Abstract
The effects of probiotics on the gut microbiota and microbial metabolites in healthy individuals are not well understood. Faecal and serum samples were collected at the start and end of a 3-month, double-blind, placebo-controlled, randomised study with three different probiotic formulations in free-living, healthy adults. The composition of the faecal microbiota and levels of faecal and/or serum short-chain fatty acids (SCFA) and bile acids (BA) were measured and the probiotic formulations were found to impart differing effects including shifts in the composition and structure of the faecal microbiota, enhanced levels of circulating short chain fatty acids such as butyrate and propionate, and elevated levels of sulphated bile acids in faeces. This was in contrast to the outcomes for the placebo population where very little change occurred over the study. These findings demonstrate that probiotic supplementation elicits formulation specific effects and that there are potential benefits for healthy individuals.
1 Introduction
A complex relationship exists between the trillions of bacteria residing in the gut (the gut microbiota) and the host (Fan and Pedersen, 2021) and the microbiota is involved in a multitude of functions within and beyond the gut that are fundamental to host health. Two key examples of these functions include the generation of short chain fatty acids (SCFAs) and the biotransformation of primary bile acids (BAs) into a wide variety of derivatives (Fan and Pedersen, 2021).
SCFAs such as acetate, butyrate and propionate are generated in the colon during the anaerobic fermentation of non-digestible dietary carbohydrates (and, to a lesser extent, amino acids) by Bacteroidota, Bacillota, Clostridiota and Actinomycetota (den Besten et al., 2013; Fusco et al., 2023). They are key regulators of intestinal immune homeostasis (Fusco et al., 2023) and their acidic properties help prevent the growth of pH sensitive pathogens whilst supporting the growth of commensal bacteria (den Besten et al., 2013). Butyrate is of particular importance, serving as the primary energy source of colonocytes and supporting the integrity of the intestinal epithelium (Recharla et al., 2023). Gut derived SCFAs can reach the peripheral tissues via the circulatory system where they impart beneficial adipogenic, inflammatory and neurogenic effects (van der Hee and Wells, 2021).
Bile acids (BAs) are released into the intestinal lumen during digestion to emulsify dietary fats and fat-soluble vitamins, facilitating their absorption. They are also involved in cellular signalling, immune modulation, and have antimicrobial activities (Larabi et al., 2023). The functional diversity of BAs, and the rate of their synthesis and excretion, is significantly influenced by the biotransformation processes mediated by the gut microbiota (Guzior and Quinn, 2021). Microbial bile salt hydrolases (BSHs) are gateway enzymes and catalyse the deconjugation of BAs to reduce their solubility and promote their excretion. BA deconjugation also enhances their antimicrobial properties, contributing to overall defence against pathogens (Bourgin et al., 2021; Larabi et al., 2023).
Probiotic bacteria are defined as âlive microorganisms that, when administered in adequate amounts, confer a health benefit on the hostâ (Hill et al., 2014) and there is evidence supporting their ability to alleviate gastrointestinal disorders such as irritable bowel syndrome (IBS) (Wu et al., 2024) and constipation (van der Schoot et al., 2022), but there are few studies of the impact of probiotic supplementation on healthy individuals. Little is known about their impact upon the gut microbiota and SCFA and BA metabolism in healthy subjects even though they represent the biggest consumers of probiotic supplements globally (Yilmaz-Ersan et al., 2020).
The tolerability of three distinct combinations of lactic acid bacteria and bifidobacteria was assessed in a double-blind, placebo-controlled, multi-arm, randomised study in healthy adults (Mullish et al., 2023). Probiotic-1 comprised two strains of Lactobacillus acidophilus, Bifidobacterium bifidum and Bifidobacterium animalis subsp. lactis, Probiotic-2 comprised the Probiotic-1 organisms with the addition of 13 others, and Probiotic-3 contained two strains of Lactobacillus rhamnosus and Bifidobacterium animalis subsp. lactis. The three formulations were found to exert markedly different effects on participantsâ gastrointestinal health (described in Mullish et al. (2023)) and here we provide details of their impact upon the faecal microbiota composition, the faecal and circulating levels of SCFA, and the faecal bile acid concentration.
2 Methods
Study design, recruitment and randomisation
This follow-up analysis was based on a single-centre, multi-arm, double-blind, randomised and placebo-controlled trial in healthy adults which assessed three different probiotic formulations. Eligible participants were assigned to one of four study arms, Placebo (PL), Probiotic-1 (P1), Probiotic-2 (P2) or Probiotic-3 (P3), at a ratio of 1:1:1:1 and they took the intervention daily for 84 days (3 months).
The intervention
The compositions and doses of the study interventions are shown in Table 1. Compliance to the intervention was monitored via the collection of unused capsules at the end of the study.

The outcomes
The outcomes presented here are changes in the faecal microbiota composition, the faecal and circulating levels of SCFA, and levels of faecal bile acids for those participants who provided faecal and/or serum samples at the start of the study and at the end of the study.
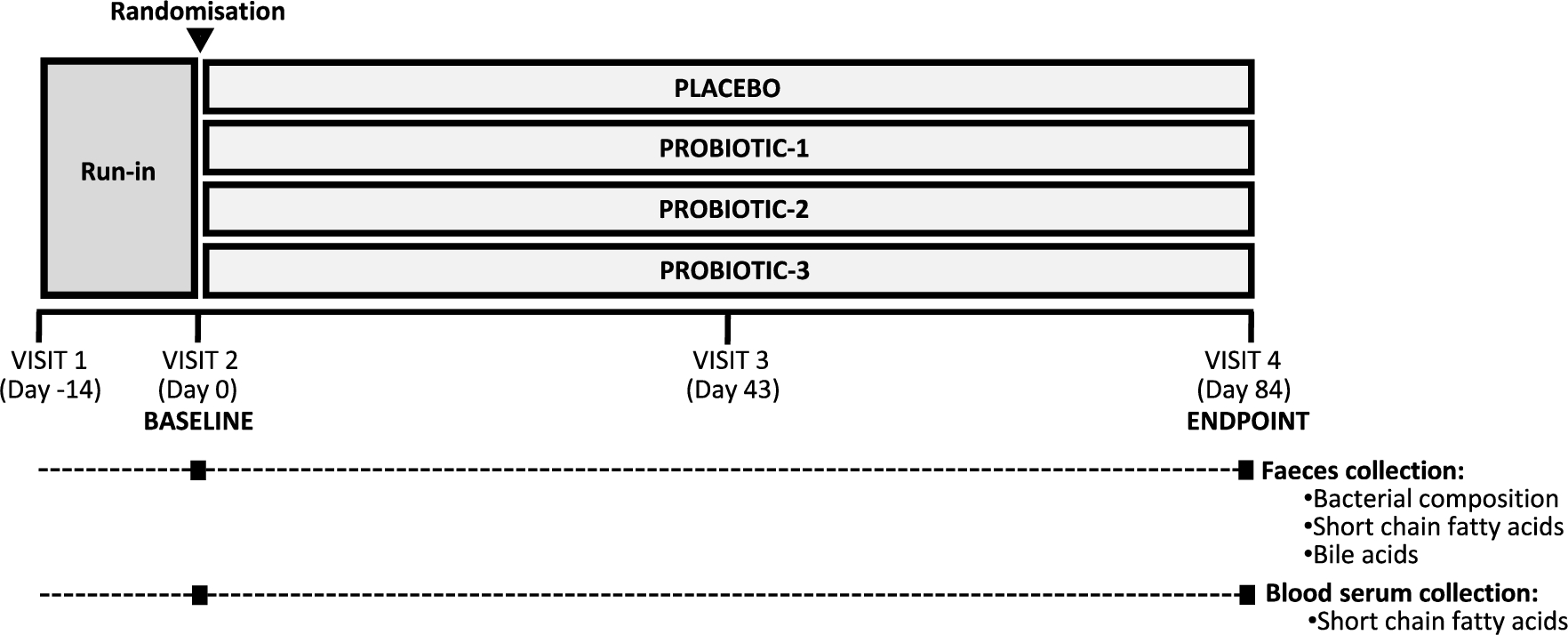
Schedule of sample collection.
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
Schedule of sample collection
The schedule of sample collection is shown in Figure 1. Participants visited the study centre on four occasions in total, and faecal and/or serum samples were collected on visit 2 (day 0, Baseline (BL)) and visit 4 (day 84, Endpoint (EP)).
Collection, storage and analysis of faecal samples
Collection and storage
Faecal samples were collected by the participants, frozen at â20 °C immediately after collection and transported frozen to the trial centre within two days, where they were stored at â80 °C pending analysis.
Genomic DNA extraction
Thawed faecal samples were mechanically lysed by bead beating for 3 à 30 s cycles (5 m/s) with 5 min intervals in Matrix Lysing D tubes and a FastPrep-24® bead beater (MPBIO, United States) and gDNA was extracted using the QIAamp® Fast DNA Stool Mini Kit (Qiagen, Germany) according to the manufacturerâs instructions.
Microbial profiling by next generation 16S rRNA gene sequencing
Sample libraries were prepared following Illuminaâs 16S Metagenomic Sequencing Library Preparation Protocol with the following alterations: amplification of the V1-V2 hypervariable region of the 16S rRNA gene was performed using custom-designed primers (Mullish et al., 2018) and the index PCR reactions were cleaned and normalised using the SequalPrep Normalisation Plate Kit (Life Technologies, UK). Sample libraries were quantified using Qubit fluorometry and NEBNext Library Quant Kit for Illumina (New England Biolabs, UK). Sequencing was performed on an Illumina MiSeq platform (Illumina Inc., UK) with the MiSeq Reagent Kit v3 (Illumina) using paired-end 2Â Ã 300Â bp chemistry. Raw sequencing data was processed following the DADA2 pipeline in R using the SILVA taxonomic database (version 138.1) to classify sequence variants, as described in Mullish et al. (2024b), and amplicon sequence variants (ASVs) were adjusted to the bacterial biomass within the sample, estimated using 16S rRNA gene qPCR as described previously (Mullish et al., 2019). To account for multiple sequencing batches, the ASV table was corrected through conditional quantile regression using the ConQuR package with default parameters (Ling et al., 2022).
SCFA analysis
Faecal SCFA were measured according to the method of Valdivia-Garcia et al. (2022). using 3-nitrophenylhydrazine derivatisation and liquid chromatography with tandem mass spectrometry (LC-MS/MS). Results are expressed as μmol per gram sample wet weight.
Bile acid profiling
Faecal samples were lyophilised and bile acids extracted according to the method of Mullish et al. (2018). Bile acid profiling was performed by the National Phenome Centre (Imperial College London, UK) using Ultra-Performance Liquid Chromatography Tandem Mass Spectrometry (UPLC-MS), as adapted from the method of Sarafian et al. (2015). Results are expressed as relative intensity (RI, from UPLC-MS) per gram sample dry weight.
Collection, storage and analysis of blood serum
Collection and storage
At BL and EP, fasted bloods were collected into serum separator tube vacutainers and the serum was separated by centrifugation (
SCFA analysis
SCFAs were measured using direct-injection gas chromatography with acidified water-extraction (Wang et al., 2020; Zhao et al., 2006). 20 μl of serum was acidified with 3.5 μl of concentrated orthophosphoric acid 2-ethylbutyric acid (Sigma-Aldrich) and was used as an internal standard. A Clarus 500 gas chromatograph with a flame ionisation detector (PerkinElmer 8500, Norwalk, CT, USA) and a TR-FFAP 30 m à 0.32 mm i.d. à 0.25 μm capillary column (Thermo Scientific) was used for separation of SCFAs. The temperature operation conditions for the GC separation/analysis were 90 °C for 1 min, 130 °C at 10 °C/min, hold for 3 min, then 200 °C at 10 °C/min, hold for 8 min. Nitrogen was used as a carrier; the injector temperature was 220 °C; the detector temperature was 240 °C with 5 μl injection volume. SCFAs were identified by comparing retention times of peaks with those of standards: acetic acid, propionic, valeric and butyric acid. PerkinElmer Total Chrom Navigator software was used for data acquisition. Results are expressed as ng/ml.
Data analysis & statistics
Next generation 16S rRNA gene sequencing
Data importation and diversity analyses were performed in R package Phyloseq (McMurdie and Holmes, 2013) with results plotted using ggplot2 (Wickham, 2016) and changes in spatial organisation within the study groups were observed with a non-metric multidimensional scaling (NMDS) plot based on the Bray-Curtis dissimilarity matrix. Pairwise PERMANOVA with 999 permutations were performed using the pairwise Adonis package (Martinez Arbizu, 2020). Homogeneity of dispersion was analysed using betadisper and permutest functions in the R vegan package (Oksanen et al., 2022). Amplicon sequence variants (ASVs) were centred log ratio (CLR) transformed before assessment of changes in relative abundances using the Wilcoxon rank sum test with Bonferroni correction for multiple comparisons. Values of
To identify keystone genera, centrality scores of (i) degree (number of direct connections to other genera), (ii) betweenness (frequency of appearing on the shortest path between genera), (iii) closeness (proximity to other genera) and (iv) Eigenvector (influence on highly connected genera) were calculated from networks of genera present in at least 10% of samples after a modified CLR transformation. Networks were generated using the NetCoMi R package (Peschel et al., 2020) and genus abundance correlations were calculated with the Semi-parametric Rank-Based Correlation and Partial Correlation Estimation for Quantitative Microbiome Data (SPRING) method (Yoon et al., 2019). A âhub scoreâ (representing overall influence of a genus on the microbiota) was determined by combining normalised scores for degree, betweenness, closeness and eigenvector and the 10 genera with highest scores were assigned as âkeystonesâ.
SCFA and bile acids
Multivariate analysis was performed using principal component analysis (PCA) on log transformed data with P values generated by PERMANOVA with Bonferroni correction. Univariate analysis was performed using the Wilcoxon signed rank test (GraphPad Prism, Version 10.20.2) with Bonferroni correction. Where appropriate, data was
3 Results
Recruitment, compliance and demographics
In total 96 participants were recruited to the study, with 23 allocated to the placebo group, 24 to P1, 24 to P2 and 25 to P3. The baseline characteristics were well matched between groups (Supplementary Table S1A). Participants were aged between 19 and 64 years, had a BMI between 18.99 to 29.90 kg/m2 and were in good general health. 69.9% of the study population were female. Compliance to the study intervention exceeded 90%. The number of participants providing samples of faeces and/or serum at both BL and EP are shown in Table 2. The baseline characteristics of participants included in each analysis are provided in Supplementary Table S1B and are representative of the complete study groups and each other.
Analysis subpopulations
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
The impact of probiotic supplementation on the faecal microbiota
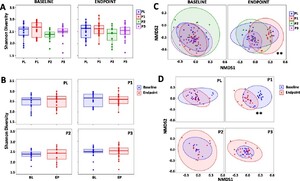
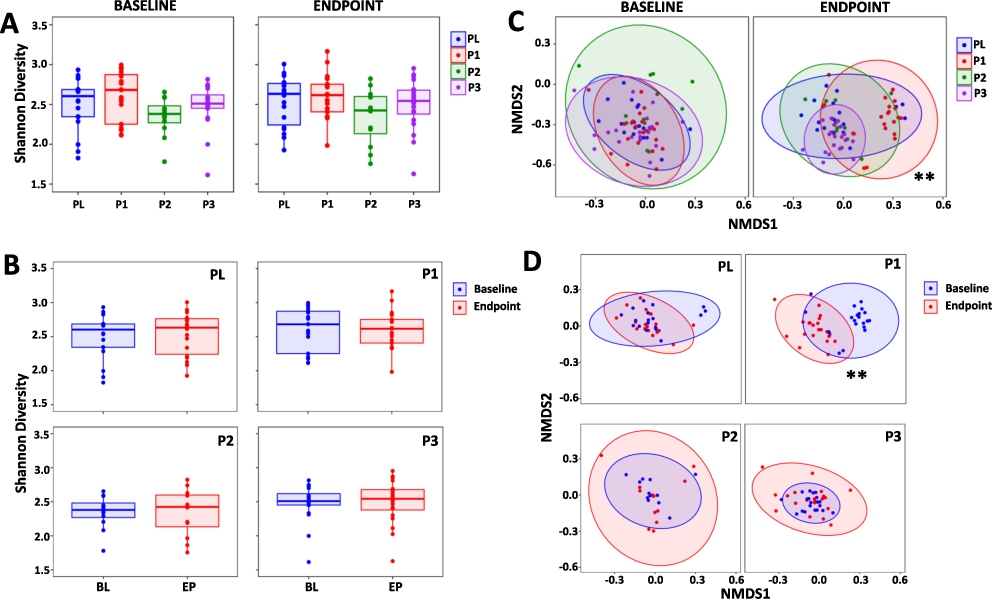
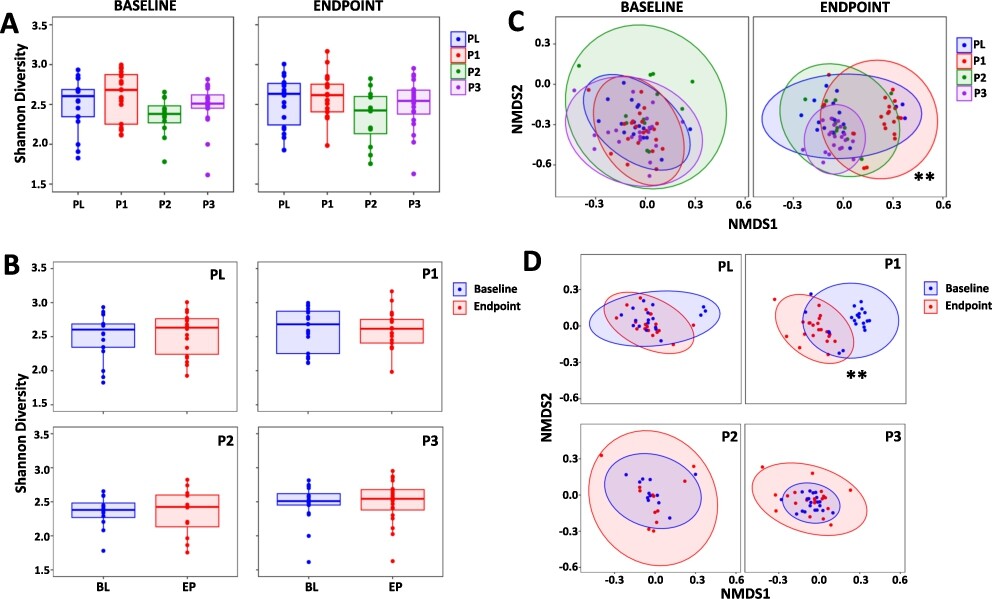
Shannonâs α-diversity (Figure 2A) and Simpson α-diversity index (Supplementary Figure S1) were assessed and there were no differences between the study groups at baseline or endpoint, nor did any changes occur within each study group over the duration of the study (Figure 2B). With regard to β-diversity, there were no differences between the study groups at the baseline, however, at endpoint there were significant differences for the P1 group compared to PL (
Changes in the composition of the gut microbiota. Shannonâs α-diversity measure comparing (A) the PL, P1, P2 and P3 groups at the baseline (BL) and at endpoint (EP) and (B) BL with EP within the PL, P1, P2 and P3 groups. (C) non-metric multidimensional scaling (NMDS) plots based on the Bray-Curtis dissimilarity matrix comparing the PL, P1, P2 and P3 groups at BL and at EP and (D) BL vs EP for the PL, P1, P2 and P3 groups. Values of P; **
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
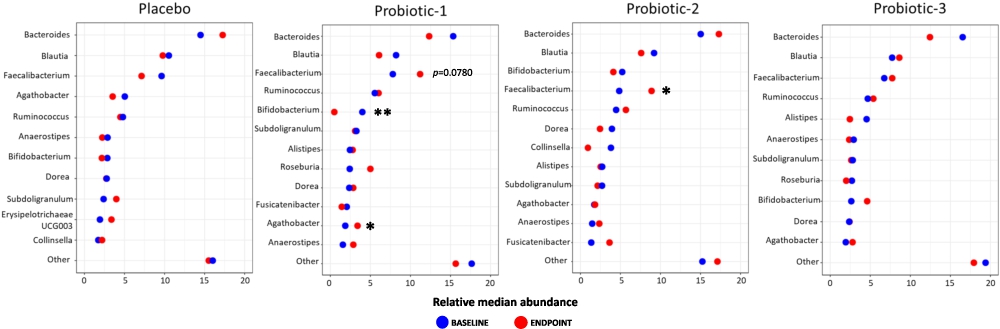
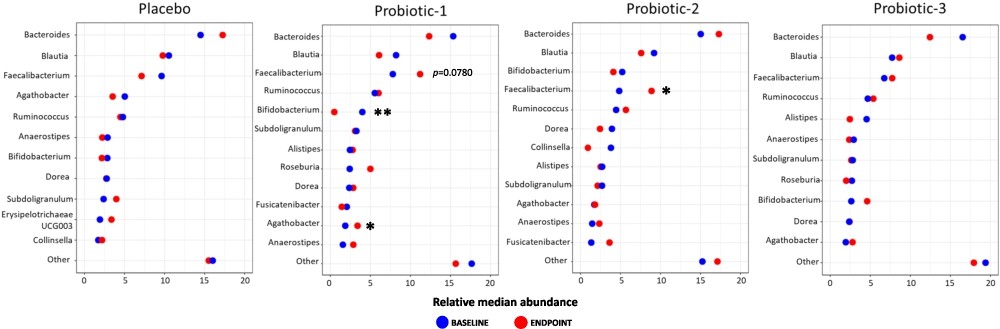
Changes in the relative abundance between BL and EP of the 10 most prevalent bacterial genera within each study group are shown in Figure 3 (top 30 shown in Supplementary Figure S2). In P1, there was a significant increase in the abundance of Agathobacter (
Microbiota abundance at genus level. Dumbbell plots showing changes in relative median abundance between baseline (BL) and endpoint (EP) of 10 most prevalent bacterial genera for PL, P1, P2 and P3 groups. Data is presented in descending order of abundance according to the group BL values. Values of P; *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
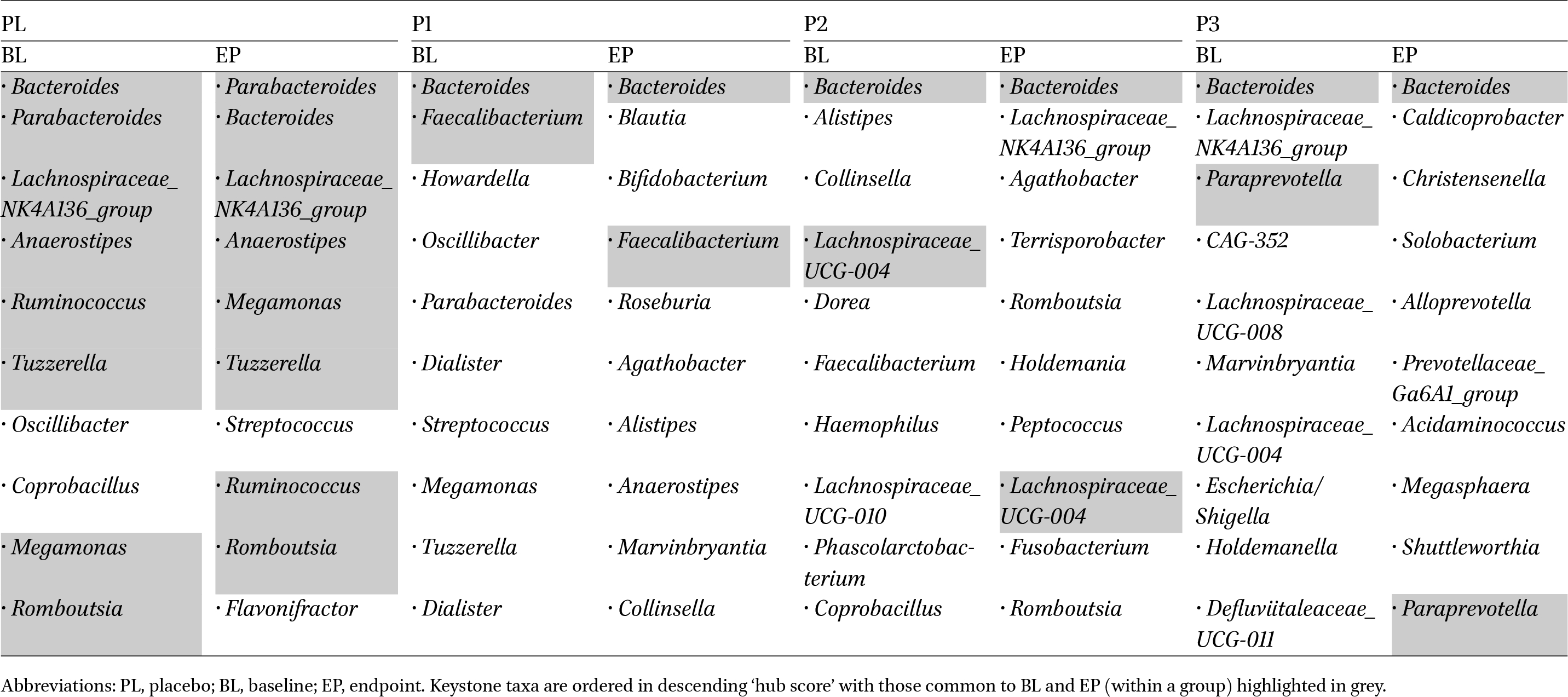
Keystone organisms have a disproportionate influence on the composition, function and stability of a microbiome, irrespective of their abundance (Banerjee et al., 2018; Mills et al., 1993) and Table 3 shows the top 10 keystone genera identified at baseline and endpoint for each study group (rankings of degree, centrality, closeness and eigenvector are provided in Supplementary Table S2). Within the placebo, there was little change in keystone genera between BL and EP with eight taxa detected at both time-points but within the probiotic groups there were considerable changes in keystone profiles. For P1, only two genera appeared at both BL and EP which were Bacteroides and Faecalibacterium; two groups for P2, Bacteroides and Lachnospiraceae_UCG-004 and two groups for P3, Bacteroides and Paraprevotella.
The impact of probiotic supplementation on SCFA levels
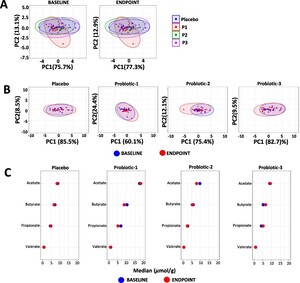
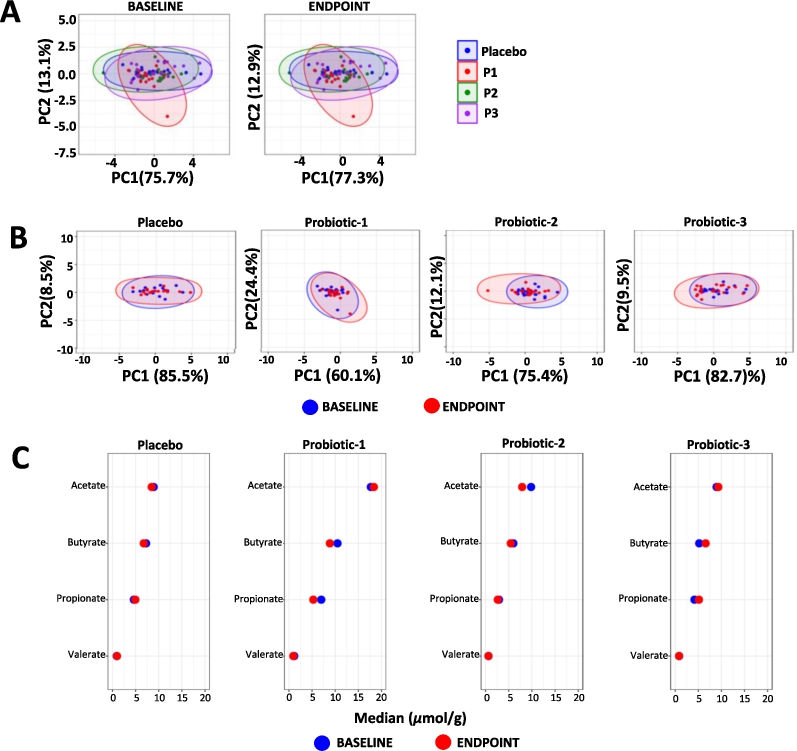
The overall profiles of faecal SCFA were unchanged both between groups at BL and at EP (Figure 4A) and within each group (Figure 4B). A univariate analysis of changes in faecal SCFA concentrations within each study group is shown in Figure 4C (detailed data in Supplementary Table S3) and no significant changes were observed.
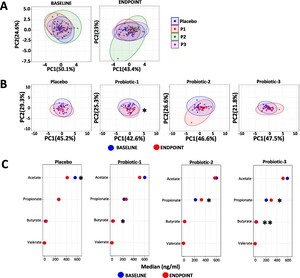
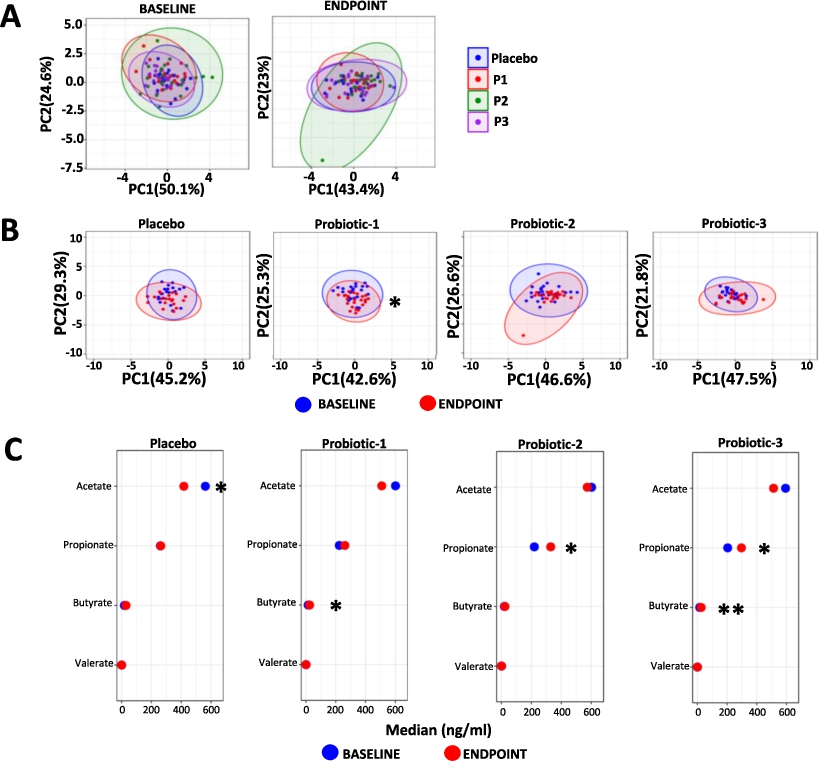
Changes in the overall serum SCFA profiles are shown in Figure 5 (detailed data in Supplementary Table S4) with no between group differences observed at the baseline or at the end point (Figure 5A). Within group serum SCFA profiles changed significantly only for P1 between BL and EP (
Top 10 ranked keystone taxa within the study groups
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
The impact of probiotic supplementation on faecal bile acids levels
There were no significant between (Figure 6A) or within (Figure 6B) group differences in overall faecal bile acid profiles at BL or at EP. Figure 6C summarises the results of a univariate analysis of the changes in bile acids within each study group (detailed data in Supplementary Table S5) and there was a significant increase in 7KLCA (
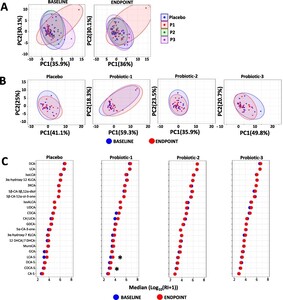
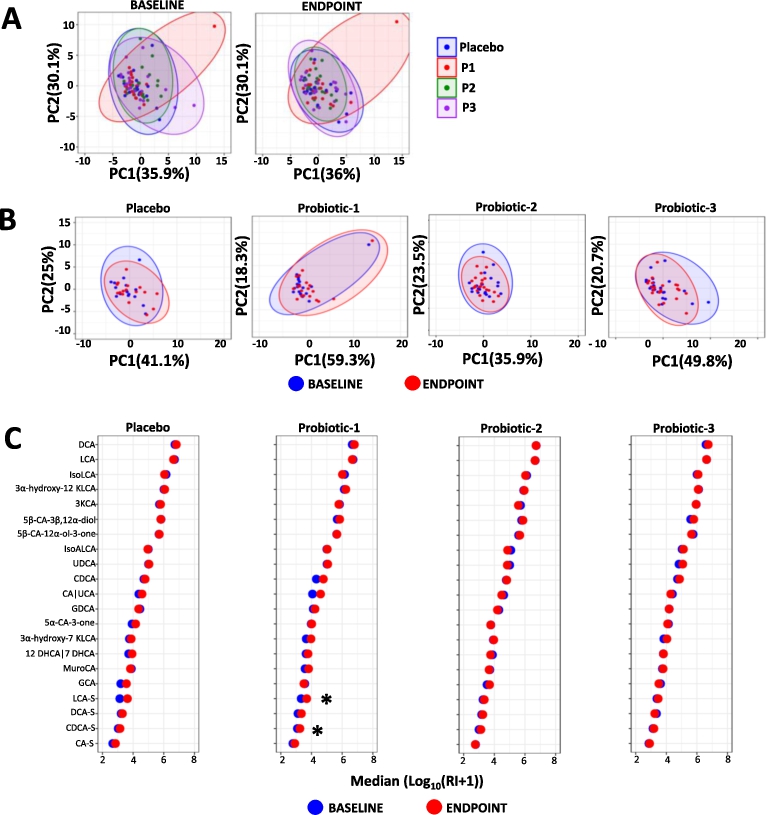
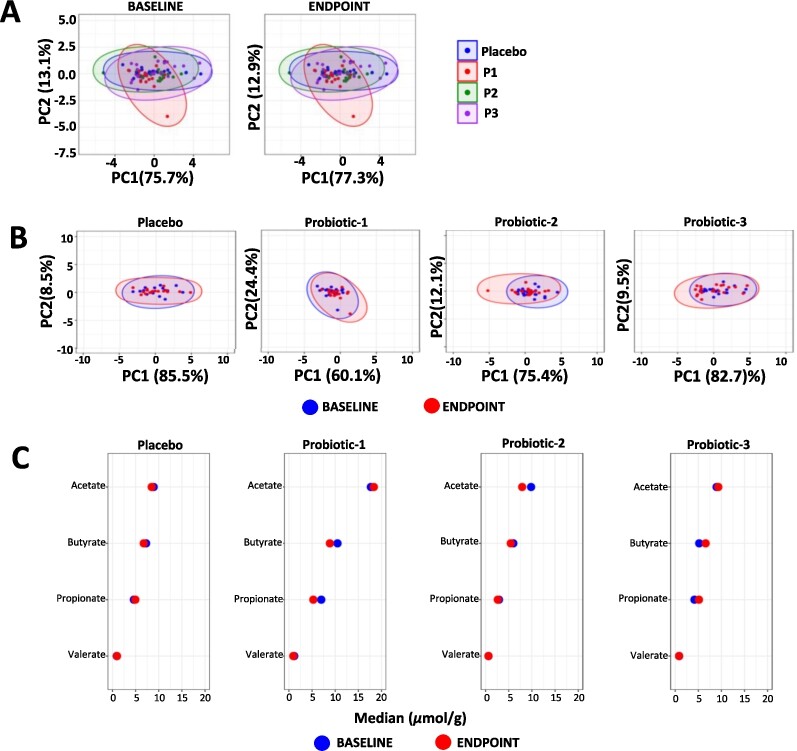
Changes in faecal SCFA. Principle component analysis (PCA) comparing faecal SCFA profiles between (A) the PL, P1, P2 and P3 groups at baseline (BL) and at endpoint (EP) and (B) within the PL, P1, P2 and P3 groups at BL and EP. (C) dumbbell plots showing changes in individual SCFA between BL and EP within the PL, P1, P2 and P3 groups. Data is presented as median μmol per g dry weight faeces and is shown in descending order of abundance according to the placebo at BL. Values of P; *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
Changes in serum SCFA. Principle component analysis (PCA) comparing serum SCFA profiles (A) between the PL, P1, P2 and P3 groups at baseline (BL) and endpoint (EP) and (B) within the PL, P1, P2 and P3 groups between BL and EP. (C) dumbbell plots showing changes in individual SCFA between BL and EP for all groups. Data is presented as median ng/ml and is shown in descending order of abundance according to the placebo at BL. Values of P: *
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
Changes in faecal bile acids. Principle component analysis (PCA) comparing faecal bile acid profiles (A) between the PL, P1, P2 and P3 groups at the baseline (BL) and at endpoint (EP) and (B) within PL, P1, P2 and P3 groups. (C) dumbbell plots comparing individual bile acids between BL and EP for the placebo, P1, P2 and P3 groups. Data is presented as the median of
Citation: Beneficial Microbes 17, 3 (2026) ; 10.1163/18762891-bja00096
4 Discussion
Faecal and blood serum samples were collected from four groups of healthy adults receiving one of three distinct probiotic supplements or a matching placebo daily for three months. The placebo group was subject to very few changes over the course of the study whereas each of the probiotics exerted varying effects on the composition of the faecal microbiota and on the levels of SCFAs and bile acids.
Probiotic-1 (P1), previously shown to improve gastrointestinal health in both healthy subjects (Mullish et al., 2023) and those with Irritable Bowel Syndrome (Mullish et al., 2024b; Williams et al., 2009), altered the composition and keystone organisms in the faecal microbiota, serum SCFA profile and faecal bile acid composition. Probiotic-2 (P2), comprising the component organisms of Probiotic-1 with 13 others, and Probiotic-3 (P3), comprising organisms with recognised gastrointestinal benefits (Bonfrate et al., 2020; Cheng et al., 2021; Pedersen et al., 2014), altered keystone organisms and serum SCFA levels.
The data presented previously (Mullish et al., 2023) using traditional microbial culture techniques to determine differences in faecal viable numbers compared to the placebo indicated decreases for Bacteroides with P1, increased lactobacilli in P2 and for P3, increases in lactobacilli and decreases in Bacteroides. Next generation sequencing identified a significant shift in β-diversity within the P1 group (the P1 formulation comprises two strains of bifidobacteria and two strains of lactobacilli). Interestingly, no changes in diversity were observed in the P2 probiotic which consists of the organisms present in the P1 formulation together with 13 more organisms. The differing response may be due to âdilutionâ by the additional bacteria or, potentially, antagonistic effects among the organisms included in this multistrain population (McFarland et al., 2018; Ouwehand et al., 2018).
At the genus level, within the P1 group there was an increase in the relative abundance of Agathobacter and Faecalibacterium and a decrease in Bifidobacterium and for the P2 group, there was an increase in abundance of Faecalibacterium; there were no changes in the PL or P3 groups. The genus Agathobacter comprises SCFA producers (Van-Wehle and Vital, 2024) and previously the P1 formulation supported an increased abundance of Agathobacter in IBS sufferers alongside a reduced symptom severity (Mullish et al., 2024b). Some members of Faecalibacterium, such a F. prausnitzii, produce butyrate, and the prevalence of these organisms suggests a âhealthy gut microbiotaâ (MartıÌn et al., 2023; Van-Wehle and Vital, 2024). The reduced levels of Bifidobacterium in the P1 group does not correspond with the viable enumeration results (Mullish et al., 2023)
Keystone organisms are those which have a disproportionately large influence on the composition, stability and/or function of a microbial community, irrespective of their abundance (Banerjee et al., 2018; Mills et al., 1993). The keystone organisms of the PL group changed very little between the start to the end of the study, but for all of the probiotic groups there was near complete re-structuring of keystone organisms, leading us to hypothesise that the probiotics may impact upon the structure/organisation and functionality of the microbiota without imparting gross changes to composition. In each of the probiotic groups, the endpoint profiles of keystones organisms were rich in carbohydrate metabolizing bacteria (Roseburia, Agathabacter and Megasphaera) and BSH positive bacteria (Bifidobacterium, Romboutsia and Christensella) which could be linked to the changes in SCFAs and bile acids in these healthy subjects. For the P1 group, Bifidobacterium was identified as one of the endpoint keystone organisms despite the decreased abundance of this organism. The composition of the post supplementation keystone profile for P1 compares closely with that seen in other studies with the same formulation (Mullish et al., 2024a).
There were no changes in faecal SCFA levels within the placebo nor any of the probiotic groups but faecal SCFA concentrations do not necessarily reflect circulating levels (Nogal et al., 2023). A significant shift in the serum SCFA profile was observed in the P1 group alone. Univariate analyses of P1 revealed a significant increase in circulating levels of butyrate and the benefits linked with butyrate include anti-constipatory, anti-inflammatory, and neuroprotective effects (Xiong et al., 2022). The P2 group was associated with elevated serum propionate levels, while P3 showed increased levels of both butyrate and propionate. Like butyrate, propionate is associated with a range of health-promoting effects (reviewed in Xiong et al. (2022)).
Another vital function of the gut microbiota is the biotransformation of bile acids, a process that plays a critical role in maintaining host health (Fogelson et al., 2023). Although there were no changes in overall faecal bile acid profiles the P1 group had increases in the levels of two sulphated bile acids â lithocholic acid sulfate (LCA-S) and chenodeoxycholic acid sulfate (CDCA-S). Increases in sulphated secondary bile acids (glycolithocholic acid-3-sulfate) have been seen in germ-free mice supplemented with Lactobacillus gasseri LA39 (Hu et al., 2023).
Bile acid sulphation is a detoxification process, mitigating the cytotoxic effects of bile acids and contributing to gut homeostasis and overall health (Alnouti, 2009; Camilleri, 2022). Sulphation of bile acids is mediated by sulfotransferase enzymes, which are found in gut microbes (Langford and Shah, 2024), but are primarily expressed in the host liver and intestinal epithelium (Chen et al., 2003; Dew et al., 1980; Teubner et al., 2007). In rats, the expression of hepatic and intestinal sulfotransferases is strongly influenced by the gut microbiota (Meinl et al., 2009) and in mice, supplementation with the VSL#3 probiotic has been shown to upregulate hepatic expression of sulfotransferase genes (Jena et al., 2019). BSHs are involved in the deconjugation of bile acids and we have shown that the P1 formulation has BSH activity in vitro and in vivo (Michael et al., 2017), and may promote deconjugation of bile acids in the intestine, thus enlarging the pool of free bile acids which are more readily available for subsequent host-mediated sulphation. Lactobacillus rhamnosus GG, present in P3, has some BSH activity in vitro (HernaÌndez-GoÌmez et al., 2021) and has been shown to promote bile acid deconjugation in vivo (Hu et al., 2024).
Future research should include powered studies to confirm the observed microbial and metabolic changes and include faecal shotgun metagenomics and serum bile acid analysis to better understand the probiotic impact upon microbiota functionality. Laboratory-based studies could help to provide a more in-depth evaluation of the ability of the probiotics to influence SCFA absorption and metabolism, and to explore the potential role of probiotics in the process of bile acid sulphation by the intestinal epithelium.
In summary, this study demonstrates that different probiotic formulations impact the gut microbiota composition, SCFA absorption, and bile acid metabolism in healthy adults in a formulation specific manner. It also suggests that there are potential benefits for healthy individuals who choose to take probiotics regularly.
Corresponding author; e-mail:Â darynm@cultech.co.uk
Acknowledgements
We appreciate the technical assistance of Josh Kerry-Smith, Victoria Clark and Tamsin Joyce.
Authorsâ contribution
DRM: Conceptualisation, Visualisation, Writing (original draft), Supervision
DAJ: Data curation, Formal analysis, Validation, Visualisation
NC: Data curation, Formal analysis, Validation, Visualisation
IG: Methodology, Investigation
JAKM: Methodology, Investigation
NPD: Methodology, Data curation, Formal analysis, Investigation
MAVG: Methodology, Investigation
GR: Writing (review & editing)
SFP: Conceptualisation, Funding acquisition, Resource, Supervision, Writing (review & editing)
DW: Formal analysis, Writing (review & editing)
JRM: Conceptualisation, Writing (review & editing)
BHM: Conceptualisation, Formal analysis, Methodology, Writing (original draft)
Conflict of interest
BHM was the recipient of an NIHR Academic Clinical Lectureship (CL-2019-21-002) and now is the recipient of a Medical Research Council (MRC) Clinician Scientist award (MR/Z504002/1). The Division of Digestive Diseases at Imperial College London receives financial support from the National Institute of Health Research (NIHR) Imperial Biomedical Research Centre (BRC) based at Imperial College London and Imperial College Healthcare NHS Trust. Metabolomic assays were performed at the National Phenome Centre at Imperial College London, which is supported by the MRC and NIHR [grant number MC_PC_12025], and the MRC UK Consortium for MetAbolic Phenotyping (MAP UK) [grant number MR/S010483/1]. DW has previously received consultancy fees from Cultech Ltd. Cultech provided the intervention for the study. DRM, DJ, NC and SFP are/were employees of Cultech Ltd and had no role in the study design, recruitment or data collection. The remaining authors declare that they have no competing interests.
Data availability
Data is available from the corresponding author upon reasonable request. Sequence data generated during the current study has been deposited in the European Molecular Biology Laboratory (EMBL) nucleotide sequence database (https://www.ebi.ac.uk/ena) under accession number PRJEB83449.
References
Alnouti, Y., 2009. Bile acid sulfation: a pathway of bile acid elimination and detoxification. Toxicological Sciences 108: 225-246. https://doi.org/10.1093/toxsci/kfn268
Banerjee, S., Schlaeppi, K. and van der Heijden, M.G.A., 2018. Keystone taxa as drivers of microbiome structure and functioning. Nature Reviews Microbiology 16: 567-576. https://doi.org/10.1038/s41579-018-0024-1
Bonfrate, L., Di Palo, D.M., Celano, G., Albert, A., Vitellio, P., De Angelis, M., Gobbetti, M. and Portincasa, P., 2020. Effects of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 in IBS patients. European Journal of Clinical Nutrition 50: e13201. https://doi.org/10.1111/eci.13201
Bourgin, M., Kriaa, A., Mkaouar, H., Mariaule, V., Jablaoui, A., Maguin, E. and Rhimi, M., 2021. Bile salt hydrolases: at the crossroads of microbiota and human health. Microorganisms 9. https://doi.org/10.3390/microorganisms9061122
Camilleri, M., 2022. Bile acid detergency: permeability, inflammation, and effects of sulfation. American Journal of Physiology: Gastrointestinal and Liver Physiology 322: G480-G488. https://doi.org/10.1152/ajpgi.00011.2022
Chen, G., Zhang, D., Jing, N., Yin, S., Falany, C.N. and Radominska-Pandya, A., 2003. Human gastrointestinal sulfotransferases: identification and distribution. Toxicology and Applied Pharmacology 187: 186-197. https://doi.org/10.1016/s0041-008x(02)00073-x
Cheng, J., Laitila, A. and Ouwehand, A.C., 2021. Bifidobacterium animalis subsp. lactis HN019 effects on gut health: a review. Frontiers in Nutrition 8: 790561. https://doi.org/10.3389/fnut.2021.790561
den Besten, G., van Eunen, K., Groen, A.K., Venema, K., Reijngoud, D.J. and Bakker, B.M., 2013. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. Journal of Lipid Research 54: 2325-2340. https://doi.org/10.1194/jlr.R036012
Dew, M.J., Hawker, P.C., Nutter, S. and Allan, R.N., 1980. Human intestinal sulphation of lithocholate: a new site for bile acid metabolism. Life Sciences 27: 317-323. https://doi.org/10.1016/0024-3205(80)90199-X
Fan, Y. and Pedersen, O., 2021. Gut microbiota in human metabolic health and disease. Nature Reviews Microbiology 19: 55-71. https://doi.org/10.1038/s41579-020-0433-9
Fogelson, K.A., Dorrestein, P.C., Zarrinpar, A. and Knight, R., 2023. The gut microbial bile acid modulation and its relevance to digestive health and diseases. Gastroenterology 164: 1069-1085. https://doi.org/10.1053/j.gastro.2023.02.022
Fusco, W., Lorenzo, M.B., Cintoni, M., Porcari, S., Rinninella, E., Kaitsas, F., Lener, E., Mele, M.C., Gasbarrini, A., Collado, M.C., Cammarota, G. and Ianiro, G., 2023. Short-chain fatty-acid-producing bacteria: key components of the human gut microbiota. Nutrients 15. https://doi.org/10.3390/nu15092211
Guzior, D.V. and Quinn, R.A., 2021. Review: microbial transformations of human bile acids. Microbiome 9: 140. https://doi.org/10.1186/s40168-021-01101-1
HernaÌndez-GoÌmez, J.G., LoÌpez-Bonilla, A., Trejo-Tapia, G., AÌvila-Reyes, S.V., JimeÌnez-Aparicio, A.R. and HernaÌndez-SaÌnchez, H., 2021. In vitro bile salt hydrolase (BSH) activity screening of different probiotic microorganisms. Foods 10. https://doi.org/10.3390/foods10030674
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology & Hepatology 11: 506-514. https://doi.org/10.1038/nrgastro.2014.66
Hu, J., Hou, Q., Zheng, W., Yang, T. and Yan, X., 2023. Lactobacillus gasseri LA39 promotes hepatic primary bile acid biosynthesis and intestinal secondary bile acid biotransformation. Journal of Zhejiang University Science B 24: 734-748. https://doi.org/10.1631/jzus.B2200439
Hu, S., Tang, B., Lu, C., Wang, S., Wu, L., Lei, Y., Tang, L., Zhu, H., Wang, D. and Yang, S., 2024. Lactobacillus rhamnosus GG ameliorates triptolide-induced liver injury through modulation of the bile acid-FXR axis. Pharmacological Research 206: 107275. https://doi.org/10.1016/j.phrs.2024.107275
Jena, P.K., Sheng, L., Li, Y. and Wan, Y.-J.Y., 2019. Probiotics VSL#3 are effective in reversing non-alcoholic steatohepatitis in a mouse model. Hepatobiliary Surgery and Nutrition 9: 170-182.
Langford, L. and Shah, D.D., 2024. Bioinformatic analysis of sulfotransferases from an unexplored gut microbe, sutterella wadsworthensis 3_1_45B: possible roles towards detoxification via sulfonation by members of the human gut microbiome. International Journal of Molecular Sciences 25. https://doi.org/10.3390/ijms25052983
Larabi, A.B., Masson, H.L.P. and Baumler, A.J., 2023. Bile acids as modulators of gut microbiota composition and function. Gut Microbes 15: 2172671. https://doi.org/10.1080/19490976.2023.2172671
Ling, W., Lu, J., Zhao, N., Lulla, A., Plantinga, A.M., Fu, W., Zhang, A., Liu, H., Song, H., Li, Z., Chen, J., Randolph, T.W., Koay, W.L.A., White, J.R., Launer, L.J., Fodor, A.A., Meyer, K.A. and Wu, M.C., 2022. Batch effects removal for microbiome data via conditional quantile regression. Nature Communications 13: 5418. https://doi.org/10.1038/s41467-022-33071-9
MartıÌn, R., Rios-Covian, D., Huillet, E., Auger, S., Khazaal, S., BermuÌdez-HumaraÌn, L.G., Sokol, H., Chatel, J.-M. and Langella, P., 2023. Faecalibacterium: a bacterial genus with promising human health applications. FEMS Microbiology Reviews 47. https://doi.org/10.1093/femsre/fuad039
Martinez Arbizu, P., 2020. pairwiseAdonis: pairwise multilevel comparison using adonis. R package version 0.4.
McFarland, L.V., Evans, C.T. and Goldstein, E.J.C., 2018. Strain-specificity and disease-specificity of probiotic efficacy: a systematic review and meta-analysis. Frontiers in Medicine 5: 124. https://doi.org/10.3389/fmed.2018.00124
McMurdie, P.J. and Holmes, S., 2013. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 8: e61217. https://doi.org/10.1371/journal.pone.0061217
Meinl, W., Sczesny, S., Brigelius-FloheÌ, R., Blaut, M. and Glatt, H., 2009. Impact of gut microbiota on intestinal and hepatic levels of phase 2 xenobiotic-metabolizing enzymes in the rat. Drug Metabolism and Disposition 37: 1179-1186. https://doi.org/10.1124/dmd.108.025916
Michael, D.R., Davies, T.S., Moss, J.W.E., Calvente, D.L., Ramji, D.P., Marchesi, J.R., Pechlivanis, A., Plummer, S.F. and Hughes, T.R., 2017. The anti-cholesterolaemic effect of a consortium of probiotics: an acute study in C57BL/6J mice. Scientific Reports 7: 2883. https://doi.org/10.1038/s41598-017-02889-5
Mills, L., SouleÌ, M. and Doak, D., 1993. The keystone-species concept in ecology and conservation. Bioscience 43: 219-224. https://doi.org/10.2307/1312122
Mullish, B.H., McDonald, J.A.K., Pechlivanis, A., Allegretti, J.R., Kao, D., Barker, G.F., Kapila, D., Petrof, E.O., Joyce, S.A., Gahan, C.G.M., Glegola-Madejska, I., Williams, H.R.T., Holmes, E., Clarke, T.B., Thursz, M.R. and Marchesi, J.R., 2019. Microbial bile salt hydrolases mediate the efficacy of faecal microbiota transplant in the treatment of recurrent Clostridioides difficile infection. Gut 68: 1791-1800. https://doi.org/10.1136/gutjnl-2018-317842
Mullish, B.H., Michael, D.R., Dabcheva, M., Webberley, T.S., Coates, N., John, D.A., Wang, D., Luo, Y., Plummer, S.F. and Marchesi, J.R., 2024a. Authorsâ reply to letter: he who controls Clostridia and Bacteroidia controls the gut microbiome: the concept of targeted probiotics to restore the balance of keystone taxa in irritable bowel syndrome. Neurogastroenterology and Motility 36: e14820. https://doi.org/10.1111/nmo.14820
Mullish, B.H., Michael, D.R., Dabcheva, M., Webberley, T.S., Coates, N., John, D.A., Wang, D., Luo, Y., Plummer, S.F. and Marchesi, J.R., 2024b. A double-blind, randomized, placebo-controlled study assessing the impact of probiotic supplementation on the symptoms of irritable bowel syndrome in females. Neurogastroenterology and Motility: e14751. https://doi.org/10.1111/nmo.14751
Mullish, B.H., Michael, D.R., Webberley, T.S., John, D., Ramanathan, G., Plummer, S.F., Wang, D. and Marchesi, J.R., 2023. The gastrointestinal status of healthy adults: a post hoc assessment of the impact of three distinct probiotics. Beneficial Microbes: 1-14. https://doi.org/10.3920/BM2022.0092
Mullish, B.H., Pechlivanis, A., Barker, G.F., Thursz, M.R., Marchesi, J.R. and McDonald, J.A.K., 2018. Functional microbiomics: evaluation of gut microbiota-bile acid metabolism interactions in health and disease. Methods 149: 49-58. https://doi.org/10.1016/j.ymeth.2018.04.028
Nogal, A., Asnicar, F., Vijay, A., Kouraki, A., Visconti, A., Louca, P., Wong, K., Baleanu, A.F., Giordano, F., Wolf, J., Hadjigeorgiou, G., Davies, R., Michelotti, G.A., Franks, P.W., Berry, S.E., Falchi, M., Ikram, A., Ollivere, B.J., Zheng, A., Nightingale, J., Mangino, M., Segata, N., Bulsiewicz, W.J., Spector, T.D., Valdes, A.M. and Menni, C., 2023. Genetic and gut microbiome determinants of SCFA circulating and fecal levels, postprandial responses and links to chronic and acute inflammation. Gut Microbes 15: 2240050. https://doi.org/10.1080/19490976.2023.2240050
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., OâHara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H. and Wagner, H., 2022. Vegan: community Ecology Package. Available at: https://cran.r-project.org/web/packages/vegan/index.html
Ouwehand, A.C., Invernici, M.M., Furlaneto, F.A.C. and Messora, M.R., 2018. Effectiveness of multistrain versus single-strain probiotics: current status and recommendations for the future. Journal of Clinical Gastroenterology 52(Suppl 1): S35-S40. https://doi.org/10.1097/MCG.0000000000001052. Proceedings from the 9th Probiotics, Prebiotics and New Foods, Nutraceuticals and Botanicals for Nutrition & Human and Microbiota Health Meeting, held in Rome, Italy from September 10 to 12, 2017.
Pedersen, N., Andersen, N.N., VeÌgh, Z., Jensen, L., Ankersen, D.V., Felding, M., Simonsen, M.H., Burisch, J. and Munkholm, P., 2014. Ehealth: low FODMAP diet vs Lactobacillus rhamnosus GG in irritable bowel syndrome. World Journal of Gastroenterology 20: 16215-16226. https://doi.org/10.3748/wjg.v20.i43.16215
Peschel, S., Müller, C.L., von Mutius, E., Boulesteix, A.-L. and Depner, M., 2020. NetCoMi: network construction and comparison for microbiome data in R. Briefings in Bioinformatics 22. https://doi.org/10.1093/bib/bbaa290
Recharla, N., Geesala, R. and Shi, X.Z., 2023. Gut microbial metabolite butyrate and its therapeutic role in inflammatory bowel disease: a literature review. Nutrients 15. https://doi.org/10.3390/nu15102275
Sarafian, M.H., Lewis, M.R., Pechlivanis, A., Ralphs, S., McPhail, M.J., Patel, V.C., Dumas, M.E., Holmes, E. and Nicholson, J.K., 2015. Bile acid profiling and quantification in biofluids using ultra-performance liquid chromatography tandem mass spectrometry. Analytical Chemistry 87: 9662-9670. https://doi.org/10.1021/acs.analchem.5b01556
Teubner, W., Meinl, W., Florian, S., Kretzschmar, M. and Glatt, H., 2007. Identification and localization of soluble sulfotransferases in the human gastrointestinal tract. Biochemical Journal 404: 207-215. https://doi.org/10.1042/bj20061431
Valdivia-Garcia, M.A., Chappell, K.E., Camuzeaux, S., Olmo-GarcıÌa, L., van der Sluis, V.H., Radhakrishnan, S.T., Stephens, H., Bouri, S., de Campos Braz, L.M., Williams, H.T., Lewis, M.R., Frost, G. and Li, J.V., 2022. Improved quantitation of short-chain carboxylic acids in human biofluids using 3-nitrophenylhydrazine derivatization and liquid chromatography with tandem mass spectrometry (LC-MS/MS). Journal of Pharmaceutical and Biomedical Analysis 221: 115060. https://doi.org/10.1016/j.jpba.2022.115060
van der Hee, B. and Wells, J.M., 2021. Microbial regulation of host physiology by short-chain fatty acids. Trends in Microbiology 29: 700-712. https://doi.org/10.1016/j.tim.2021.02.001
van der Schoot, A., Helander, C., Whelan, K. and Dimidi, E., 2022. Probiotics and synbiotics in chronic constipation in adults: a systematic review and meta-analysis of randomized controlled trials. Clinical Nutrition 41: 2759-2777. https://doi.org/10.1016/j.clnu.2022.10.015
Van-Wehle, T. and Vital, M., 2024. Investigating the response of the butyrate production potential to major fibers in dietary intervention studies. npj Biofilms and Microbiomes 10: 63. https://doi.org/10.1038/s41522-024-00533-5
Wang, C.Y., Chen, Y.W., Tain, Y.L., Chang, S.K.C., Huang, L.T., Hsieh, C.W. and Hou, C.Y., 2020. Fast quantification of short-chain fatty acids in rat plasma by gas chromatography. Journal of Food Science 85: 1932-1938. https://doi.org/10.1111/1750-3841.15172
Wickham, H., 2016. Package âggplot2â: elegant graphics for data analysis, Vol. 10. Springer-Verlag, New York.
Williams, E.A., Stimpson, J., Wang, D., Plummer, S., Garaiova, I., Barker, M.E. and Corfe, B.M., 2009. Clinical trial: a multistrain probiotic preparation significantly reduces symptoms of irritable bowel syndrome in a double-blind placebo-controlled study. Alimentary Pharmacology & Therapeutics 29: 97-103. https://doi.org/10.1111/j.1365-2036.2008.03848.x
Wu, Y., Li, Y., Zheng, Q. and Li, L., 2024. The efficacy of probiotics, prebiotics, synbiotics, and fecal microbiota transplantation in irritable bowel syndrome: a systematic review and network meta-analysis. Nutrients 16. https://doi.org/10.3390/nu16132114
Xiong, R.G., Zhou, D.D., Wu, S.X., Huang, S.Y., Saimaiti, A., Yang, Z.J., Shang, A., Zhao, C.N., Gan, R.Y. and Li, H.B., 2022. Health benefits and side effects of short-chain fatty acids. Foods 11. https://doi.org/10.3390/foods11182863
Yilmaz-Ersan, L., Ozcan, T. and Akpinar-Bayizit, A., 2020. Assessment of socio-demographic factors, health status and the knowledge on probiotic dairy products. Food Science and Human Wellness 9: 272-279. https://doi.org/10.1016/j.fshw.2020.05.004
Yoon, G., Gaynanova, I. and Müller, C.L., 2019. Microbial networks in SPRING â semi-parametric rank-based correlation and partial correlation estimation for quantitative microbiome data. Frontiers in Genetics 10. https://doi.org/10.3389/fgene.2019.00516
Zhao, G., Nyman, M. and Jönsson, J.A., 2006. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomedical Chromatograph 20: 674-682. https://doi.org/10.1002/bmc.580





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}