
Abstract
Predicting probiotic strain properties that translate into beneficial effects in the host is challenging and could be considered the holy grail in developing new probiotics. One approach is to selectively evolve well-studied probiotic strains known to elicit beneficial effects in clinical studies. In this study, Limosilactobacillus reuteri DSM 17938, among the world’s most studied probiotic strains, constituted the origin from which the novel strain L. reuteri BG-R46® (DSM 32846) was obtained by exposure to bile, a major stressor of the gastrointestinal tract. The bioactivity of DSM 32846 in preclinical models is documented elsewhere; this work adds crucial strain characteristics, genomic and phenotypic safety profiles, and safety and tolerability in a randomised human study. DSM 32846 was compared to DSM 17938 in terms of tolerance to bile and synthetic gastric juice, with DSM 32846 showing increased bile tolerance and slightly reduced tolerance to gastric pH. Morphological examination by scanning electron microscopy revealed high abundance of extracellular membrane vesicles on the surface of DSM 32846, a previously described bioactive feature of the strain. In addition, the activity of 5′-nucleotidase, an adenosine-producing enzyme expressed on the bacterial cell surface and membrane vesicles, was increased in large-scale production batches of DSM 32846 compared to DSM 17938. Phenotypic safety assessment showed that DSM 32846 does not produce any of the evaluated biogenic amines, produces D- and L-lactate with a ratio typical for the species, and lacks haemolytic activity. The minimum inhibitory concentration profile for antibiotics did not raise any safety concerns. A genome safety assessment revealed no antibiotic resistance and virulence genes. DSM 32846 was found safe for human consumption with no differences from the control group in any of the evaluated parameters. In conclusion, L. reuteri DSM 32846 is a safe strain that shows interesting differences to its parent strain DSM 17938.

Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00101
1 Introduction
The basis for probiotic products is the included bacterial strains, and there are several requirements that must be met for a strain to be called probiotic (Hill et al., 2014). Binda and colleagues further clarified criteria required to use the terminology correctly stating that probiotic strains must be sufficiently characterised, safe for the intended use, supported by at least one positive human clinical trial conducted according to generally accepted scientific standards, and alive in the product at an efficacious dose throughout shelf life (Binda et al., 2020). Further, Dunne and colleagues have described criteria to assist in the selection of probiotic strains (Dunne et al., 2001). Within the latter criteria are tolerance to processing and stress related to passage through the gastrointestinal tract, adherence in the intestine, influence and modulation of immune responses. In addition to these general probiotic characteristics, there are species- and strain-specific properties that are important for the beneficial effects elicited by the probiotics. Limosilactobacillus reuteri DSM 17938 has been shown to carry the adenosine-producing enzyme 5′-nucleotidase, which likely plays an important role in the anti-inflammatory properties of the strain (He et al., 2017). Further, DSM 17938 affects gut motility, improves mucosal integrity and barrier function, inhibits pain signalling, and exhibits antimicrobial properties through the secretion of 3-hydroxipropionaldehyde (commonly known as reuterin) (Burgos et al., 2015; Chung et al., 1989; Karimi et al., 2018). Although selection of strains with improved activity towards any of the bioactivity traits is difficult, traits related to tolerance can be used as tools for generation of adapted strains. By exposing strains to stressful environments related to the mentioned traits, one can obtain naturally adapted mutants with improved properties (Foster, 2007).
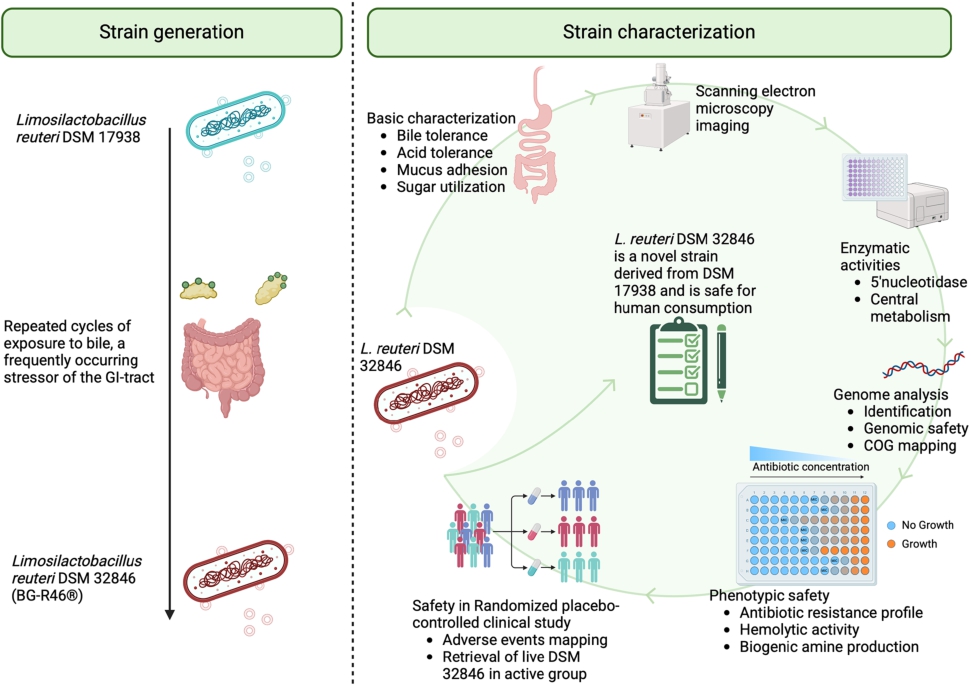
L. reuteri DSM 17938 is among the most clinically well-studied probiotic strains in the world and has, for example, been shown to alleviate infantile colic (Gutiérrez-Castrellón et al., 2017; Reis Buzzo Zermiani et al., 2021; Szajewska et al., 2014; Sung et al., 2017) and reduce diarrhoeal symptoms (Mu et al., 2018; Sun et al., 2023). DSM 17938 has many beneficial properties and is well-documented both clinically and preclinically but has a lower bile tolerance compared to other L. reuteri strains (Sendelius et al., 2023). A high tolerance to the stressful conditions in the upper GI tract could potentially be linked to increased activity and probiotic activity, and thus we speculated that a strain with the properties of DSM 17938 and having an improved bile tolerance could have an even greater probiotic efficacy. By exposing the strain to bile, a new strain named DSM 32846 was obtained (Pang et al., 2022). Changes such as improved bile tolerance rarely occur in isolation and Galhardo and colleagues emphasise how microbial evolution is achieved through stressful environments, and that the initial stress response in cells that are poorly adapted to the environment is the occurrence of random mutations (Galhardo et al., 2007). Mutations and possible alterations in the regulation of the genome may explain why the two strains differ. We have previously described that extracellular membrane vesicles from DSM 32846 have increased bioactivity in several models (Pang et al., 2022) and the strain increased both adenosine and inosine levels in the cecum of scurfy mice (Liu et al., 2023). However, the basic properties and safety of DSM 32846 have not been described previously. Therefore, this paper aimed to describe the characteristics of DSM 32846 in depth and elucidate whether any other basic property of the strain has been altered in the strain development procedure. Furthermore, we aimed to evaluate the safety of the strain in a human safety study.
2 Materials and methods
Bacterial strains and culture conditions
Limosilactobacillus reuteri strain DSM 32846 (commercial name L. reuteri BG-R46®, a trademark of BioGaia AB) is an evolved strain obtained by repeated bile stress of the well-characterised probiotic strain L. reuteri DSM 17938 (Pang et al., 2022). DSM 17938 was previously obtained after curation of antibiotic resistance-carrying plasmids from L. reuteri ATCC 55730, a strain derived from breast milk (Rosander et al., 2008; Wolf et al., 1995). DSM 32846, DSM 17938, as well as L. reuteri strains ATCC PTA 4659 (Sendelius et al., 2023), ATCC PTA 5289 and ATCC PTA 6475 were used in this study and were all kind gifts from BioGaia (Stockholm, Sweden).
Genomic analysis
Whole-genome sequencing
The genome of DSM 32846 was obtained by hybrid assembly of Nanopore and Illumina reads. The Unicycler pipeline v0.4.8 in hybrid mode was used to obtain de novo assemblies. All dependencies for Unicycler were installed in a Conda environment. The dependency programs include SPAdes v3.13.0, racon v1.4.1, bowtie2 v2.3.5.1, and pilon v1.23. The hybrid assemblies were annotated using Prokka v1.14.5 (https://github.com/tseemann/prokka). Total genomic DNA was extracted from an overnight culture in DeMan Rogosa and Sharpe (MRS, Sigma Aldrich, Saint Louis, MO, USA) broth. The culture was centrifugated for 10 min at 4,500 rpm at 4 °C, after which the pellet was washed once with phosphate buffered saline pH 7.4 (PBS) to remove carry-over contaminants. DNA for Illumina short-reads sequencing was extracted using the NucleoSpin Soil kit (740780.50, Macherey-Nagel) as described by the manufacturer with SL2 lysis buffer and Sx enhancer. Cells were bead beated at 5.5 m/s for 60 s twice in a FastPrep-24 Instrument (MP Biomedicals), with incubation on ice for 5 min in between the two bead-beating bursts. DNA quality was evaluated using Tapestation 4200 with Genomic DNA ScreenTape and reagents (Agilent), and quantification was performed using Quant-iT dsDNA BR Assay Kit (ThermoFisher Scientific). Libraries for sequencing were prepared using Covaris S220 Focused-ultrasonicator (Covaris), fragmented to average 550-bp insert size, and the TruSeq DNA PCR-free Library Preparation kit (20015963 and 20015949, Illumina). Libraries were quantified using Quant-iT dsDNA HS Assay Kit (ThermoFisher Scientific) and sequenced on an Illumina Miseq instrument using MiSeq Reagent Kit v3, 600 cycles.
Large amounts of high-quality DNA for Nanopore long-reads sequencing were obtained with a modified version of the Marmur procedure (Marmur, 1961). Cells were suspended in Tris-EDTA buffer (20 mM Tris HCl pH 8.0, 2 mM EDTA) and lysed with lysozyme (20 mg/ml) and SDS (2%, w/v) in the presence of proteinase K (80-200 μl/ml, Qiagen, Hilden, Germany). The extracted total DNA was purified by repeated extraction in phenol:chloroform:isoamyl alcohol (25:24:1, v/v) and chloroform:isoamyl alcohol (24:1, v/v), followed by precipitation by addition of 2 volumes of cold ethanol (99.5%, v/v) and spooling of the DNA on a glass rod. The DNA was washed with 70% (v/v) ethanol, dried at room temperature and resuspended in water overnight at 4 °C. DNA integrity and concentration were evaluated using Tapestation 4150 with Genomic DNA ScreenTape and reagents (Agilent) and Qubit 3.0 Fluorometer and Qubit dsDNA BR assay kit (ThermoFisher Scientific). Isolated DNA was prepared using Rapid barcoding kit (SQK-RBK004) following the manufacturer’s instructions (ONT) and sequenced on a ONT MinION device on a R9.4.1 flow cell (FLO-MIN106D). Base-calling was performed using ONT Guppy v. 4.2.2.
The genome sequence was deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank) and has the accession number CP169042 (NCBI RefSeq: GCF_041888795.1). The genome of the parental strain DSM 17938 (generated with the above-described method) was deposited with the accession number CP169045 (NCBI RefSeq: GCF_041888805.1). Both strains carry two plasmids, pLR580 and pLR584, and the sequences were deposited with the accession numbers CP169047 and CP169046 for DSM 17938, and CP169044 and CP169043 for DSM 32846.
The average nucleotide identity (ANI) value was calculated based on MUMmer using the JSpeciesWS ANI calculator (Richter et al., 2016).
Genome annotation and risk analysis
The genome annotations of the coding sequences (CDS) of the strains DSM 32846, DSM 17938 and ATCC PTA 6475 (NCBI RefSeq: GCF_000159475.2 (MM4-1a)) were obtained by subjecting the de novo assembled genomic DNA sequences in FASTA format to NCBI’s ‘Prokaryotic Genome Annotation Pipeline (PGAP)’, version 2024-04-27.build7426 (https://github.com/ncbi/pgap). The genome annotations of the L. reuteri strains JCM 1112 and SD2112 (same strain as ATCC 55730) are publicly available with NCBI accession numbers GCF_000010005.1 and GCF_000159455.2, respectively. To make the functional annotation comparable between the different strains, the CDSs and the genomic DNA sequences were further annotated by using eggnog-mapper (version 2.1.12, https://github.com/eggnogdb/eggnog-mapper) based on their ortholog and functional cluster database – EggNOG (v5). The mapping mode used for the analysis was ‘MMseqs2’ and the taxonomic level was set to ‘Bacteria’, while other parameters for eggnog-mapper were unchanged. The Clusters of Orthologous Genes (COG) identified by eggnog-mapper were compared across the different strains, with a special consideration for the ‘COG V’ category, namely ‘defence mechanisms’.
Detection of antibiotic resistance genes, genomic islands and virulence factors
To determine whether any antibiotic resistance genes are present in the strains, Resistance Gene Identifier (RGI v6.0.3, https://card.mcmaster.ca/analyze/rgi), developed based on the Comprehensive Antibiotic Resistance Database (CARD, version 3.2.9), was used with the selection criteria ‘Perfect and Strict hits only’. As a complement to CARD, ResFinder (v4.5.0, http://genepi.food.dtu.dk/resfinder) was used to identify potential antibiotic resistance genes.
IslandViewer (v4, http://www.pathogenomics.sfu.ca/islandviewer/) and MobileElementFinder (v1.0.3, https://cge.food.dtu.dk/services/MobileElementFinder/) were used to detect genomic islands (GI)/mobile genetic elements (MGEs) on the genomes. IslandViewer uses a set of different tools and databases to identify GIs as well as relevant antibiotic resistance genes and genes encoding virulence factors that may be located on the GIs. MobileElementFinder is another tool for identifying MGEs using a curated database and it can also assess antimicrobial resistance genes and virulence genes.
The above-mentioned assessments were conducted in June-July 2024 and the analysis was performed separately on the chromosomal and plasmid DNAs.
Phenotypic safety assessment
Determination of minimum inhibitory concentrations (MICs)
DSM 32846 was assessed for susceptibility to ampicillin, chloramphenicol, clindamycin, erythromycin, gentamicin, kanamycin, streptomycin, and tetracycline using a broth microdilution method according to guidelines from the European Food Safety Authority (EFSA) (EFSA FEEDAP Panel et al., 2018). The tests were performed at RISE Research Institutes of Sweden AB (Borås, Sweden) following ISO 10932:2010 (British Standards Institute Staff, 2010). Two-fold dilutions of the antibiotics were prepared, and bacterial cells were diluted in LSM broth (Klare et al., 2005). Fifty μl of the bacterial cells were added to each well (approximately 5 × 105 CFU/ml) and the microwell plates were incubated anaerobically at 37 °C for 48 h. Lacticaseibacillus paracasei ATCC 334 and Lactiplantibacillus plantarum ATCC 14917 were tested in tandem with DSM 32846 to verify the methodology, and they exhibited MICs within accepted ranges according to ISO 10932:2010 (British Standards Institute Staff, 2010). MIC values were compared with the cut-off values for L. reuteri strains defined by EFSA (EFSA FEEDAP Panel et al., 2018).
In addition, MIC measurement for chloramphenicol was performed using the Etest strip method (bioMérieux, Lyon, France) according to the manufacturer’s instructions. The antibiotic concentration gradient of the Etest strips was 0.016 to 256 μg/ml chloramphenicol. DSM 32846 was cultivated anaerobically at 37 °C on LSM agar supplemented with Tween 80 (1 g/l) to ensure optimal growth. Colonies from overnight cultures (22 to 26 h) were suspended in saline solution (0.9% NaCl, wt/vol). The cell suspension (3 × 108 CFU/ml) soaked in a sterile cotton swab was streaked with a plate rotation 60 degrees three times on LSM agar supplemented with Tween 80 (1 g/l). After drying the plate surface, Etest strips were applied to the agar medium. The Etest agar plates were incubated anaerobically at 37 °C for 48 h.
If the MIC value for an antibiotic was above the cut-off value, the genome sequence of DSM 32846 was manually investigated to find potential explanations. This was performed by comparisons with known determinants of elevated MICs as well as by review of the background information used to define the cut-off values (EFSA FEEDAP Panel et al., 2018).
Biogenic amines
Production of the biogenic amines histamine, tyramine, cadaverine and putrescine was investigated for DSM 17938 and DSM 32846. The analyses were performed at RISE Research Institutes of Sweden (Borås, Sweden). After culturing bacteria overnight in MRS broth (Merck, Darmstadt, Germany) at 37 °C, the culture was centrifuged at
Production of d- and l-lactate
The production of d- and l-lactate was analysed at RISE Research Institutes of Sweden (Borås, Sweden). Supernatants from overnight cultures in MRS broth at 37 °C were obtained after centrifugation at
Haemolytic activity
Assessment of haemolytic activity was performed by cultivation on Brain Heart Infusion (BHI, Merck) agar supplemented with 5% sheep blood. DSM 32846 was inoculated onto the agar plates from frozen stocks by gently tapping the inoculation loop onto the agar. Staphylococcus aureus S151:2 (α-haemolytic) (Rosengren et al., 2010) and Streptococcus sanguis Kx126C2 (β-haemolytic, human origin, strain collection at Department of Molecular Sciences, Swedish University of Agricultural Sciences) were used as positive controls. Plates containing DSM 32846 were incubated anaerobically at 37 °C for 72 h and plates containing S. aureus and S. sanguis were incubated aerobically at 37 °C for 24 h.
Physiological characterisation
Morphology assessment by scanning electron microscopy
The morphological appearance of DSM 32846 was observed using scanning electron microscopy. The analysis was performed at the Center for Cellular Imaging, Core Facilities, Sahlgrenska Academy, University of Gothenburg. Bacteria were cultivated, lyophilised and visualised as described by Pang et al. (2022) and Ermann Lundberg et al. (2024). Briefly, lyophilised bacteria were rehydrated in PBS for 20 min, coverslips coated with 0.1% poly-L-lysine for 10 min followed by drying for 5 min, then incubated in cell suspension for 30 min at room temperature. The cells were washed and incubated in 2.5% glutaraldehyde in 0.1 M PIPES for 30 min at room temperature. After post-fixation in 1% osmium tetroxide in 0.1 M PIPES for 1 h at 4 °C in the dark, the samples were dehydrated successively in a series of ethanol solutions of increasing ethanol concentration (35-100%). The samples were suspended in Hexamethyldisilazane solution, air dried and sputtered with gold (Emitech, Taunusstein, Germany) before visualisation with a Zeiss Gemini 450 II scanning electron microscope (Carl Zeiss, Toronto, ON, Canada).
Carbohydrate utilisation
To determine the ability to utilise carbohydrates, the API 50 CHL kit (bioMérieux) was used according to the manufacturer’s instructions. Bacterial colonies on MRS agar (Oxoid) were suspended in API 50 CHL medium to a cell density corresponding to McFarland Standard No. 2 (bioMérieux). The suspended cells were inoculated into a well-type plate provided by the manufacturer and then incubated at 37 °C for 48 h. The ability to utilise carbohydrates was assessed by determining colour changes.
API ZYM
The enzymatic activity profile was determined using the API ZYM kit (bioMérieux) according to the manufacturer’s protocol. Briefly, DSM 17938 and DSM 32846 were grown overnight at 37 °C in MRS broth (Merck). The bacteria were harvested by centrifugation and the pellet was resuspended in sterile saline to a turbidity of 5.0-6.0 McFarland. The resuspended cells were added to a well-type plate provided by the manufacturer and the plate was incubated at 37 °C for 4 h. Then, one drop of each of ZYM-A and ZYM-B reagents (bioMérieux) was added to each well. After 5 min at room temperature, the colour changes were assessed according to the manufacturer’s instructions and used to obtain an enzyme activity profile.
Reuterin production
The production of reuterin was analysed by using a colorimetric method that allows the produced reuterin (3-hydroxy-propinoaldehyde) to react with the reagent 2,4-dinitrophenylhydrazine (Rosander et al., 2008). After the bacteria had grown on MRS agar (Oxoid) for 48 h, the plates were overlaid with 500 mM glycerol (1% agar) followed by an incubation for 30 min at 37 °C. Then 5 ml 2,4-dinitrophenylhydrazine (0.1% in 2 M HCl) was added and the plates were incubated for 3 min at room temperature. Thereafter the solution was poured off and 5 ml of 5 M KOH was added. The production of reuterin resulted in reddish-brown zones around the colonies with sizes that reflected the magnitude of the production.
5′-nucleotidase activity
5′-nucleotidase (5′NT) activity of freeze-dried large-scale batches of DSM 17938 and DSM 32846 was measured using the 5′-nucleotidase Assay kit (CrystalChem, Elk Grove Village, IL, USA). Briefly, 0.2 g of freeze-dried bacteria (obtained by BioGaia AB) was first suspended in 10 ml water. After incubation at room temperature for 20 min, the suspensions were centrifuged at 4,000 g for 10 min. 10 μl of the supernatants were added to a 96-well plate containing 180 μl of reagent CC1 per well. After 5 min incubation at 37 °C, 90 μl of reagent CC2 was added and the quinone dye was measured at 550 nm after 3, 5, 9 and 13 min using a plate reader. The change in absorbency values was used for calculations of 5′NT activities according to the manufacturer’s instructions.
Acid tolerance
Survival at pH 2.0 in synthetic gastric fluid without digestive enzymes was analysed according to Wall et al. (2007). Duplicate samples were taken after 50 and 90 min. The survival rate was calculated as the percentage of bacteria surviving acid stress relative to the number of bacteria at the start. The assay was performed in biological triplicates.
Bile tolerance
The bile tolerance was analysed in two different ways. (i) Both DSM 32846 and DSM 17938 were grown in MRS broth (Oxoid) for 16-20 h and centrifuged for 10 min at
Bile salt hydrolase assay
Bile salt hydrolase (BSH) activity was analysed by adding 10 μl of overnight cultures in MRS broth (Merck) on MRS agar plates (Merck) supplemented with 0.5% (w/v) sodium salt of glycodeoxycholic acid (GDCA; Sigma-Aldrich) and 0.37 g/l CaCl2. The plates were incubated anaerobically at 37 °C for 72 h. Bacterial spots surrounded by precipitation zones (opaque halos) were considered BSH positive (Hernández-Gómez et al., 2021). The diameter of the zones was measured. MRS agar plates without supplementation were used as controls.
Adhesion to mucus
The mucus attachment assay is a modification to the original method described by Roos et al. (2000). The modified method has been described in full in Ermann Lundberg et al. (2024).
Briefly, cells were removed from porcine small intestine mucus by centrifugated at
Clinical safety and tolerability
A randomised, double-blind, placebo-controlled clinical study was performed on healthy volunteers. The study had a parallel-group design, and the objective was to evaluate safety and tolerability of L. reuteri DSM 32846. The study was carried out by Clinical Trial Consultants AB in Uppsala, Sweden in October to December 2021. The study was approved by the independent Swedish Ethical Review Authority (Dnr 2021-02923) and registered at ClinicalTrials.gov (NCT05044949) prior start of the study. The study was conducted in accordance with ethical principles originating in the Declaration of Helsinki and was compliant with International Conference of Harmonization (ICH)/Good Clinical Practice (GCP) E6 (R2) guidance. Applicable parts of the European Union Clinical Trials Directive and local regulatory requirements were followed. All participants gave verbal and written informed consent before being included in the study and study-specific assessments were performed. Inclusion and exclusion criteria are provided in Supplementary Tables S1 and S2.
The study was comprised of one screening visit and four study visits: at baseline (day 1), day 7, day 14 and day 28, and a final follow-up telephone call was performed 14 days after day 28 to collect adverse events and use of concomitant medications (Supplementary Figure S1).
36 healthy subjects (28 females) with a mean age of 33.9 ± 13.6 years and a mean BMI of 23.7 ± 2.2 kg/m2, were included in the study. Subjects were randomly assigned at a ratio of 1:1:1, to receive low dose (1 × 108 CFU), high dose (1 × 1010 CFU) or placebo product (
The subjects were instructed to consume the study product orally once a day in the morning together with at least one glass of water, milk or yogurt, but not juice or carbonated soft drinks. To be included in the per protocol set, >80% of the study product should have been consumed according to instructions without any major deviations that were judged to compromise the analysis of the data. All protocol violations were classified as major or minor prior to database lock.
Vital signs were measured in terms of systolic and diastolic blood pressure and pulse rate.
Physical examination included general appearance including skin, auscultations of lungs and heart, and by abdomen palpation of liver and spleen. Blood samples were collected through venous puncture and in vitro assessments of clinical safety were evaluated. This included measurement of chemical parameters such as alanine aminotransferase, alkaline phosphatase, aspartate aminotransferase, bilirubin, creatinine and non-fasting glucose. Haematological parameters including haematocrit, haemoglobin (Hb), platelet count, and white blood cell count with differential count including leukocytes, lymphocytes, monocytes, neutrophils, eosinophils, and basophils, were also evaluated. Follicle stimulating hormone (FSH), serum and urine pregnancy tests were conducted in females. To evaluate GI symptoms the gastrointestinal symptom rating scale (GSRS) was used, which is comprised of 15 items that cluster into five symptom clusters: reflux, abdominal pain, indigestion, diarrhoea and constipation (Svedlund et al., 1988). The absolute changes in GSRS survey items, subscores for each symptom cluster, and total scores between baseline and final visit were analysed using Wilcoxon Rank Sum test to investigate any differences between the study arms.
Stool samples were collected at baseline and after the treatment period on day 28 for analysis of the presence of viable DSM 32846. The samples, still blinded, were analysed for viable DSM 32846 by using the following method: Samples were thawed at room temperature and 0.1 g was suspended in 0.9 ml PBS and then added to 9 ml MRS broth. After incubation at 37 °C for 24 h, 100 μl of the enrichment culture was streaked onto Rogosa agar (Merck) supplemented with 50 μg/ml vancomycin and 2 μg/ml ampicillin (Romani Vestman et al., 2013). The plates were incubated anaerobically at 37 °C for 24 h before further analysis. A mix of bacterial colonies on the plate was collected with a 1-μl loop and thereafter suspended in 100 μl sterile water. The presence of DSM 32846 was examined by PCR analysis using DreamTaq Green PCR Master Mix (Thermo Fisher) and strain-specific primers FwdR46p and RevR46p (see below; 0.4 μM of each). Bacterial suspension (0.5 μl) was added to the PCR mix and the PCR reaction was performed by running the program 95 °C for 7 min; 35 cycles of 95 °C for 30 s, 62 °C for 30 s, and 72 °C for 30 s; and 72 °C for 10 min. The PCR products were separated and visualised by standard agarose gel electrophoresis. A pure culture of DSM 32846 was used as positive control and L. reuteri DSM 17938 and ATCC PTA 6475 were used as negative controls.
The primers FwdR46p (CACAAACGAAATACTATTCCACGG) and RevR46p (TTGCGATGAGTGGCTTTGG) were designed to amplify parts of the unique region of one of the duplicated insertion sequence elements in DSM 32846. The optimization procedure showed that the primers resulted in a PCR product (224 bp) from DSM 32846 but no product from L. reuteri strains DSM 17938, ATCC PTA 4659, ATCC PTA 5289 and ATCC PTA 6475 (Supplementary Figure S2).
3 Results
Whole genome sequence analysis
Limosilactobacillus reuteri ATCC 55730 (also designated SD2112), the parental strain of DSM 17938, has been identified as L. reuteri subsp. kinnaridis (Li et al., 2021). The genome of L. reuteri DSM 32846 was sequenced, resulting in one contig with a total size of 2,225,488 base pairs. Analysis of the genome sequence identified the strain as L. reuteri subsp. kinnaridis with an ANI value of 98.90% between DSM 32846 and the strain L. reuteri AP3, which is described as the type-strain for L. reuteri subsp. kinnaridis (Li et al., 2021). The ANI values between DSM 32846 and the ancestor strains ATCC 55730 and DSM 17938 were both 99.99% and ANI values between DSM 32846 and some L. reuteri subsp. reuteri strains were above 96.00% (Supplementary Table S3). A comparative genomic assessment between DSM 32846 and DSM 17938 revealed that there are two insertion sequence elements that have been duplicated and inserted in the genome of DSM 32846 (positions 1102637-1103717 and 1809646-1810722).
Minimum inhibitory concentrations of antibiotics for DSM 32846
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00101
Genomic risk assessment
The annotation from the PGAP pipeline revealed that, on the chromosome, DSM 32846 has a total number of CDS of 2115, which is in the same range as DSM 17938, ATCC PTA 6475, JCM 1112, and SD2112, all of which range from 2004-2181 CDSs on the chromosome. The CDSs identified on the plasmids were 18 on pLR584 and 5 on pLR580, which are the same as on the plasmids of DSM 17938. The eggnog-mapper analysis showed that the overall COG composition of DSM 32846 was very similar to the other related strains on both the chromosomes and plasmids. The COG category ‘V’, which is a functional cluster of orthologs with defence mechanisms, was also found to be very similar to the other strains (Supplementary Table S4). No category ‘V’ COG was identified on the plasmids. In conclusion, from a functional composition point of view, there is no safety concern for DSM 32846.
Analysis conducted using RGI (CARD database) revealed a match to resistance gene ‘vanT gene in vanG cluster’, but only with 33.6% alignment, and this was not further identified by ResFinder, suggesting an extremely low possibility of being linked to antibiotic resistance. IslandViewer identified 26 GIs from DSM 32846, but no antibiotic resistance genes or genes encoding virulence factors were found within or outside the GI regions. MobileElementFinder identified 33 MGEs (12 with high prediction quality fulfilling three criteria: alignment coverage > 95%, sequence identity > 90%, and truncation < 30 nt) with no resistance genes or genes encoding virulence factors within or outside the MGEs. In conclusion, the genomic risk analysis showed that DSM 32846 had no safety concerns when used as a probiotic.
Determination of and explanations of minimum inhibitory concentrations (MICs)
The minimum inhibitory concentrations (MIC) of the antibiotics ampicillin, chloramphenicol, clindamycin, erythromycin, gentamicin, kanamycin, streptomycin, and tetracycline were tested by broth microdilution for DSM 32846 (Table 1). The MICs for clindamycin, erythromycin, gentamicin, kanamycin, and streptomycin were all below or equal to the cut-off values, but the MICs for ampicillin, chloramphenicol, and tetracycline were above the cut-off values defined by EFSA. However, using the Etest strip method, the MIC value for chloramphenicol was determined to be 4 μg/ml, which is equal to the cut-off value. As described above, no antibiotic resistance genes could be found in the genome sequence and to explain the elevated MIC values, the genome sequence of DSM 32846 was manually investigated. Point mutations in the penicillin-binding proteins of DSM 17938 have previously been linked to the elevated MIC for ampicillin (Rosander et al., 2008) and the same point mutations were found in DSM 32846 (PBP1A – GenBank accession number XGD31112; PBP2X – accession number XGD30840). In the search for an explanation for the elevated MIC for tetracycline, the rpsJ gene encoding ribosomal protein S10 was identified. The genes of both DSM 32846 (XGD29732) and DSM 17938 (XGD31779) differ from sensitive strains such as ATCC PTA 6475 at two adjacent positions. These result in a change from amino acid residues KF (positions 57-58) in ATCC PTA 6475 (in GenBank designated MM4-1A; Accession EGC14332) to IS in DSM 32846 and DSM 17938. A similar change of rpsJ in Neisseria gonorrhoeae results in increased tolerance to tetracycline (Hu et al., 2005). This strongly suggests that the exchange of amino acid residues in ribosomal protein S10 in DSM 32846 and DSM 17938 is the cause of the increased tolerance of tetracycline. Both rpsJ and the genes encoding the penicillin-binding proteins are housekeeping genes located on the chromosome without adjacent mobile genetic elements. The risk for gene transfer is therefore negligible.
No potential explanation for the elevated MIC for chloramphenicol could be identified in the genome of DSM 32846. However, a review of the background information used for defining the EFSA cut-off values revealed that the value for chloramphenicol is likely to be incorrectly defined. The background is that EUCAST used the information from (Egervärn et al., 2007) to build its MIC database for L. reuteri (https://mic.eucast.org/search/, accessed March 4, 2025). This database has further been used for defining the cut-off values in the document Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance (EFSA FEEDAP Panel, 2012). This document clearly states that a microdilution method should be used for the MIC analysis. However, the EUCAST MIC database contains a mix of values obtained by both microdilution and Etest, and the value for chloramphenicol has been obtained from Egervärn et al. (2007) that used Etest for chloramphenicol. Based on our analysis of the cut-off value using both microdilution (8 μg/ml) and Etest (4 μg/ml) a correct cut-off value probably should be 8 μg/ml.
Analysis of biogenic amines and d,l isomers of lactate
Biogenic amine production was evaluated because the production of the biogenic amines histamine, tyramine, cadaverine, and putrescine may be a potential health concern (Ruiz-Capillas and Herranz, 2019). According to HPLC analysis, neither DSM 32846 nor DSM 17938 produced any of the biogenic amines histamine, tyramine, cadaverine, and putrescine neither in MRS broth nor in MRS broth supplemented with l-histidine, l-lysine, l-ornithine or l-tyrosine (Supplementary Table S5).
Many lactic acid bacteria produce both the l- and d-isomers of lactate (Hammes and Hertel, 2006). d-Lactate is not a highly toxic compound, but it is a metabolite that under specific circumstances can cause health problems (Petersen, 2005). Quantitative analysis of lactic acid production showed that DSM 32846 produced both l-lactate (69%) and d-lactate (31%). The ratio of l- to d-lactate is consistent with the results for DSM 17938 (l-lactate: 71% and d-lactate: 29%) a strain with a solid safety profile (Fatheree et al., 2017; Mu et al., 2018; Urbańska and Szajewska, 2014) and has in addition been shown not to cause an increase in urinary d-lactate after 28 days of administration to healthy infants (Papagaroufalis et al., 2014). Furthermore, no elevation of d-lactate was detected in serum of healthy infants given L. reuteri ATCC 55730 (the parental strain of DSM 17938) for 12 months (Connolly et al., 2005).
Analysis of haemolytic and enzymatic activities
Haemolytic activity results in lysis of red blood cells and thus destruction of haemoglobin. Beta-haemolytic activity is a representative virulence factor, and the absence of this activity is a criterion for the selection of novel probiotic strains. The ability of DSM 32846 to cause haemolysis was therefore evaluated and the result showed that the strain did not exhibit haemolytic activity. DSM 32846 did not show clear zones on BHI agar containing sheep blood, while the positive controls S. aureus S151:2 and S. sanguis Kx126C2 showed beta- and alpha-haemolysis, respectively (Supplementary Table S6).
A semiquantitative assessment of enzymatic activities was performed using the API ZYM kit. The results are presented in Supplementary Table S7. The following enzymatic activities were detected for DSM 32846 and DSM 17938: esterase, leucine arylamidase, acid phosphatase, naphthol-AS-BI phosphohydrolase, α-galactosidase, β-galactosidase and α-glucosidase. The enzymatic profile was similar to other L. reuteri strains (Ali et al., 2023; Lee et al., 2021). An example of a potential enzymatic activity that may cause concern and that would need to be further investigated are glucuronidases, which can release toxins and mutagens and thus increase the risk of cancer (Humblot et al., 2007; Kim and Jin, 2001). However, no production of β-glucuronidase by DSM 32846 or DSM 17938 was detected.
Physiological characterisation
Carbohydrate utilisation, reuterin production and 5′-nucleotidase activity
The ability of both DSM 32846 and DSM 17938 to ferment different carbohydrates was analysed (Supplementary Table S8), and the strains were found to utilise ten of the 50 substrates evaluated. The fermentation profile was consistent with that of other L. reuteri subsp. kinnaridis strains (Li et al., 2021).
Furthermore, the production of reuterin, a potent antimicrobial substance, was evaluated. DSM 32846 produced reuterin of the same order of magnitude (14 mm zones) as DSM 17938.
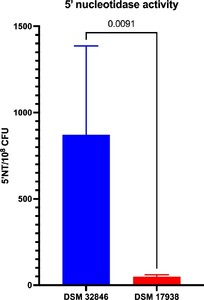
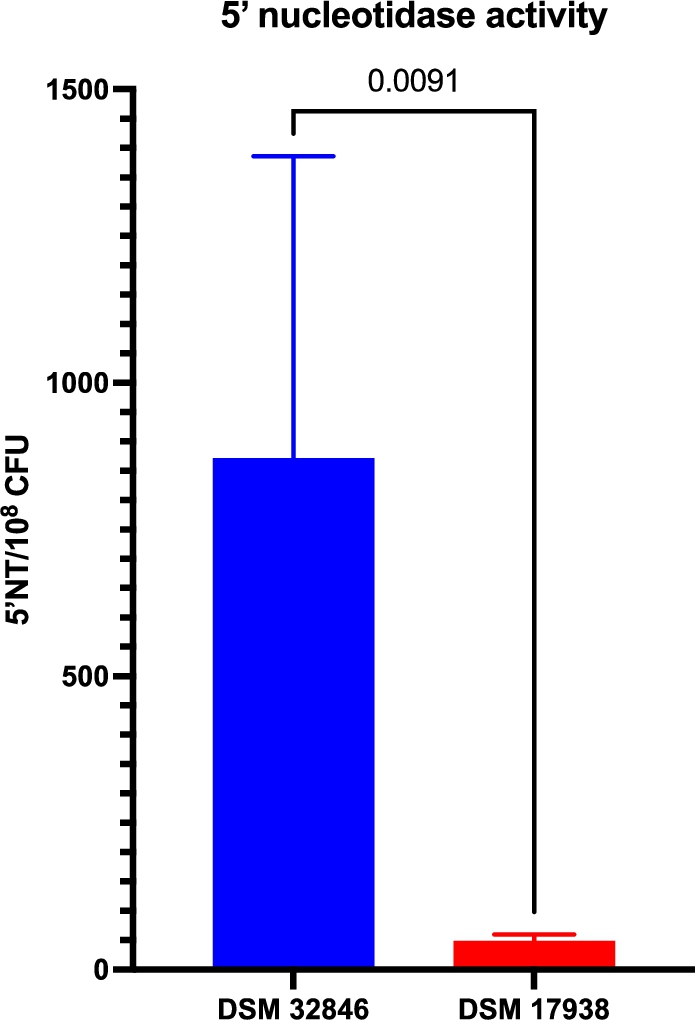
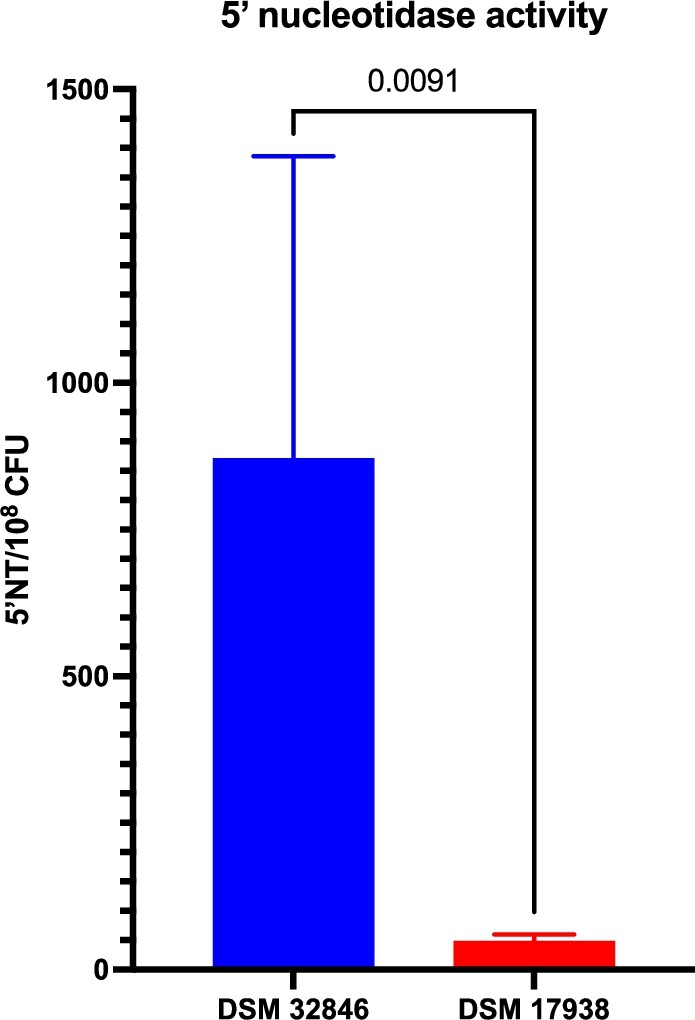
5′-nucleotidase activity has been described as a major contributor to the bioactivity of DSM 32846 and DSM 17938 and has previously been shown to be higher in the former strain (Liu et al., 2023; Pang et al., 2022). In this study, we evaluated supernatants of industrially produced freeze-dried culture powders with DSM 32846 or DSM 17938 and in agreement with previously reported elevated 5′NT activity in DSM 32846, the industrially produced DSM 32846 supernatant expressed higher activity compared to DSM 17938 (
Secreted 5′-nucleotidase activity in supernatants from industrially produced batches of DSM 32846 (
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00101
Scanning electron microscopy
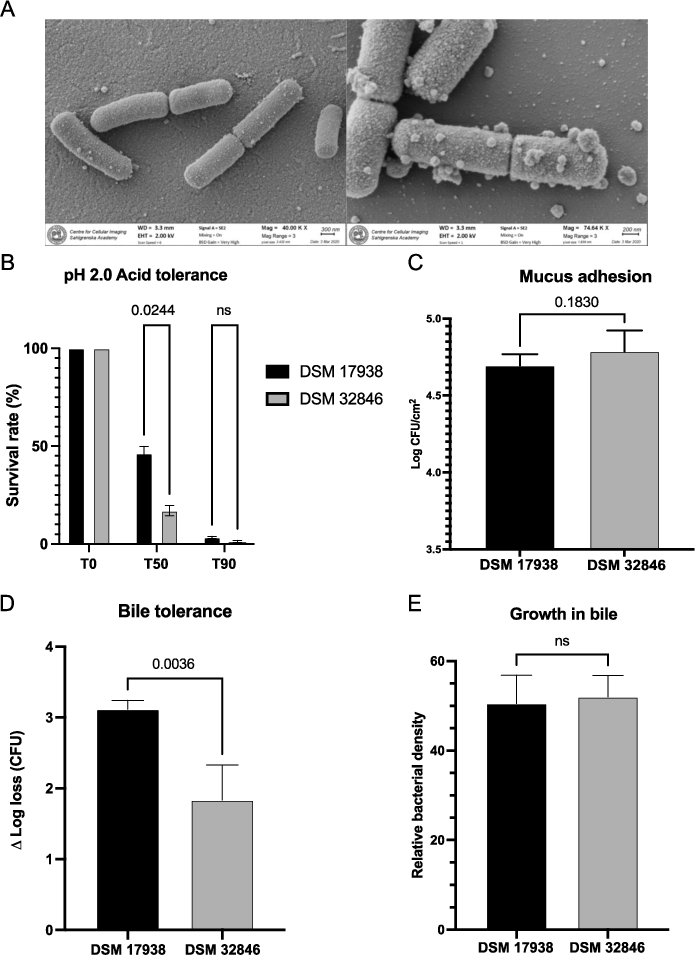
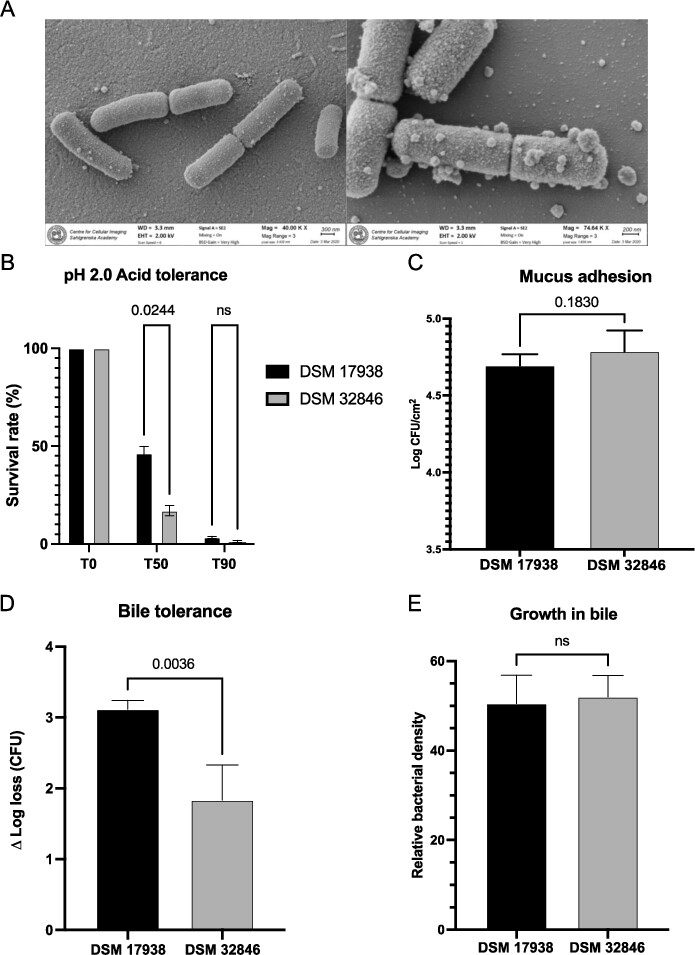
SEM images showed that DSM 32846 is rod-shaped with an estimated size of 1.2-2.2 μm (Figure 2A). Attached on the surface and in the surrounding area extracellular membrane vesicles of varying size were present.
(A) scanning electron microscopy imaging of DSM 32846 was performed on freeze-dried culture powder. The bacteria were fixed on 0.1% poly-l-lysine coated coverslips, gold sputtered and imaged with a Zeiss Gemini 450 II scanning electron microscope. (B) acid tolerance at pH 2.0 was evaluated for DSM 32846 and DSM 17938. The bacteria were suspended in synthetic gastric juice at pH 2.0 and then incubated at 37 °C. Samples were taken at 0, 50 and 90 min. Survival rates were calculated after performing viable count analysis of the samples. (C) mucus adhesion was evaluated for L. reuteri DSM 32846 and compared to DSM 17938. The strains were allowed to adhere to mucus-coated microtiter wells, after which they were released by trypsin treatment and CFU determined by plate count analysis. (D) bile tolerance of L. reuteri DSM 32846 was assessed and compared with DSM 17938. The bacteria were suspended in MRS supplemented with 0.5% (w/v) porcine bile, incubated at 37 °C and samples were taken at 0 and 180 min. The survival rate was calculated after performing viable count analysis of the samples. (E) growth in bile was evaluated for DSM 17938 and DSM 32846 where MRS broth supplemented with 0.2% bovine bile was used as substrate as the strains were grown in a microtiter plate and OD was measured. All data is presented as mean with SD and t-tests were used for statistical comparisons.
Citation: Beneficial Microbes 2026; 10.1163/18762891-bja00101
Acid tolerance
Probiotic strains taken orally should preferably survive passage through the stomach with its acidic pH. The survival of DSM 32846 and DSM 17938 in synthetic gastric juice with pH 2.0 was therefore analysed (Figure 2B). DSM 32846 had a slightly lower survival rate after 50 min of exposure compared to DSM 17938, but the two strains were in the same range after 90 min. Compared to most probiotics, both strains survived relatively well, as many species are very sensitive to pH 2 (Ko et al., 2022).
Mucus adhesion
Adhesion to mucus is considered an important property of probiotic strains as it allows the bacteria to get closer to the underlying intestinal host cells and is also a prerequisite for colonisation (long-term or transient). Therefore, the mucus-adhesion capacity of DSM 32846 was compared with DSM 17938. There was no difference in the binding capacity of the two strains, and the average number of adhered bacteria was 4.9 × 104 cells/cm2 for DSM 17938 and 6.1 × 104 cells/cm2 for DSM 32846 (Figure 2C).
Bile tolerance
L. reuteri DSM 32846 was selected by exposure to bile and the strain was found to be more tolerant to bile (0.5% porcine) compared to the parental strain DSM 17938. The average Δ log loss after 180 min of bile exposure was 1.8 for DSM 32846 and 3.1 for DSM 17938 (Figure 2D), showing that DSM 32846 tolerates exposure to bile to a greater extent. The bile tolerant phenotype was stable for more than 10 passages. However, methodological variance in bile tolerance assays has a major impact on bacterial survival, and to broaden the concept of bile tolerance, we also evaluated growth of DSM 32846 and DSM 17938 in MRS supplemented with bile. The strains were not shown to differ in this assay (Figure 2E).
Analysis of BSH activity
Bile salt hydrolases (BSH) catalyse the degradation of bile salts. BSH activity was analysed for DSM 32846 and the three well-known probiotic L. reuteri strains DSM 17938, ATCC PTA 5289 and ATCC PTA 6475. The two strains ATCC PTA 5289 and ATCC PTA 6475 showed activity against glycodeoxycholic acid (GDCA; precipitation zone diameters in Supplementary Table S9). DSM 32846 and DSM 17938 did not grow on MRS agar plates supplemented with GDCA, whereas they grew well on MRS agar plates without supplementation, demonstrating that DSM 32846 does not express BSH activity.
Clinical safety and tolerability
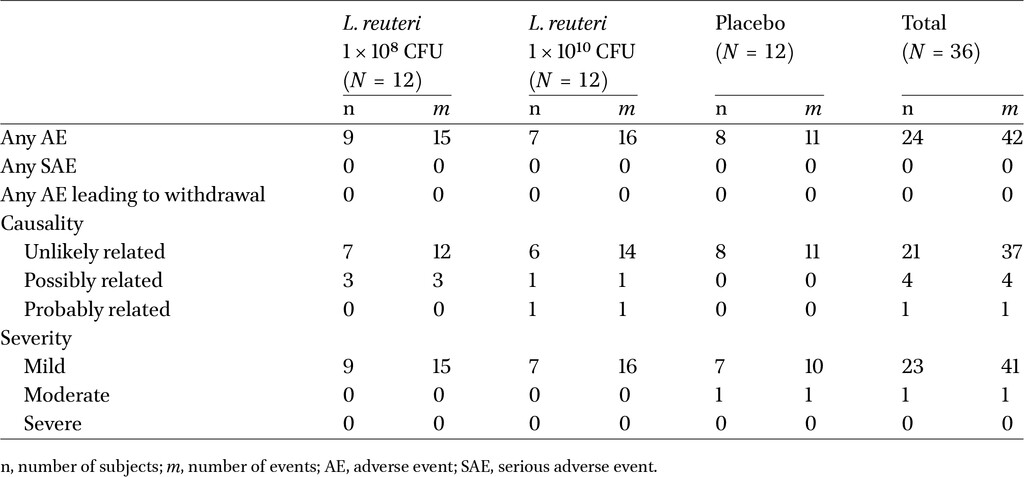
The safety and tolerability of L. reuteri DSM 32846 was evaluated in a clinical study. 35 subjects completed the study, of whom one subject was lost to follow-up. Gender, ethnicity, age and body mass index were equally distributed across the three study arms. Of the 35 subjects who completed the study, 27 had 100% compliance, meaning that the study product was consumed according to the instructions every day. Five of the 27 subjects took one dose extra. Six subjects missed one dose each, one subject missed two doses, and one subject missed one dose and then took two doses too many.
There were no clinically significant changes from baseline to Day 28 in the primary endpoints vital signs and physical examination. There were no deaths, serious adverse events, or other significant adverse events (AE), or withdrawals due to AE during the study. Most reported AEs were mild and unlikely to be related to the study product (Table 2). Constipation, flatulence, gastroesophageal reflux disease and dyspepsia were reported as single events by four separate subjects, and these AEs were assessed as possibly related. Flatulence was reported by two subjects in the high-dose arm and was assessed as mild in intensity. The only GI AE assessed as likely related to the use of the study product was one of the two instances of flatulence. Apart from the slight occurrence of flatulence in the DSM 32846 high-dose arm, there was no trend with regards to AE type between the high-dose, low-dose and placebo arms of the study.
There were no clinically significant changes in clinical chemistry or haematology parameters from baseline to Day 28, overall or for any of the study products used. There were no individual values assessed as abnormal. Subjective tolerability, as measured by GSRS, indicated low levels of GI discomfort across the three study arms. The GSRS scores did not show any obvious differences between the study arms at any timepoint, and DSM 32846 did not increase any of the symptom clusters or total GSRS scores compared to placebo. The presence of viable DSM 32846 in stools was measured at baseline and Day 28. At baseline, none of the participants had detectable levels of DSM 32846 in faeces, and the same result was obtained for the Day 28 samples from participants receiving placebo. At Day 28, viable DSM 32846 was detected in 73% (8 out of 11) of the samples from both the high- and low-dose groups.
In conclusion, L. reuteri DSM 32846 at a dose of up to 1010 CFU/day was safe and well tolerated in a healthy male and female population as assessed by AEs, vital signs, clinical laboratory parameters, physical examinations, and subjective GI tolerability assessment during the study.
4 Discussion
This paper describes the development, characterisation and demonstration of safety and tolerability of the novel strain Limosilactobacillus reuteri DSM 32846. Like L. reuteri DSM 17938, DSM 32846 was shown to be safe from a genomic, phenotypic and clinical perspective, which adds a safety perspective to the already existing preclinical data for DSM 32846.
Common strategies to increase efficacy of probiotics are to provide the bacteria with a substrate, e.g. a prebiotic fibre that in combination with the probiotics can be the components of a so-called synbiotic (Swanson et al., 2020), or by combining probiotic strains that together provide synergistic effects (Dias et al., 2022). Microbial strain development is another strategy for improving probiotics (Wang et al., 2023) and a suitable target for the selection may be a trait that is known or thought to contribute directly to the bioactivity of the strain. Selecting strains with improved tolerance to a stressor is a relatively straightforward task, but selecting strains with increased bioactivity is more challenging. For example, L. reuteri DSM 17938 is known to have calming effects on a stressed intestine, normalising intestinal motility (Burgos et al., 2015), but selecting for an improvement in this function is complicated. Instead, we hypothesised that a strain with increased robustness under stressful conditions relevant for the GI-tract may result in higher metabolic activity and increased likelihood of beneficial interactions with host cells. In this work, we have applied microbial strain development to modulate the properties of L. reuteri DSM 17938, and DSM 32846 was obtained after exposure of the parental strain to bile. DSM 32846 was found to be more tolerant to bile exposure, suggesting an adaptation to conditions in the small intestine. However, the strain did not show increased growth rate in a bile-rich substrate. Probiotic bacteria rarely colonise the GI-tract but are considered transient colonisers, and bile contributes to this microbial equilibrium and to the colonisation resistance of the GI tract (Larabi et al., 2023). In line with this, it is reasonable to believe that DSM 32846 overcomes the passage of the bile-rich upper small intestine better than the parental strain and thereby obtains an increased metabolic activity in the ileum. Interestingly, previous studies have shown that the strain has altered properties that extend beyond increased bile tolerance (Liu et al., 2023; Pang et al., 2022). This is in line with what has been described for various forms of adaptive mutagenesis in Escherichia coli (Foster, 2007), which describes adaptive mutation as ‘The appearance among cells under selection of mutations that relieve the selective pressure whether or not other nonselected mutations are also produced’. The development of DSM 32846 with its bile-tolerant phenotype is indeed a result of stress, but the underlying mechanisms of its properties remain unknown and are a topic for future studies. The results of this study, together with the previous preclinical studies showing that DSM 32846 and DSM 17938 exhibit different bioactivities (Liu et al., 2023; Pang et al., 2022), indicate that the bile-tolerant phenotype has come with other properties. Clinical studies addressing the probiotic effects of DSM 32846 are warranted.
Another interesting difference between DSM 32846 and DSM 17938 is the increased secretion of 5′-nucleotidase (5′NT) in DSM 32846. The role of the enzyme is to convert AMP to adenosine, a potent signalling molecule involved in the regulation of many host functions such as gastrointestinal motility, intestinal epithelial barrier function and immune homeostasis (Antonioli et al., 2008; Haskó and Cronstein, 2004; Liu and Xia, 2015; Stepanova and Aherne, 2024). The increased production has been previously described (Liu et al., 2023; Pang et al., 2022) and to this we now add that the increased secreted activity of DSM 32846 is also retained during industrial-scale production of freeze-dried bacterial culture powder. It has been shown that extracellular membrane vesicles (MV) of DSM 32846 express higher 5′NT activity than MV from DSM 17938 (Pang et al., 2022). Furthermore, He and colleagues have shown that DSM 17938 rescues scurfy mice from multiorgan inflammation and that adenosine receptor 2A is required for this effect (He et al., 2017). Later, the same group reported increased adenosine and inosine in the cecum of DSM 32846-administered scurfy mice, an observation not seen with DSM 17938, confirming that DSM 32846 produces adenosine and inosine in vivo (Liu et al., 2023). Showing that the strain retains the increased adenosine production capacity in culture powders that can be used for the production of probiotic products strengthens the notion that the strain may exhibit potentiated bioactivity.
Limosilactobacillus reuteri was already 2007 evaluated by the European Food Safety Authority (EFSA) and has since been listed as a Qualified Presumption of Safety (QPS) species. In addition, there is a need to evaluate the safety of new probiotic strains at three different levels: genotypic, phenotypic and clinical. The genome safety assessment showed that there are no safety concerns with DSM 32846. However, the phenotypic assessment of antibiotic resistance showed that the MIC values for ampicillin, tetracycline and chloramphenicol all exceeded the breakpoints defined by EFSA. The resistance to ampicillin can be explained by point mutations in genes encoding penicillin-binding proteins, similar to what was previously described for DSM 17938 (Rosander et al., 2008). Both DSM 32846 and DSM 17938 lack the tetracycline resistance gene tet(W), which is found in their common ancestor L. reuteri ATCC 55730 (Rosander et al., 2008). DSM 17938 was shown in the same study to have an MIC value for tetracycline below the cut-off value. In that study, the Etest method was used, but when using the methods defined in later guidelines (broth microdilution), both DSM 17938 and DSM 32846 got MIC’s above the cut-off values. By analysing the genome sequences, point mutations of rpsJ were identified that result in an exchange of amino acid residues in the ribosomal protein S10. Both Hu et al. (2005) and Lupien et al. (2015) have described corresponding point mutations in the homologous rpsJ in Neisseria and Streptococcus pneumoniae, respectively, resulting in increased tolerance to tetracycline. Most likely, the substitution of amino acid residues in S10 leads to a decreased affinity of tetracycline to its target, the 30S subunit of the ribosome, which increases tolerance to the antibiotics. The rpsJ gene as well as the genes encoding the penicillin binding proteins are all housekeeping genes located on the chromosomes of DSM 32846 and DSM 17938 and furthermore have no adjacent mobile genetic element. This type of genes is considered to pose a low risk of transmission (Devirgiliis et al., 2011; van Reenen and Dicks, 2011) and is therefore considered as generally acceptable by EFSA (EFSA FEEDAP Panel, 2012). Regarding the elevated MIC for chloramphenicol, our study suggests that it can be explained by an incorrectly defined cut-off value. EFSA should only use data generated with a microdilution method but for L. reuteri and chloramphenicol they have based the cut-off value on Etest results (EFSA FEEDAP Panel, 2012; Egervärn et al., 2007) (https://mic.eucast.org/search/, accessed 4 March 2025).
In the clinical safety study, we demonstrated that L. reuteri DSM 32846 is safe and well tolerated in healthy adults. In addition, no significant changes were found in any of the study groups high-dose DSM 32846, low-dose DSM 32846, or placebo in physical examination, vital signs, biochemical markers, AE or GSRS scores. Subjective tolerability as measured by GSRS indicated low levels of GI discomfort. In addition, viable DSM 32846 was detected in faeces from 73% of subjects receiving the strain, which is consistent with a previous study with L. reuteri DSM 17938 (Rosander et al., 2008). In summary, no safety concerns were identified and there was no trend between the three study arms with regards to the types of AEs reported, AE reporting frequency, intensity or relatedness to the study products. Additionally, subjective evaluation of GI tolerability, as evaluated by GSRS, showed no clear differences between the study arms, and the use of active study product did not result in increased GI discomfort or less tolerance compared to placebo.
5 Conclusions
Limosilactobacillus reuteri DSM 32846, a novel strain derived from L. reuteri DSM 17938, exhibits increased bile tolerance and increased secretion of 5′-nucleotidase involved in the production of the signalling molecule adenosine. Genomic, phenotypic and clinical safety assessments show that the strain is safe for human consumption at a concentration up to 1010 CFU. Altogether, DSM 32846 is an interesting strain that differs from its parental strain and is interesting for further evaluation in clinical studies.
Corresponding author; e-mail: stefan.roos@slu.se
Acknowledgements
Ludwig Ermann Lundberg and Malin Sendelius have contributed equally to this work and share first authorship. We would like to thank Jonas Faijerson Säljö, Pernilla Garberg and Helena Bysell for their support during this work; Anna Pielach at the Center for Cellular Imaging (University of Gothenburg) for aiding with SEM imaging. We also acknowledge Lena Brive (RISE Research Institutes of Sweden AB) for performing analyses of biogenic amines and D,L-lactate. We further thank Roger Karlsson and Anders Karlsson (Nanoxis Consulting AB), and Beatriz Piñeiro Iglesias (Department of Infectious Diseases, Sahlgrenska Academy, University of Gothenburg) for fruitful discussions and preparations before the SEM imaging, and Erik Rein-Hedin (CTC Clinical Trial Consultants AB) for supervising and aiding with the clinical study.
Authors’ contribution
Conceptualisation, L.E.L., MS and S.R.; Methodology, L.E.L., M.S., AP., P.L., PPM, and S.R.; Software, L.E.L., P.L.; Validation, C.L., S.R.; Formal analysis, L.E.L., M.S., A.P., P.L. and P.P.M.; Investigation, L.E.L., M.S., A.P., S.R. and P.P.M.; Resources, G.G. and S.R.; Writing – original draft, L.E.L., M.S., C.L., and S.R.; Writing – review & editing, L.E.L., M.S., C.L., G.G., and S.R.; Visualisation, L.E.L., M.S., C.L. and S.R.; Supervision, S.R.; Project administration, L.E.L, M.S. and S.R.; Funding acquisition, G.G. and S.R. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
L.E.L., M.S., C.L., G.G. and S.R. are employees of BioGaia AB.
Funding
This work was supported by financial support by BioGaia AB, Stockholm, Sweden.
References
Ali, M.S., Lee, E.-B., Lim, S.-K., Suk, K. and Park, S.-C., 2023. Isolation and identification of Limosilactobacillus reuteri PSC102 and evaluation of its potential probiotic, antioxidant, and antibacterial properties. Antioxidants 12(2): 238.
Antonioli, L., Fornai, M., Colucci, R., Ghisu, N., Tuccori, M., Del Tacca, M. and Blandizzi, C., 2008. Regulation of enteric functions by adenosine: pathophysiological and pharmacological implications. Pharmacology & Therapeutics 120(3): 233-253.
Binda, S., Hill, C., Johansen, E., Obis, D., Pot, B., Sanders, M.E., Tremblay, A. and Ouwehand, A.C., 2020. Criteria to qualify microorganisms as ‘probiotic’ in foods and dietary supplements. Frontiers in Microbiology 11: 1662.
British Standards Institute Staff, 2010. ISO 10932. Milk and Milk Products. Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB).
Burgos, A.P., Wang, L., Neufeld, K.A.M., Mao, Y.K., Ahmadzai, M., Janssen, L.J., Stanisz, A.M., Bienenstock, J. and Kunze, W.A., 2015. The TRPV1 channel in rodents is a major target for antinociceptive effect of the probiotic Lactobacillus reuteri DSM 17938. Journal of Physiology 593(17): 3943-3957.
Chung, T.C., Axelsson, L., Lindgren, S.E. and Dobrogosz, W.J., 1989. In vitro studies on reuterin synthesis by Lactobacillus reuteri. Microbial Ecology in Health and Disease 2(2): 137-144.
Connolly, E., Abrahamsson, T. and Björkstén, B., 2005. Safety of D(-)-lactic acid producing bacteria in the human infant. Journal of Pediatric Gastroenterology and Nutrition 41(4): 489-492.
Devirgiliis, C., Barile, S. and Perozzi, G., 2011. Antibiotic resistance determinants in the interplay between food and gut microbiota. Genes & Nutrition 6(3): 275-284.
Dias, J.A.R., Alves, L.L., Barros, F.A.L., Cordeiro, C.A.M., Meneses, J.O., Santos, T.B.R., Santos, C.C.M., Paixão, P.E.G., Filho, R.M.N., Martins, M.L., Pereira, S.A., Mouriño, J.L.P., Diniz, L.E.C., Maria, A.N., Carneiro, P.C.F. and Fujimoto, R.Y., 2022. Comparative effects of using a single strain probiotic and multi-strain probiotic on the productive performance and disease resistance in Oreochromis niloticus. Aquaculture 550: 737855.
Dos Reis Buzzo Zermiani, A.P., de Paula Soares, A.L.P.P., da Silva Guedes de Moura, B.L., Miguel, E.R.A., Lopes, L.D.G., de Carvalho Scharf Santana, N., da Silva Santos, T., Demarchi, I.G. and Teixeira, J.J., 2021. Evidence of Lactobacillus reuteri to reduce colic in breastfed babies: systematic review and meta-analysis. Complementary Therapies in Medicine 63: 102781.
Dunne, C., O’Mahony, L., Murphy, L., Thornton, G., Morrissey, D., O’Halloran, S., Feeney, M., Flynn, S., Fitzgerald, G., Daly, C., Kiely, B., O’Sullivan, G.C., Shanahan, F. and Collins, J.K., 2001. In vitro selection criteria for probiotic bacteria of human origin: correlation with in vivo findings. The American Journal of Clinical Nutrition 73(2 Suppl): 386S-392S.
EFSA FEEDAP Panel, 2012. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA Journal 10(6): 559.
EFSA FEEDAP Panel, 2018. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA Journal 16(3): e05206.
Egervärn, M., Danielsen, M., Roos, S., Lindmark, H. and Lindgren, S., 2007. Antibiotic susceptibility profiles of Lactobacillus reuteri and Lactobacillus fermentum. Journal of Food Protection 70(2): 412-418.
Ermann Lundberg, L., Pallabi Mishra, P., Liu, P., Forsberg, M.M., Sverremark-Ekström, E., Grompone, G., Håkansson, S., Linninge, C. and Roos, S., 2024. Bifidobacterium longum subsp. longum BG-L47 boosts growth and activity of Limosilactobacillus reuteri DSM 17938 and its extracellular membrane vesicles. Applied and Environmental Microbiology 90(7): e0024724.
Fatheree, N.Y., Liu, Y., Taylor, C.M., Hoang, T.K., Cai, C., Rahbar, M.H., Hessabi, M., Ferris, M., McMurtry, V., Wong, C., Vu, T., Dancsak, T., Wang, T., Gleason, W., Bandla, V., Navarro, F., Tran, D.Q. and Rhoads, J.M., 2017. Lactobacillus reuteri for infants with colic: a double-blind, placebo-controlled, randomized clinical trial. The Journal of Pediatrics 191: 170-178.e172.
Foster, P.L., 2007. Stress-induced mutagenesis in bacteria. Critical Reviews in Biochemistry and Molecular Biology 42(5): 373-397.
Galhardo, R.S., Hastings, P.J. and Rosenberg, S.M., 2007. Mutation as a stress response and the regulation of evolvability. Critical Reviews in Biochemistry and Molecular Biology 42(5): 399-435.
Gutiérrez-Castrellón, P., Indrio, F., Bolio-Galvis, A., Jiménez-Gutiérrez, C., Jimenez-Escobar, I. and López-Velázquez, G., 2017. Efficacy of Lactobacillus reuteri DSM 17938 for infantile colic: systematic review with network meta-analysis. Medicine 96(51): e9375.
Hammes, W.P. and Hertel, C., 2006. The genera Lactobacillus and Carnobacterium. In: Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H. and Stackebrandt, E. (eds.) The prokaryotes: volume 4: bacteria: firmicutes, cyanobacteria. Springer US, New York, NY, pp. 320-403.
Haskó, G. and Cronstein, B.N., 2004. Adenosine: an endogenous regulator of innate immunity. Trends in Immunology 25(1): 33-39.
He, B., Hoang, T.K., Tran, D.Q., Rhoads, J.M. and Liu, Y., 2017. Adenosine A2A receptor deletion blocks the beneficial effects of Lactobacillus reuteri in regulatory T-deficient scurfy mice. Frontiers in Immunology 8: 1680.
Hernández-Gómez, J.G., López-Bonilla, A., Trejo-Tapia, G., Ávila-Reyes, S.V., Jiménez-Aparicio, A.R. and Hernández-Sánchez, H., 2021. In vitro Bile Salt Hydrolase (BSH) activity screening of different probiotic microorganisms. Foods 10(3): 674.
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology & Hepatology 11(8): 506-514. https://doi.org/10.1038/nrgastro.2014.66
Hu, M., Nandi, S., Davies, C. and Nicholas, R.A., 2005. High-level chromosomally mediated tetracycline resistance in Neisseria gonorrhoeae results from a point mutation in the rpsJ gene encoding ribosomal protein S10 in combination with the mtrR and penB resistance determinants. Antimicrobial Agents and Chemotherapy 49(10): 4327-4334.
Humblot, C., Murkovic, M., Rigottier-Gois, L., Bensaada, M., Bouclet, A., Andrieux, C., Anba, J. and Rabot, S., 2007. β-Glucuronidase in human intestinal microbiota is necessary for the colonic genotoxicity of the food-borne carcinogen 2-amino-3-methylimidazo[4,5-f] quinoline in rats. Carcinogenesis 28(11): 2419-2425.
Karimi, S., Jonsson, H., Lundh, T. and Roos, S., 2018. Lactobacillus reuteri strains protect epithelial barrier integrity of IPEC-J2 monolayers from the detrimental effect of enterotoxigenic Escherichia coli. Physiological Reports 6(2): e13514.
Kim, D.H. and Jin, Y.H., 2001. Intestinal bacterial beta-glucuronidase activity of patients with colon cancer. Archives of Pharmacal Research 24(6): 564-567.
Klare, I., Konstabel, C., Müller-Bertling, S., Reissbrodt, R., Huys, G., Vancanneyt, M., Swings, J., Goossens, H. and Witte, W., 2005. Evaluation of new broth media for microdilution antibiotic susceptibility testing of Lactobacilli, Pediococci, Lactococci, and Bifidobacteria. Applied and Environmental Microbiology 71(12): 8982-8986.
Ko, H.I., Jeong, C.H., Hong, S.W., Eun, J.-B. and Kim, T.-W., 2022. Optimizing conditions in the acid tolerance test for potential probiotics using response surface methodology. Microbiology Spectrum 10(4): e0162522.
Larabi, A.B., Masson, H.L.P. and Bäumler, A.J., 2023. Bile acids as modulators of gut microbiota composition and function. Gut Microbes 15(1): 2172671.
Lee, B.S., Ban, O.-H., Bang, W.Y., Chae, S.A., Oh, S., Park, C., Lee, M., Kim, S.-J., Yang, J. and Jung, Y.H., 2021. Safety assessment of Lactobacillus reuteri IDCC 3701 based on phenotypic and genomic analysis. Annals of Microbiology 71(1): 1-6.
Li, F., Cheng, C.C., Zheng, J., Liu, J., Quevedo, R.M., Li, J., Roos, S., Gänzle, M.G. and Walter, J., 2021. Limosilactobacillus balticus sp. nov., Limosilactobacillus agrestis sp. nov., Limosilactobacillus albertensis sp. nov., Limosilactobacillus rudii sp. nov. and Limosilactobacillus fastidiosus sp. nov., five novel Limosilactobacillus species isolated from the vertebrate gastrointestinal tract, and proposal of six subspecies of Limosilactobacillus reuteri adapted to the gastrointestinal tract of specific vertebrate hosts. International Journal of Systematic and Evolutionary Microbiology 71(2): 004644.
Liu, H. and Xia, Y., 2015. Beneficial and detrimental role of adenosine signaling in diseases and therapy. Journal of Applied Physiology 119(10): 1173-1182.
Liu, Y., Armbrister, S.A., Okeugo, B., Mills, T.W., Daniel, R.C., Oh, J.-H., Van Pijkeren, J.-P., Park, E.S., Saleh, Z.M., Lahiri, S., Roos, S. and Rhoads, J., 2023. Probiotic-derived ecto-5’-nucleotidase produces anti-inflammatory adenosine metabolites in treg-deficient scurfy mice. Probiotics and Antimicrobial Proteins 15(4): 1-13.
Lupien, A., Gingras, H., Leprohon, P. and Ouellette, M., 2015. Induced tigecycline resistance in Streptococcus pneumoniae mutants reveals mutations in ribosomal proteins and rRNA. Journal of Antimicrobial Chemotherapy 70(11): 2973-2980.
Marmur, J., 1961. A procedure for the isolation of deoxyribonucleic acid from micro-organisms. Journal of Molecular Biology 3(2): 208-218. IN1.
Mu, Q., Tavella, V.J. and Luo, X.M., 2018. Role of Lactobacillus reuteri in human health and diseases. Frontiers in Microbiology 9: 25.
Pang, Y., Ermann Lundberg, L., Mata Forsberg, M., Ahl, D., Bysell, H., Pallin, A., Sverremark-Ekström, E., Karlsson, R., Jonsson, H. and Roos, S., 2022. Extracellular membrane vesicles from Limosilactobacillus reuteri strengthen the intestinal epithelial integrity, modulate cytokine responses and antagonize activation of TRPV1. Frontiers in Microbiology 13: 1032202.
Papagaroufalis, K., Fotiou, A., Egli, D., Tran, L.-A. and Steenhout, P., 2014. A randomized double blind controlled safety trial evaluating d-lactic acid production in healthy infants fed a Lactobacillus reuteri-containing formula. Nutrition and Metabolic Insights 7: 19-27.
Petersen, C., 2005. D-lactic acidosis. Nutrition in Clinical Practice 20(6): 634-645.
Richter, M., Rosselló-Móra, R., Oliver Glöckner, F. and Peplies, J., 2016. JSpeciesWS: a web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 32(6): 929-931.
Romani Vestman, N., Hasslöf, P., Keller, M.K., Granström, E., Roos, S., Twetman, S. and Stecksén-Blicks, C., 2013. Lactobacillus reuteri influences regrowth of mutans streptococci after full-mouth disinfection: a double-blind, randomised controlled trial. Caries Research 47(4): 338-345.
Roos, S., Karner, F., Axelsson, L. and Jonsson, H., 2000. Lactobacillus mucosae sp. nov., a new species with in vitro mucus-binding activity isolated from pig intestine. International Journal of Systematic and Evolutionary Microbiology 50(1): 251-258.
Rosander, A., Connolly, E. and Roos, S., 2008. Removal of antibiotic resistance gene-carrying plasmids from Lactobacillus reuteri ATCC 55730 and characterization of the resulting daughter strain, L. reuteri DSM 17938. Applied and Environmental Microbiology 74(19): 6032-6040.
Rosengren, A., Fabricius, A., Guss, B., Sylvén, S. and Lindqvist, R., 2010. Occurrence of foodborne pathogens and characterization of Staphylococcus aureus in cheese produced on farm-dairies. International Journal of Food Microbiology 144(2): 263-269.
Ruiz-Capillas, C. and Herranz, A.H., 2019. Biogenic amines on food safety. MDPI. https://doi.org/10.3390/books978-3-03921-055-8
Scheijen, J.L.J.M., Hanssen, N.M.J., van de Waarenburg, M.P.H., Jonkers, D.M.A.E., Stehouwer, C.D.A. and Schalkwijk, C.G., 2012. L(+) and D(−) lactate are increased in plasma and urine samples of type 2 diabetes as measured by a simultaneous quantification of L(+) and D(−) lactate by reversed-phase liquid chromatography tandem mass spectrometry. Journal of Diabetes Research 2012(1): 234812. 10. https://doi.org/10.1155/2012/234812
Sendelius, M., Axelsson, J., Liu, P. and Roos, S., 2023. Genomic, phenotypic, and clinical safety of Limosilactobacillus reuteri ATCC PTA 4659. Journal of Industrial Microbiology & Biotechnology 50(1): kuad041.
Stepanova, M. and Aherne, C.M., 2024. Adenosine in intestinal epithelial barrier function. Cells 13(5): 381.
Sun, X., Kong, J., Zhu, S. and Liu, C., 2023. A systematic review and meta-analysis: the therapeutic and preventive effect of Lactobacillus reuteri DSM 17,938 addition in children with diarrhea. BMC Gastroenterology 23(1): 1-11.
Sung, V., D’Amico, F., Cabana, M.D., Chau, K., Koren, G., Savino, F., Szajewska, H., Deshpande, G., Dupont, C., Indrio, F., Mentula, S., Partty, A. and Tancredi, D., 2017. Lactobacillus reuteri to treat infant colic: a meta-analysis. Pediatrics 141(1): e20171811.
Svedlund, J., Sjödin, I. and Dotevall, G., 1988. GSRS – a clinical rating scale for gastrointestinal symptoms in patients with irritable bowel syndrome and peptic ulcer disease. Digestive Diseases and Sciences 33(2): 129-134.
Swanson, K.S., Gibson, G.R., Hutkins, R., Reimer, R.A., Reid, G., Verbeke, K., Scott, K.P., Holscher, H.D., Azad, M.B., Delzenne, N.M. and Sanders, M.E., 2020. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nature Reviews Gastroenterology & Hepatology 17: 687-701. https://doi.org/10.1038/s41575-020-0344-2
Szajewska, H., Urbańska, M., Chmielewska, A., Weizman, Z. and Shamir, R., 2014. Meta-analysis: Lactobacillus reuteri strain DSM 17938 (and the original strain ATCC 55730) for treating acute gastroenteritis in children. Beneficial Microbes 5(3): 285-293.
Urbańska, M. and Szajewska, H., 2014. The efficacy of Lactobacillus reuteri DSM 17938 in infants and children: a review of the current evidence. European Journal of Pediatrics 173(10): 1327-1337.
van Reenen, C.A. and Dicks, L.M.T., 2011. Horizontal gene transfer amongst probiotic lactic acid bacteria and other intestinal microbiota: what are the possibilities? A review. Archives of Microbiology 193(3): 157-168.
Wall, T., Båth, K., Britton, R.A., Jonsson, H., Versalovic, J. and Roos, S., 2007. The early response to acid shock in Lactobacillus reuteri involves the ClpL chaperone and a putative cell wall-altering esterase. Applied and Environmental Microbiology 73(12): 3924-3935.
Wang, G., Li, Q., Zhang, Z., Yin, X., Wang, B. and Yang, X., 2023. Recent progress in adaptive laboratory evolution of industrial microorganisms. Journal of Industrial Microbiology & Biotechnology 50(1): kuac023.
Wolf, B.W., Garleb, K.A., Ataya, D.G. and Casas, I.A., 1995. Safety and tolerance of Lactobacillus reuteri in healthy adult male subjects. Microbial Ecology in Health and Disease 8(2): 41-50.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}