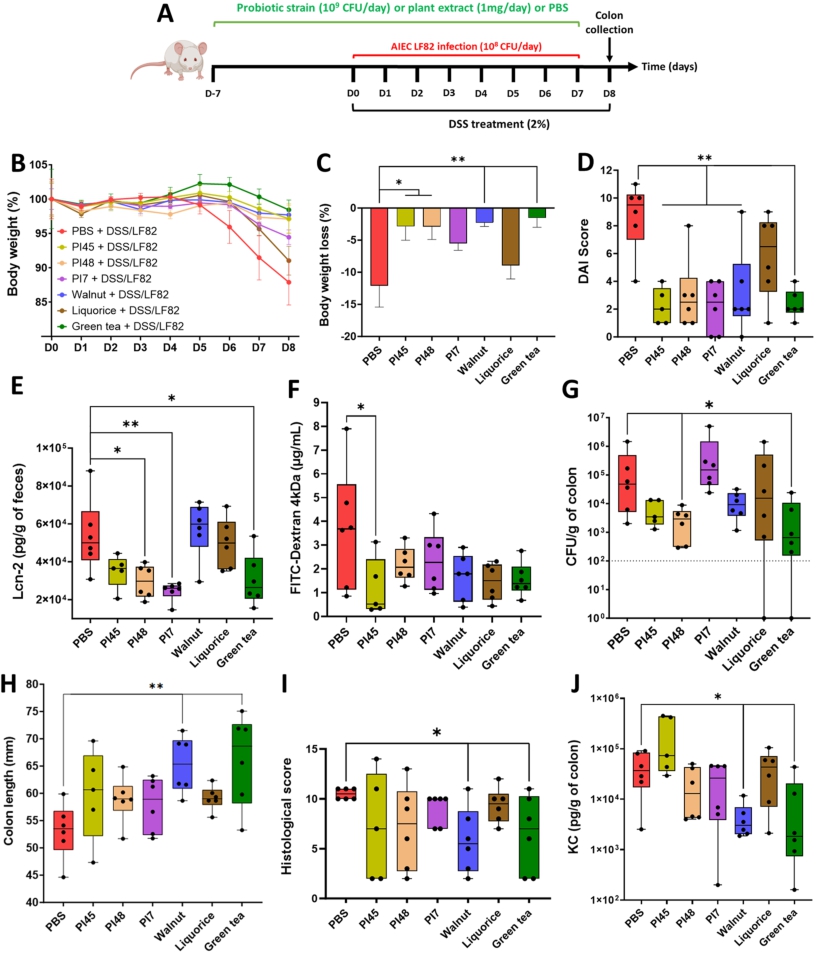
Selected bacterial strains and plant extracts significantly alleviate colitis symptoms in a DSS-induced colitis mouse model infected with AIEC LF82
Three strains, PI45, PI48 and PI7, were selected for in vivo testing, based on their ability to counteract AIEC adhesion and to reduce the associated inflammatory response. WAL, LIQ and GT extracts were also included in this assay to evaluate their global impact on the host. They were studied individually in a mouse model of colitis induced by the administration of DSS combined with a daily AIEC challenge. Candidate strains and plant extracts were administered daily by gavage (109 CFU/day or 1 mg/day) for one week in a prophylactic manner (D-7 to D-1) and continued throughout the DSS/AIEC LF82 challenge (from D0 to D8) (Figure 5A). The administration of bacteria or plant extracts alone did not induce any signs of inflammation and are well tolerated in vivo (data not shown). Body weight loss was observed for the untreated group starting from D5 and reaching 12.1% ± 8.1% at D8. PI45, PI48, WAL and GT effectively prevented this loss (Figure 5B, 5C). By D6, all treatments except LIQ significantly reduced the Disease Activity Index (DAI) score, which reflects body weight loss, stool consistency and the presence of blood in faeces (Figure 5D,
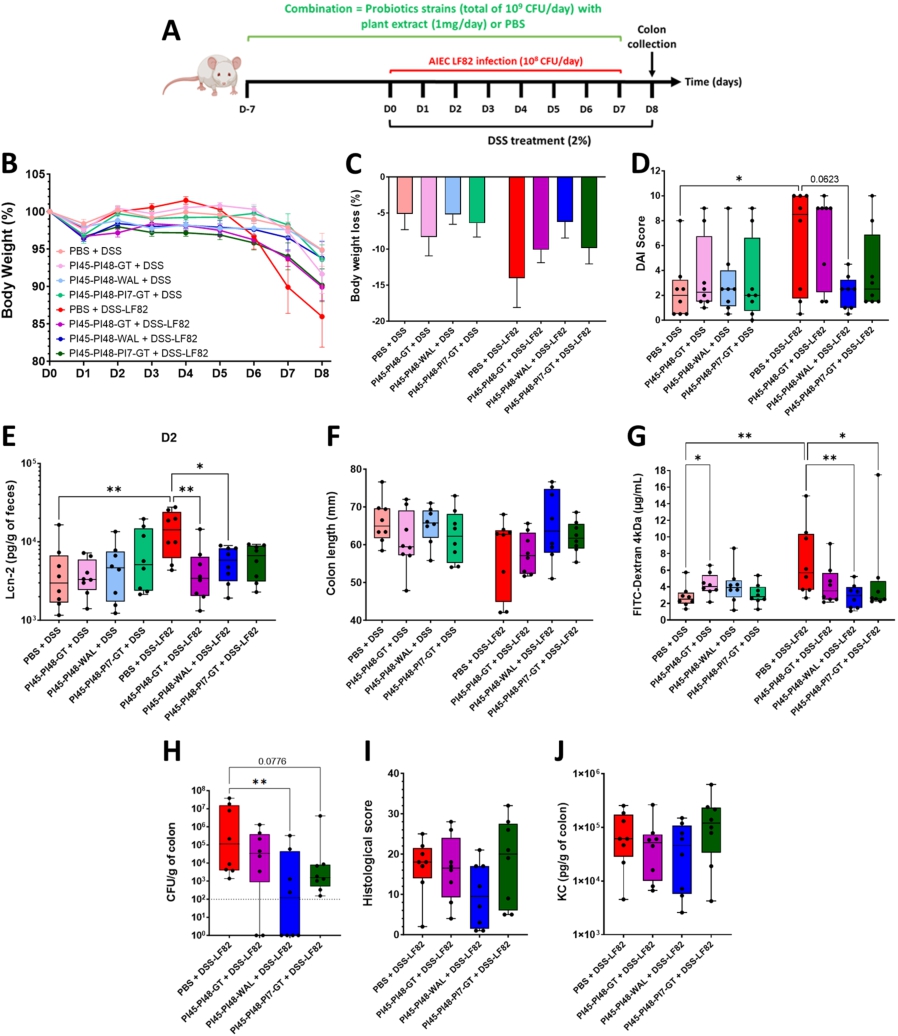
A combination of two lactobacilli and walnut extract significantly mitigates DSS-induced colitis in mice infected with AIEC bacteria
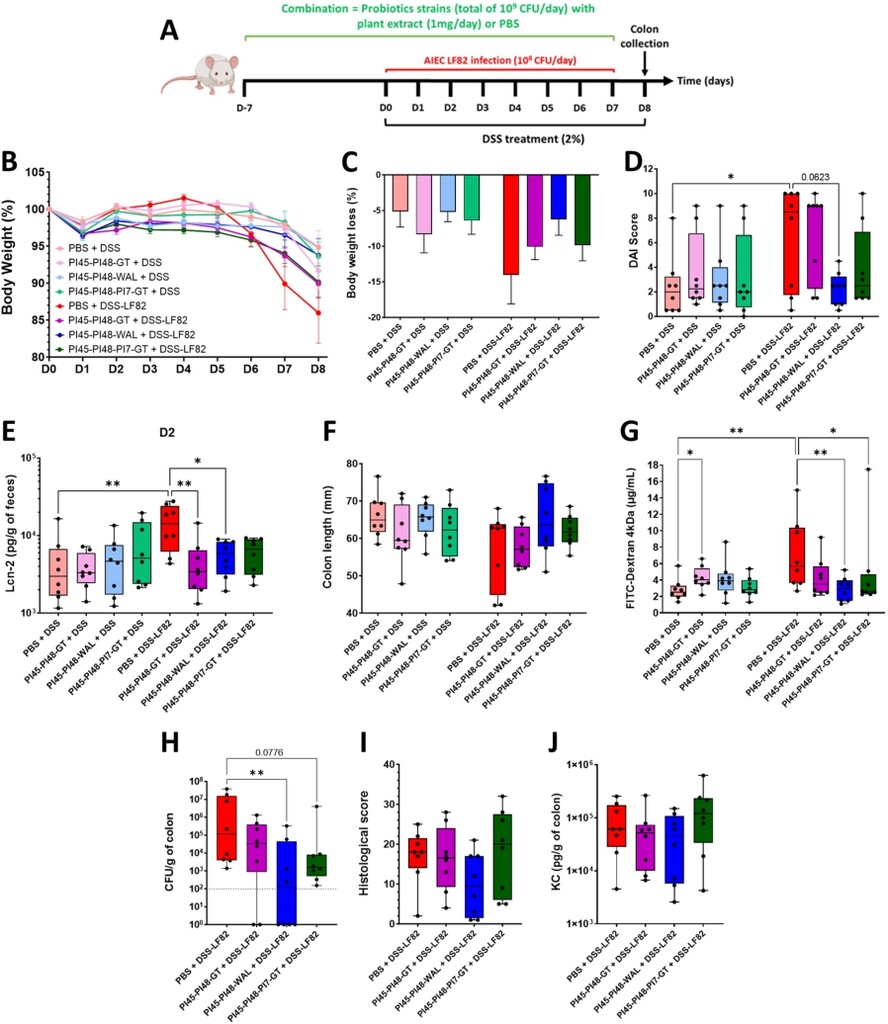
Three combinations of bacterial strains and plant extracts were tested in the DSS-LF82 mouse model to evaluate the potential for synergistic effects (Figure 6A). All combinations included the PI45 and PI48 strains, with either GT or WAL, or with strain PI7 and GT. Plant extracts showed minimal toxicity towards probiotic candidates, even after 16 h of exposure (Supplementary Figure S5), confirming that bacteria/plant formulations do not compromise bacterial viability. Body weight loss was more pronounced in AIEC-infected mice (DSS-LF82) (â14.0% ± 4.1% at D8) compared to non-infected controls (DSS-PBS) (â5.1% ± 2.2% at D8). The three combinations limited body weight loss induced by DSS-LF82 treatment, especially PI45-PI48-WAL, with an average loss of 6.2% ± 6.3% at D8 (Figure 6B, 6C). Whereas AIEC LF82 significantly exacerbated colitis severity (DSS compared to DSS+LF82,
Beneficial effects of bacterial strains and plant extracts in DSS-induced colitis aggravated by AIEC LF82 infection in mice. (A) BALB/c mice received 2% DSS in their drinking water for 8 days and were infected daily with 108 CFU of AIEC LF82 bacteria by gavage from DO to D7. Bacteria or plant extracts were administrated daily by gavage in a prophylactic manner for one week and continued during the DSS/LF82 challenge (B) evolution of body weight according to the treatments (baseline set at 100% on DO). (C) body weight loss at D8. (D) Disease Activity Index (DAI) score assessed on D6. (E) faecal Lcn-2 (lipocalin-2) concentrations measured by ELISA on D2. (F) intestinal permeability was assessed on D7 by measuring serum fluorescence 5 h after intragastric administration of FITC-dextran 4 kDa (FD4). (G) quantification of AIEC LF82 associated with the colonic mucosa on D8. (H) colon length measurements on D8. (I) histopathological scores determined on colonic tissue sections on D8, as defined in Supplementary Table S2. (J) KC cytokine released by the proximal colon measured by ELISA on D8. Data are presented as mean values ± SEM (n = 6; 5B and 5C) or as median (min to max; n = 6). *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
Effects of different bacteria and plant extracts combinations in a mouse model of DSS-induced colitis aggravated by AIEC LF82 infection. (A) BALB/c mice received 2% DSS in their drinking water for eight days and were infected daily with 108 CFU of AIEC LF82 bacteria by gavage from DO to D7. Treatment combinations were administrated daily by gavage in a prophylactic manner for 1 week and continued during the DSS/LF82 challenge. (B) evolution of body weight according to the treatments (baseline set at 100% on DO). (C) body weight loss on D8. (D) Disease Activity Index (DAI) score assessed on D6. (E) faecal Lcn-2 (lipocalin-2) concentrations measured by ELISA on D2. (F) colon length measurements on D8. (G) intestinal permeability on D7 assessed by measuring serum fluorescence 5 h after intragastric administration of FITC-dextran 4 kDa (FD4). (H) quantification of LF82 associated with the colonic mucosa on D8. (I) histopathological scores determined on colonic tissue sections on D8, as defined in Supplementary Table S3. (J) KC cytokine released by the proximal colon measured by ELISA on D8. Data are presented as mean values ± SEM (n = 8; 6B, 6C) or as median (min to max; n = 8). *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00095
4 Discussion
The aim of our study was to evaluate 17 bacterial strains belonging to the Lactobacillaceae and Bifidobacteriaceae families, along with three plant extracts, through complementary in vitro and in vivo experiments, to identify a bacteria-plant combination capable of limiting the colonisation of the AIEC pathobiont in the context of CD.
Two main criteria were considered during the screening process. The first focused on the anti-inflammatory potential of the candidate strains, assessed through the modulation of two key pro-inflammatory cytokines, IL-8 and CCL-2. Both cytokines are elevated in patients with IBD (Lu et al., 2023; Singh et al., 2016) and were consistently induced by AIEC LF82 in TC7 cells under our experimental conditions. In contrast, other pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-1β were not significantly induced by LF82 in this model. Among the strains tested, lactobacilli PI3, PI8, PI17, PI45, PI46 and PI48 efficiently decreased LF82-induced cytokine production. We observed heterogeneity in the anti-inflammatory responses among the three Lactobacillus gasseri strains (PI17, PI41, and PI56), demonstrating a strain-specific effect. Previous studies have demonstrated the anti-inflammatory potential of lactobacilli in various inflammatory and infectious contexts, particularly Lacticaseibacillus (Lbs) casei and Ligilactobacillus (Lgb) salivarius strains which alleviate inflammation by inhibiting the NF-κB pathway in cells infected with Shigella flexneri or enterotoxigenic E. coli (Qiao et al., 2020; Tien et al., 2006). In our study, two Lbs casei strains, PI45 and PI8, efficiently prevented LF82-induced inflammation, whereas the Lgb salivarius strain PI2 failed to inhibit the pro-inflammatory response and elicited an inflammatory response alone. This finding highlights the strain-specific effects of bacteria depending on the biological context.
In the second step of the bacterial strain screening, the anti-adhesive properties of the strains were assessed by incubating them with the epithelial cells during LF82 infection. Five lactobacillus strains (PI2, PI41, PI45, PI46, and PI48) and one bifidobacterium strain (PI7) demonstrated the ability to reduce LF82 adhesion.
Three strains, PI45, PI46, and PI48 combined both anti-inflammatory and anti-adhesive activities. To complete the analysis, the most effective anti-adhesive and/or anti-inflammatory strains against LF82 were tested on additional AIEC strains. Indeed, although E. coli strains classified as AIEC pathobionts share certain phenotypic traits, they are phylogenetically diverse, and the genetic factors and mechanisms underlying their pathogenicity remain incompletely understood (Bonet-Rossinyol et al., 2023; Martinez-Medina et al., 2009a). PI45, PI46, and PI48 confirmed their strong anti-inflammatory activity across different AIEC isolates. In adhesion assays, PI45 was the most effective in reducing bacterial adhesion across the AIEC panel, while PI7 markedly decreased the adhesion of the CEA218S strain. Thus, among the 17 candidate strains, PI45 (Lbs. casei), PI48 (Lbs. rhamnosus) and PI7 (Bifidobacterium animalis subsp. lactis) were selected for in vivo study. PI46 belongs to the same species as PI45 and was excluded due to its lower efficacy in inhibiting the adhesion of other AIEC strains relative to PI45. PI2 was excluded due to its pro-inflammatory effect.
In parallel, we investigated the mechanisms by which bacteria counteract AIEC adhesion to IEC. In a previous study, we demonstrated that the anti-adhesive effect of the yeast strain S. cerevisiae CNCM I-3856 against AIEC LF82 was mainly attributed to its ability to block type 1 pili from bacteria (Sivignon et al., 2015a). Indeed, AIEC bacteria primarily adhere to IEC through the mannose-binding adhesin FimH located at the tip of their type 1 pili (Boudeau et al., 2001; Dreux et al., 2013). In our study, none of the bacteria with anti-adhesive properties interfered with the FimHÂ â mannose interaction in agglutination assays, indicating that these bacteria neither block the FimH adhesin nor its mannosylated binding sites. Anti-adhesive strains may also inhibit AIEC adhesion through a non-specific manner, such as competitive adhesion to host cells. The tested bacterial strains displayed varying levels of adhesion to IEC. Lactobacillus strains PI2, PI45 and PI46 adhered strongly to TC7 cells compared to PI48, and this correlated with their higher ability to inhibit LF82 adhesion. However, no reduction in adhesion was observed against the AIEC strain S136 for these three strains, suggesting that the anti-adhesive effect may not be solely explained by physical obstruction of pathogen access to host cells. Additional experiments are required to further investigate the mechanism underlying this anti-adhesive effect.
Concerning plant extracts, none of the three compounds exhibited anti-inflammatory or anti-adhesive effects when applied during infection of IEC by AIEC bacteria. Although type 1 pili functionality is slightly affected following LF82 growth in the presence of walnut (WAL) and green tea (GT), this had no measurable impact on LF82 adhesion to IEC. Moreover, biofilm formation, another key virulence factor of the AIEC pathobiont, was not affected when bacteria were grown in the presence of the plant extracts. Interestingly, the three plant extracts affected the growth of LF82. These plant extracts have previously demonstrated bactericidal and fungicidal activity against a variety of pathogens (Falcinelli et al., 2017; Irani et al., 2010; ZÌurek et al., 2023). For instance, alcohol soluble extracts from walnut leaves at a concentration of 10Â mg/ml showed bactericidal effects against six Gram-positive and Gram-negative bacteria, and the yeast Candida albicans (ZÌurek et al., 2023). Thus, the three extracts were selected for the in vivo study, as none of the in vitro data enabled their distinction.
PI45, PI48, PI7 and plant extracts were individually evaluated in a mouse model of colitis challenged with AIEC bacteria. Given the enhanced anti-adhesive effect of the bacterial candidates in pre-incubation experiments, bacteria and extracts were administered prophylactically for seven days before the DSS/LF82 challenge. Globally, each compound, with the exception of liquorice extract (LIQ), demonstrated beneficial effects on the clinical signs of colitis and protected the intestinal mucosa from injury. The two Lbs. strains reduced the number of AIEC bacteria associated with the colonic mucosa, supporting anti-adhesive properties observed in vitro. Strain PI48 has previously been reported to alleviate colonic inflammation and to prevent increase of intestinal permeability in a murine model of low-grade inflammation induced by dinitrobenzene sulfonic acid (DNBS) (Torres-Maravilla et al., 2024). In our study, PI48 efficiently prevented early signs of intestinal inflammation, as evidence by a decreased faecal lipocalin-2 concentration on day two. However, it failed to exert a significant effect on KC release by colonic tissues at the end of experiment. The PI7 strain showed a similar profile to PI48 but the moderate anti-adhesive effect of PI7 observed in vitro did not translate into reduced LF82 colonisation in vivo. Daily administration of WAL and GT to mice significantly reduced AIEC adhesion to the intestinal mucosa although these compounds did not affect LF82 adhesion or biofilm formation in vitro. This effect may result from a direct impact on LF82 viability as observed in vitro. Additionally, it is plausible that these plant extracts modulate the gut microbiota, as green tea polyphenols and walnut peptides have been shown to promote the growth of short-chain fatty acid-producing bacterial species in mouse models of colitis (Wu et al., 2021; Zhi et al., 2022).
Three combinations were tested in mice. PI48 and PI45 were paired for their beneficial effects: both decreased colitis symptoms and LF82 colonisation, with PI45 additionally preserving intestinal barrier integrity. These strains were then combined with either WAL or GT, which showed multiple in vivo benefits. LIQ was excluded due to its lack of efficacy. A third combination, including the four candidates that prevented the early rise in faecal lipocalin-2 (PI45, PI48, PI7, and GT), was also evaluated. While the administration of these combinations to DSS-treated mice was not harmful, it did not alleviate colitis symptoms in absence of AIEC. As previously reported, LF82 exacerbated DSS-induced colitis (Carvalho et al., 2008) and despite the heterogeneity of the DAI scores, the three mice with the highest scores were those most colonised by AIEC. In LF82-infected groups, the PI45-PI48-WAL combination was as effective as each compound individually tested, in alleviating signs of colitis, in protecting intestinal barrier and in preventing the early increase of lipocalin-2. However, the beneficial effect of WAL on KC release was lost when associated with PI45 and PI48. On the other hand, this combination was greater than the individual compounds to reduce the AIEC gut colonisation, with half of the mice showing bacterial levels below the detection threshold. This may result from the combined anti-adhesive properties of the two bacteria, along with either a direct effect on AIEC viability or an indirect influence on the gut microbiota by WAL. Traditionally used in medicine, walnut leaves are rich in tannins, polyphenols, and flavonoids and are renowned for their anti-diabetic, anti-diarrheal, and antimicrobial properties (Altemimi et al., 2023; Zhi et al., 2022). Additionally, in a mouse model of candidiasis, walnut leaf extract significantly reduced gut colonisation by Candida albicans, alleviated colonic inflammation, and increased the abundance of anti-inflammatory bacterial species (Authier et al., 2022). Lastly, the two combinations containing GT were less effective in mice than the individual components, suggesting that GT may interfere with the beneficial properties of the bacteria. Additional studies are required to fully elucidate the mechanisms by which the bacteria â GT interaction may interfere with the beneficial effects of each component.
In conclusion, we screened 17 bacteria and three plant extracts for their anti-AIEC effects using complementary in vitro and in vivo approaches and assessed the benefits of their varying combinations. A formulation composed of two bacterial strains (PI45 and PI48) and walnut leaf extract was shown to reduce AIEC gut colonisation, alleviate AIEC-induced inflammation, and preserve intestinal barrier integrity. Although further investigations are needed to fully elucidate the underlying mechanisms, this combination may offer a promising therapeutic strategy for CD patients who harbour AIEC or are at risk of being colonised by AIEC, by potentially extending remission duration or improving clinical symptoms by reducing inflammatory markers and preserving or restoring the intestinal epithelium.
Corresponding author; e-mail:Â adeline.sivignon@uca.fr
We dedicate this article to Prof. Nicolas BARNICH, who sadly passed away before the publication of this work
Acknowledgements
We would like to thank Laetitia Prival from animal facility (Plateforme MutualiseÌe des Animaleries) and the CICS platform (UniversiteÌ Clermont Auvergne) for technical assistance with tissue preparation for histological analysis. We would like to thank Pierre Cordier and JeÌroÌme Abadie from Oniris (Nantes, France) for histological examinations. We would like to thank Sibylle ColombieÌ and ZoeÌ Oblette for technical help and Devon Kavanaugh for English editing. We would like to thank Vegepolys Valley for help in finding funding. We thank Claude Blondeau (PiLeJe Laboratoire) for proofreading the manuscript. We thank AureÌlie Wauquiez, CeÌsar Cotte, Loïc Lenoir and ValeÌrie Bardot for preparing the plant extracts and freeze-dried bacterial samples.
Authorsâ contribution
Conceptualisation, NB (N. Barnich) and AS; formal analysis, FDC and CL; funding acquisition, NB (N. Boisseau), SH, NB (N. Barnich) and AS; investigation, FDC and CL; methodology, FDC and AS; project administration, NB (N. Boisseau), NB (N. Barnich) and AS; supervision, NB (N. Boisseau), NB (N. Barnich) and AS; validation, NB (N. Boisseau), SH, AS and NB (N. Barnich); visualisation, FDC and AS; writing-original draft, FDC, NB (N. Boisseau) and AS; writing-review & editing, FDC, NB (N. Boisseau), CL, JD, SH and AS.
Conflict of interest
SH. is an employee of PiLeJe Laboratoire. The other authors report there are no conflict of interest to declare.
Funding
This work was supported by the ReÌgion ARAÂ â Pack Ambition Recherche 2021Â â Projet TargetBIOT. This work was also supported by the MinisteÌre de lâEnseignement SupeÌrieur, de la Recherche et de lâInnovation (MESRI), Inserm (Institut national de la santeÌ et de la recherche meÌdicale; UMR1071), INRAE (Institut national de recherche pour lâagriculture, lâalimentation et lâenvironnement; USC 1382), and by the French government IDEX-ISITE initiative 16-IDEX-0001 (CAP 20-25), I-SITE project of the University of Clermont Auvergne (CIR3). This work was also supported by PiLeJe Industry.
References
Alard, J., Peucelle, V., Boutillier, D., Breton, J., Kuylle, S., Pot, B., Holowacz, S. and Grangette, C., 2018. New probiotic strains for inflammatory bowel disease management identified by combining in vitro and in vivo approaches. Beneficial Microbes 9: 317-331. https://doi.org/10.3920/BM2017.0097
Altemimi, A.B., Al-Haliem, S.M., Alkanan, Z.T., Mohammed, M.J., Hesarinejad, M.A., Najm, M.A.A., Bouymajane, A., Cacciola, F. and Abedelmaksoud, T.G., 2023. Exploring the phenolic profile, antibacterial, and antioxidant properties of walnut leaves (Juglans regia L.). Food Sciences and Nutrition 11: 6845-6853. https://doi.org/10.1002/fsn3.3554
Authier, H., Bardot, V., Berthomier, L., Bertrand, B., Blondeau, C., Holowacz, S. and Coste, A., 2022. Synergistic effects of licorice root and walnut leaf extracts on gastrointestinal candidiasis, inflammation and gut microbiota composition in mice. Microbiology Spectrum 10: e0235521. https://doi.org/10.1128/spectrum.02355-21
Bonet-Rossinyol, Q., CamprubıÌ-Font, C., LoÌpez-Siles, M. and Martinez-Medina, M., 2023. Identification of differences in gene expression implicated in the adherent-invasive Escherichia coli phenotype during in vitro infection of intestinal epithelial cells. Frontiers in Cellular and Infection Microbiology 13: 1228159. https://doi.org/10.3389/fcimb.2023.1228159
Boudeau, J., Barnich, N. and Darfeuille-Michaud, A., 2001. Type 1 pili-mediated adherence of Escherichia coli strain LF82 isolated from Crohnâs disease is involved in bacterial invasion of intestinal epithelial cells. Molecular Microbiology 39: 1272-1284. https://doi.org/10.1111/j.1365-2958.2001.02315.x
Boudeau, J., Glasser, A.-L., Julien, S., Colombel, J.-F. and Darfeuille-Michaud, A., 2003. Inhibitory effect of probiotic Escherichia coli strain Nissle 1917 on adhesion to and invasion of intestinal epithelial cells by adherent-invasive E. coli strains isolated from patients with Crohnâs disease. Alimentary Pharmacology & Therapeutics 18: 45-56. https://doi.org/10.1046/j.1365-2036.2003.01638.x
Boudeau, J., Glasser, A.L., Masseret, E., Joly, B. and Darfeuille-Michaud, A., 1999. Invasive ability of an Escherichia coli strain isolated from the ileal mucosa of a patient with Crohnâs disease. Infection and Immunity 67: 4499-4509. https://doi.org/10.1128/IAI.67.9.4499-4509.1999
Bringer, M.-A., Glasser, A.-L., Tung, C.-H., MeÌresse, S. and Darfeuille-Michaud, A., 2006. The Crohnâs disease-associated adherent-invasive Escherichia coli strain LF82 replicates in mature phagolysosomes within J774 macrophages. Cellular Microbiology 8: 471-484. https://doi.org/10.1111/j.1462-5822.2005.00639.x
Buisson, A., Sokol, H., Hammoudi, N., Nancey, S., Treton, X., Nachury, M., Fumery, M., HeÌbuterne, X., Rodrigues, M., Hugot, J.-P., Boschetti, G., Stefanescu, C., Wils, P., Seksik, P., Le Bourhis, L., Bezault, M., Sauvanet, P., Pereira, B., Allez, M., Barnich, N. and Remind study group, 2023. Role of adherent and invasive Escherichia coli in Crohnâs disease: lessons from the postoperative recurrence model. Gut 72: 39-48. https://doi.org/10.1136/gutjnl-2021-325971
Carvalho, F.A., Barnich, N., Sauvanet, P., Darcha, C., Gelot, A. and Darfeuille-Michaud, A., 2008. Crohnâs disease-associated Escherichia coli LF82 aggravates colitis in injured mouse colon via signaling by flagellin. Inflammatory Bowel Diseases 14: 1051-1060. https://doi.org/10.1002/ibd.20423
Darfeuille-Michaud, A., Boudeau, J., Bulois, P., Neut, C., Glasser, A.-L., Barnich, N., Bringer, M.-A., Swidsinski, A., Beaugerie, L. and Colombel, J.-F., 2004. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohnâs disease. Gastroenterology 127: 412-421. https://doi.org/10.1053/j.gastro.2004.04.061
de Mattos, B.R.R., Garcia, M.P.G., Nogueira, J.B., Paiatto, L.N., Albuquerque, C.G., Souza, C.L., Fernandes, L.G.R., Tamashiro, W.M.S.C. and Simioni, P.U., 2015. Inflammatory bowel disease: an overview of immune mechanisms and biological treatments. Mediators of Inflammation 2015: 493012. https://doi.org/10.1155/2015/493012
Dolinger, M., Torres, J. and Vermeire, S., 2024. Crohnâs disease. The Lancet 403: 1177-1191. https://doi.org/10.1016/S0140-6736(23)02586-2
Dreux, N., Denizot, J., Martinez-Medina, M., Mellmann, A., Billig, M., Kisiela, D., Chattopadhyay, S., Sokurenko, E., Neut, C., Gower-Rousseau, C., Colombel, J.-F., Bonnet, R., Darfeuille-Michaud, A. and Barnich, N., 2013. Point mutations in FimH adhesin of Crohnâs disease-associated adherent-invasive Escherichia coli enhance intestinal inflammatory response. PLoS Pathogens 9: e1003141. https://doi.org/10.1371/journal.ppat.1003141
Eaves-Pyles, T., Allen, C.A., Taormina, J., Swidsinski, A., Tutt, C.B., Eric Jezek, G., Islas-Islas, M. and Torres, A.G., 2008. Escherichia coli isolated from a Crohnâs disease patient adheres, invades, and induces inflammatory responses in polarized intestinal epithelial cells. International Journal of Medical Microbiology 298: 397-409. https://doi.org/10.1016/j.ijmm.2007.05.011
Falcinelli, S.D., Shi, M.C., Friedlander, A.M. and Chua, J., 2017. Green tea and epigallocatechin-3-gallate are bactericidal against Bacillus anthracis. FEMS Microbiology Letters 364. https://doi.org/10.1093/femsle/fnx127
FAO/WHO, 2001. Evaluation of health and nutritional properties of probiotics in food, including powder milk with live lactic acid bacteria. Food and Agriculture Organization of the United Nations and World Health Organization Expert Consultation Report.
Floyd, D.N., Langham, S., SeÌverac, H.C. and Levesque, B.G., 2015. The economic and quality-of-life burden of Crohnâs disease in Europe and the United States, 2000 to 2013: a systematic review. Digestive Diseases and Sciences 60: 299-312. https://doi.org/10.1007/s10620-014-3368-z
Galtier, M., De Sordi, L., Sivignon, A., de ValleÌe, A., Maura, D., Neut, C., Rahmouni, O., Wannerberger, K., Darfeuille-Michaud, A., Desreumaux, P., Barnich, N. and Debarbieux, L., 2017. Bacteriophages targeting adherent invasive Escherichia coli strains as a promising new treatment for Crohnâs disease. Journal of Crohnâs and Colitis 11: 840-847. https://doi.org/10.1093/ecco-jcc/jjw224
Glasser, A.L., Boudeau, J., Barnich, N., Perruchot, M.H., Colombel, J.F. and Darfeuille-Michaud, A., 2001. Adherent invasive Escherichia coli strains from patients with Crohnâs disease survive and replicate within macrophages without inducing host cell death. Infection and Immunity 69: 5529-5537. https://doi.org/10.1128/IAI.69.9.5529-5537.2001
HrdyÌ, J., Alard, J., Couturier-Maillard, A., Boulard, O., Boutillier, D., Delacre, M., Lapadatescu, C., Cesaro, A., Blanc, P., Pot, B., Ryffel, B., Chamaillard, M. and Grangette, C., 2020. Lactobacillus reuteri 5454 and Bifidobacterium animalis ssp. lactis 5764 improve colitis while differentially impacting dendritic cells maturation and antimicrobial responses. Scientific Reports 10: 5345. https://doi.org/10.1038/s41598-020-62161-1
Ingrassia, I., Leplingard, A. and Darfeuille-Michaud, A., 2005. Lactobacillus casei DN-114 001 inhibits the ability of adherent-invasive Escherichia coli isolated from Crohnâs disease patients to adhere to and to invade intestinal epithelial cells. Applied and Environmental Microbiology 71: 2880-2887. https://doi.org/10.1128/AEM.71.6.2880-2887.2005
Irani, M., Sarmadi, M., Bernard, F., Ebrahimi Pour, G.H. and Shaker Bazarnov, H., 2010. Leaves antimicrobial activity of Glycyrrhiza glabra L. Iranian Journal of Pharmaceutical Research 9: 425-428.
Lagha, R., Ben Abdallah, F., Al-Sarhan, B.O. and Al-Sodany, Y., 2019. Antibacterial and biofilm inhibitory activity of medicinal plant essential oils against Escherichia coli isolated from UTI patients. Molecules 24: 1161. https://doi.org/10.3390/molecules24061161
Leccese, G., Bibi, A., Mazza, S., Facciotti, F., Caprioli, F., Landini, P. and Paroni, M., 2020. Probiotic Lactobacillus and Bifidobacterium strains counteract adherent-invasive Escherichia coli (AIEC) virulence and Hamper IL-23/Th17 axis in ulcerative colitis, but not in Crohnâs disease. Cells 9: 1824. https://doi.org/10.3390/cells9081824
Limketkai, B.N., Akobeng, A.K., Gordon, M. and Adepoju, A.A., 2020. Probiotics for induction of remission in Crohnâs disease. Cochrane Database of Systematic Reviews 7: CD006634. https://doi.org/10.1002/14651858.CD006634.pub3
Lu, X., Zhao, S., Ye, X. and Wang, P., 2023. Correlation between IL-8 gene polymorphisms and pathogenesis of Crohnâs disease. Alternative Therapies in Health and Medicine 29: 112-120.
Martinez-Medina, M., Aldeguer, X., Lopez-Siles, M., GonzaÌlez-Huix, F., LoÌpez-Oliu, C., Dahbi, G., Blanco, J.E., Blanco, J., Garcia-Gil, L.J. and Darfeuille-Michaud, A., 2009a. Molecular diversity of Escherichia coli in the human gut: new ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohnâs disease. Inflammatory Bowel Diseases 15: 872-882. https://doi.org/10.1002/ibd.20860
Martinez-Medina, M., Naves, P., Blanco, J., Aldeguer, X., Blanco, J.E., Blanco, M., Ponte, C., Soriano, F., Darfeuille-Michaud, A. and Garcia-Gil, L.J., 2009b. Biofilm formation as a novel phenotypic feature of adherent-invasive Escherichia coli (AIEC). BMC Microbiology 9: 202. https://doi.org/10.1186/1471-2180-9-202
Ni, Y., Zhang, Y., Zheng, L., Rong, N., Yang, Y., Gong, P., Yang, Y., Siwu, X., Zhang, C., Zhu, L. and Fu, Z., 2023. Bifidobacterium and Lactobacillus improve inflammatory bowel disease in zebrafish of different ages by regulating the intestinal mucosal barrier and microbiota. Life Sciences 324: 121699. https://doi.org/10.1016/j.lfs.2023.121699
Olivas-Quintero, S., Bernal-Reynaga, R., Lopez-Saucedo, C., Maldonado-Puga, S., DıÌaz-Camacho, S.P., Uribe-Carvajal, S., Delgado-Vargas, F. and Estrada-Garcia, T., 2022. Bacteriostatic effect of Echeveria extracts on diarrheagenic E. coli pathotypes and non-cytotoxicity on human Caco-2 cells. The Journal of Infection in Developing Countries 16: 147-156. https://doi.org/10.3855/jidc.15125
Plaza-Diaz, J., Ruiz-Ojeda, F.J., Gil-Campos, M. and Gil, A., 2019. Mechanisms of action of probiotics. Advances in Nutrition 10: S49-S66. https://doi.org/10.1093/advances/nmy063
Qiao, J., Sun, Z., Liang, D. and Li, H., 2020. Lactobacillus salivarius alleviates inflammation via NF-κB signaling in ETEC K88-induced IPEC-J2 cells. Journal of Animal Science and Biotechnology 11: 76. https://doi.org/10.1186/s40104-020-00488-5
Singh, U.P., Singh, N.P., Murphy, E.A., Price, R.L., Fayad, R., Nagarkatti, M. and Nagarkatti, P.S., 2016. Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine 77: 44-49. https://doi.org/10.1016/j.cyto.2015.10.008
Sivignon, A., de ValleÌe, A., Barnich, N., Denizot, J., Darcha, C., PigneÌde, G., Vandekerckove, P. and Darfeuille-Michaud, A., 2015a. Saccharomyces cerevisiae CNCM I-3856 prevents colitis induced by AIEC bacteria in the transgenic mouse model mimicking Crohnâs disease. Inflammatory Bowel Diseases 21: 276-286. https://doi.org/10.1097/MIB.0000000000000280
Sivignon, A., Yan, X., Alvarez Dorta, D., Bonnet, R., Bouckaert, J., Fleury, E., Bernard, J., Gouin, S.G., Darfeuille-Michaud, A. and Barnich, N., 2015b. Development of heptylmannoside-based glycoconjugate antiadhesive compounds against adherent-invasive Escherichia coli bacteria associated with Crohnâs disease. mBio 6: e01298-15. https://doi.org/10.1128/mBio.01298-15
Sivignon, A., Yu, S.-Y., Ballet, N., Vandekerckove, P., Barnich, N. and Guerardel, Y., 2021. Heteropolysaccharides from S. cerevisiae show anti-adhesive properties against E. coli associated with Crohnâs disease. Carbohydrate Polymers 271: 118415. https://doi.org/10.1016/j.carbpol.2021.118415
Szandruk-Bender, M., Rutkowska, M., Merwid-Ląd, A., Wiatrak, B., Szeląg, A., Dzimira, S., SobieszczanÌska, B., Krzystek-Korpacka, M., Kucharska, A.Z., Matuszewska, A., Nowak, B., PioÌrecki, N., Duda-Madej, A., Walczuk, U., Turniak, M., Bednarz-Misa, I. and SozanÌski, T., 2020. Cornelian cherry iridoid-polyphenolic extract improves mucosal epithelial barrier integrity in rat experimental colitis and exerts antimicrobial and antiadhesive activities in vitro. Oxidative Medicine and Cellular Longevity 2020: 7697851. https://doi.org/10.1155/2020/7697851
Tien, M.-T., Girardin, S.E., Regnault, B., Le Bourhis, L., Dillies, M.-A., CoppeÌe, J.-Y., Bourdet-Sicard, R., Sansonetti, P.J. and PeÌdron, T., 2006. Anti-inflammatory effect of Lactobacillus casei on Shigella-infected human intestinal epithelial cells. The Journal of Immunology 176: 1228-1237. https://doi.org/10.4049/jimmunol.176.2.1228
Torres-Maravilla, E., Carvalho, F.A., Holowacz, S., Delannoy, J., Lenoir, L., Jacouton, E., Barbut, F., Langella, P., BermuÌdez-HumaraÌn, L.G. and Waligora-Dupriet, A.-J., 2024. Screening of probiotic strains to improve visceral hypersensitivity in irritable bowel syndrome by using in vitro and in vivo approaches. Beneficial Microbes 15: 293-310. https://doi.org/10.1163/18762891-bja00006
Wang, L., Li, W., Li, X., Liu, J. and Chen, Y., 2023. Antimicrobial activity and mechanisms of walnut green husk extract. Molecules 28: 7981. https://doi.org/10.3390/molecules28247981
Wu, Z., Huang, S., Li, T., Li, N., Han, D., Zhang, B., Xu, Z.Z., Zhang, S., Pang, J., Wang, S., Zhang, G., Zhao, J. and Wang, J., 2021. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 9: 184. https://doi.org/10.1186/s40168-021-01115-9
Zhang, D., Verstrepen, L., De Medts, J., Duysburgh, C., Van den Abbeele, P., Marzorati, M. and Khoo, C., 2021. A cranberry concentrate decreases adhesion and invasion of Escherichia coli (AIEC) LF82 in vitro. Pathogens 10: 1217. https://doi.org/10.3390/pathogens10091217
Zhi, T., Hong, D., Zhang, Z., Li, S., Xia, J., Wang, C., Wu, Y., Jia, Y. and Ma, A., 2022. Anti-inflammatory and gut microbiota regulatory effects of walnut protein derived peptide LPF in vivo. Food Research International 152: 110875. https://doi.org/10.1016/j.foodres.2021.110875
ZÌurek, N., Pycia, K., PawÅowska, A., Potocki, L. and Kapusta, I.T., 2023. Chemical profiling, bioactive properties, and anticancer and antimicrobial potential of Juglans regia L. leaves. Molecules 28: 1989. https://doi.org/10.3390/molecules28041989







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}