Abstract
Metabolic disorders are a major global health problem. Gut microbiota not only affect host metabolism through metabolites, inflammatory processes, and microbial-derived extracellular vesicles, but they also modulate the host microRNA, which may impact the host metabolism. Hence, the underlying mechanisms between gut microbiota-microRNA interaction can potentially be a novel alternative strategy for treating metabolic disorders. This review aims to give an update on the latest evidence and current knowledge of the underlying mechanisms of gut microbiota-miRNA interaction, focusing on metabolic homeostasis. Gut microbiota mainly communicate with host microRNA through lipopolysaccharide and secondary microbial metabolites. These signalling messengers circulate around the metabolic organs and modify gene expression through microRNA interference. Interestingly, while intestinal microRNAs play a vital role in both intestinal barrier and gut microbiota homeostasis, the presence of gut microbiota is also required for the proper functioning of intestinal microRNAs, suggesting a cooperative mechanism in intestinal health. Although the correlations between gut microbiota and microRNA have been observed in both mice and humans, a causal relationship should be confirmed. Moreover, further investigation is needed to provide more evidence of a gut microbiota-microRNA interaction to support the possibility of using that axis as a novel therapeutic target to treat metabolic disorders.

Citation: Beneficial Microbes 15, 1 (2024) ; 10.1163/18762891-20230103
1 Introduction
Metabolic disorders, diseases caused by metabolic imbalance, including type 2 diabetes (T2DM), dyslipidaemia (DLP), hypertension, metabolic-associated fatty liver disease, and obesity, are currently a global health problem. The prevalence of these diseases have been increasing in the past two decades (Hirode and Wong, 2020; Kang et al., 2020), even in the young adult (Al-Shehri et al., 2021) and paediatric populations (Song et al., 2022). Moreover, these metabolic disorders increase the risk of ischemic heart disease, which has been a worldwide leading cause of death since 2000 (Nowbar et al., 2019). Therefore, the primary and secondary prevention of metabolic disorders is essential to reduce the number of ischemic heart disease cases.
Gut microbiota, the microorganisms colonising the gastrointestinal tract, have been reported to be associated with metabolic disorders. Loss of gastrointestinal microbial equilibrium, also known as gut dysbiosis, has been found in people with T2DM (Wu et al., 2020) and metabolic syndrome (MetS) (León Aguilera et al., 2022), which is a group of risk factors for T2DM and cardiovascular disease (CVD). As a result, the potential role of gut microbiota as both therapeutics and targets for metabolic disorder prevention has been proposed (León Aguilera et al., 2022; Mutalub et al., 2022). Lipopolysaccharide (LPS), a gram-negative microbial component, and microbial metabolites, e.g. short-chain fatty acids (SCFA), indole-3-propionic acid (IPA), and secondary bile acids, are the primary signalling messengers in the communication between gut microbiota and the host (Salazar et al., 2020; Sikalidis and Maykish, 2020). Apart from these mechanisms, epigenetic modulations by gut microbiota that influence the host’s metabolism have received a lot of attention in recent years.
MicroRNA (miRNA), which are short, non-coding, single-stranded RNA, act as epigenetic modulators and various miRNAs have been extensively investigated for their regulatory role in lipid and glucose metabolism in the past decade (Du et al., 2021a). After being synthesised, the interaction between miRNA and a messenger RNA (mRNA) target leads to either mRNA translational repression or degradation depending on the miRNA-mRNA complementarity. Each miRNA may have one or more mRNA targets. Various miRNAs and their metabolism-related target genes have been validated, suggesting that miRNA may be a promising therapeutic option for metabolic disorders (Desgagné et al., 2017; Du et al., 2021a).
The association between miRNA and gut microbiota has been widely studied recently. Gut microbiota can modulate the host miRNA and be regulated by either host miRNA or diet-derived miRNA (Bi et al., 2020). Although the conclusive mechanism of miRNA and gut microbiota interaction has not yet been clarified, several pathways have been proposed (Behrouzi et al., 2020; Wu et al., 2021). These underlying mechanisms can potentially be a novel or alternative strategy for preventing and treating metabolic disorders. Therefore, this review aims to update the latest evidence for gut microbiota-miRNA interaction in metabolic disorders and the current knowledge on the underlying mechanisms focusing on metabolic homeostasis.
2 Overview of miRNA
The biogenesis of miRNA
The first miRNA was identified in 1993 (Lee et al., 1993), and it was reported to be 22 nucleotides long on average. A primary miRNA (pri-miRNA) is generated with a 5′ cap and a 3′ poly (A) tail by either RNA polymerase II or RNA polymerase III in the nucleus. The structure of the hairpin loop on pri-miRNA is recognised and cleaved by DiGeorge Syndrome Critical Region 8 (DGCR8) and Drosha, a ribonuclease (RNase) III family protein, respectively, forming a precursor miRNA (pre-miRNA). The pre-miRNA is transported to the cytoplasm by exportin-5 and the Ras-related nuclear protein guanosine triphosphate (Matsuyama and Suzuki, 2019; O’Brien et al., 2018).
In the cytoplasm, dicer, an RNase III family member, together with the trans-activation response RNA binding protein removes the terminal loop of the pre-miRNA hairpin, generating a mature miRNA duplex. The mature miRNA strand initiated from the 5′ end of hairpin loop is labelled with the suffix −5p, while the strand with −3p indicates the duplex of the −5p strand. Although both mature strands are loaded into the Argonaute (Ago) proteins afterward, only the guide strand is preserved. The passenger strand (miRNA*) is removed from the Ago proteins and degraded (Matsuyama and Suzuki, 2019; O’Brien et al., 2018). The guide strand forms the RNA-induced silencing complex (RISC) with Ago proteins. RISC interacts with the mRNA target through the seed sequence, the 2-8 nucleotides at the 5′ end of miRNA, and the supplementary sequence, the 13-16 nucleotides at the 5′ end (Marzec, 2020). While a partial complementarity between RISC and mRNA target suppresses mRNA translation, the target mRNA is degraded upon perfect complementarity with RISC (Matsuyama and Suzuki, 2019; O’Brien et al., 2018).
miRNA and host communication
In 2008, Lawrie et al. first reported the presence of miRNA in serum and plasma samples (Lawrie et al., 2008). The levels of circulating miRNAs in humans with cancer showed no difference from the cancer xenograft-bearing mice and remained constant after exposure to plasma RNase (Mitchell et al., 2008), food intake, and sampling at different times (Kupec et al., 2022; Sabbaghian et al., 2022). These findings brought attention to the idea that miRNA may not only be responsible for an autocrine signalling, but also may be a mediator of cell-to-cell communication (Mori et al., 2019).
Apart from the communication between human cells, miRNAs present in plants and animal-sourced foods can be absorbed into the circulatory system and interfere with mammalian genes (Baier et al., 2014; Zhang et al., 2012). In addition, food-derived exosomes containing miRNAs can be taken up by gut microbiota and interact with bacterial mRNAs, supporting the cross-domain communicating capacity of miRNAs (Teng et al., 2018). Hence, several review articles in the last few years have proposed a new role for miRNA in gut microbiota-host communication (Bi et al., 2020; Du et al., 2021b; Li et al., 2020c). This communication between the intestinal microbiome and host cells may explain why and how the alterations to microbial composition influence the host’s health.
3 Current evidence of molecular mechanisms underlying gut microbiota-miRNA interaction in host metabolic homeostasis
The association between gut microbiota and host miRNA was first published in 2011. Dalmasso et al. (2011) observed a difference in miRNA expression between germ-free mice and colonised mice. After this discovery, the trend of global research on gut microbiota-miRNA interaction substantially increased from 1 to 24% of this study field over the span of 10 years (Yan et al., 2022). Briefly, the expression of hepatic miRNAs in animal studies was found to be significantly changed after introducing a HFD (Blasco-Baque et al., 2017; Dong et al., 2021; Jia et al., 2018). The altered patterns of these miRNAs were consistent with the shifting of gut microbiota composition. Interestingly, many of the predicted target genes of these miRNA were involved in metabolic pathways, confirming the role of miRNAs in modulating metabolism. In line with the animal studies, correlations between gut microbiota and miRNA were reported in patients with T2DM, obesity, and MetS (Assmann et al., 2020; Li et al., 2020b; Wortelboer et al., 2022). Although correlations between gut microbiota and miRNA were observed in MetS, the causal relationship still needs to be confirmed to support the possibility of using gut microbiota-miRNA interaction as a novel therapeutic target to treat metabolic disorders. Hence, understanding the underlying mechanisms of this interconnection is necessary.
Immunity and inflammation
McKenna et al. (2010) reported the presence of an association between miRNA and the intestinal barrier in 2010. Loose and fatty stools were observed in mice with genetic ablation of Dicer1, a gene-encoded miRNA processing protein. Correspondingly, both intestinal permeability and inflammatory cells were more prominent in Dicer1-deficient mice compared to non-mutant group. Although the importance of miRNA processing for intestinal homeostasis was demonstrated in this study, the expression of intestinal miR-21 was inconsistent with inflammatory bowel disease (IBD) patients (Yan et al., 2020). Higher expression of intestinal miR-21 was found in both dextran sodium sulphate (DSS) colitis model mice and patients with IBD (Johnston et al., 2018; Yan et al., 2020), while a downregulated miR-21 was observed in Dicer1-deficient mice (McKenna et al., 2010). The benefits of a deficiency of miR-21 were confirmed in DSS-induced colitis model mice lacking miR-21, showing less bloody stool and lower inflammation and cell damage compared to the wild-type group (Johnston et al., 2018). Since the Dicer1 gene has a crucial role in non-specific miRNA maturation (Foulkes et al., 2014), there are various factors affecting intestinal homeostasis. Interestingly, the benefits of a deficiency of miR-21 in DSS-induced colitis model mice decreased after administration of antibiotics and there was a difference in gut microbiota composition observed between miR-21-knockout mice and the control group, implying a role of gut microbiota in the function of miR-21 (Johnston et al., 2018). This finding supported the previous study that showed that expression of different miRNAs depended on the presence of gut microbiota (Singh et al., 2012). Collectively, these results indicated that intestinal miRNA expression and gut microbiota cooperate in the regulation of intestinal inflammation.
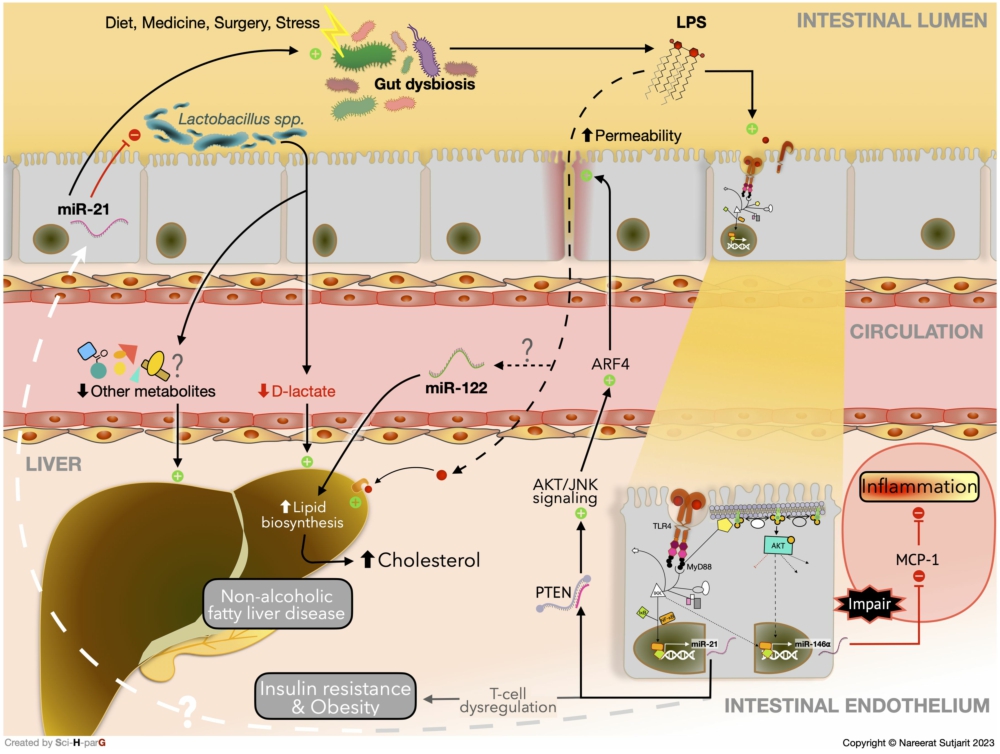
In a comparable result to the study by Johnston et al. (2018), miR-21-deficient mice also showed an anti-inflammatory effect on intestinal inflammation in the bile duct ligation (BDL) surgery model (Santos et al., 2020). This model has been recognised for its complications, including elevated plasma LPS, liver injury, gut dysbiosis, and metabolic disturbance (Zhou et al., 2023). Although the mRNA expression of intestinal tight-junction proteins and junctional adhesion molecules were significantly decreased after BDL, in miR-21-knockout mice, the mRNA levels were maintained after BDL. In addition, the lack of miR-21 alleviated the shift in gut microbiota composition after BDL-induced liver injury and facilitated the growth of probiotic Lactobacillus. Lower serum endotoxin levels were reported in miR-21-deficient mice, corresponding with a stronger intestinal barrier and gut microbiota homeostasis. The severity of hepatic injury and fibrosis was also reduced in miR-21-knockout mice (Santos et al., 2020). Likewise, the improvement of metabolic disturbance after providing an antagonist of miR-21 was confirmed in streptozotocin-induced T2DM rat (Wang et al., 2020). The role of miR-21 on lipid metabolism and chronic liver diseases, e.g. non-alcoholic fatty liver disease (NAFLD) or liver fibrosis, has been well-documented (Zhang et al., 2020). Therefore, these findings suggested that miR-21 deficiency has a hepatic protective effect through modulating gut microbiota, regulating the tight junctions of intestinal epithelial cells (IECs), and reducing endotoxemia (Santos et al., 2020). Nonetheless, the crosstalk between gut microbiota and miR-21 on lipid metabolism should be more explored.
Similar to the previous study (Singh et al., 2012), the expression of miR-21-5p in germ-free (GF) mice was markedly different from conventional mice. LPS enhanced the expression of miR-21-5p in human IEC lines and the LPS-induced miR-21-5p was suppressed with anti-TLR4 antibody treatment (Nakata et al., 2017). This outcome supported the study of Frederick et al., suggesting that LPS stimulates miR-21 expression through TLR4/MyD88/NF-κB (Sheedy et al., 2010). The mechanism of miR-21-5p in increasing IEC permeability was further investigated. It was revealed that miR-21-5p targets PTEN, leading to the activation of the protein kinase B (Akt) and c-Jun N-terminal kinase (JNK) signalling pathway, which resulted in the augmentation of ADP ribosylation factor 4 (ARF4), a member of the family of small GTPases involved in IEC homeostasis. IEC permeability was then found to be increased. In contrast, ARF4 inhibition promoted the expression of tight junction proteins (Nakata et al., 2017). In line with the above evidences, downregulated PTEN with overexpression of miR-21 was observed in the intestinal tissue of Crohn’s disease patients (Wang et al., 2022), which has been reported to be associated with MetS (Dragasevic et al., 2020). Moreover, in a mouse model of colitis, the decreased ratio of regulatory T cells and T helper-1 cells, which plays a vital role in obesity with insulin resistance, was reversed by treatment with antagomiR-21, a synthesised antagonist of miR-21 (Liu et al., 2021; Wang et al., 2022). As a result, chronic inflammation in the intestine and gut dysbiosis-related LPS may aggravate the progression of metabolic disorders through miR-21 resulting in impaired permeability of IECs and induction of a systemic inflammatory response.
Apart from the importance of miRNA in impairing the intestinal barrier, miRNA also modulates the inflammation caused by LPS from gut dysbiosis (Anzola et al., 2018). The benefits of miR-146a on anti-inflammation and metabolic disorders have been reported in several studies (Li et al., 2020a; Runtsch et al., 2019). The level of IEC miR-146a was substantially elevated with the highest response to LPS stimulation. Upon the binding of LPS with TLR4, MyD88, a primary downstream adapter of TLR, is triggered and subsequently activates the NF-κB and Akt pathways. Since the inhibition in each step of the TLR4/MyD88/NF-κB-Akt pathway resulted in downregulation of both pre- and mature miR-146a, the underlying mechanism of miR-146a overexpression induced by LPS is likely to be this pathway. In addition, the overexpression of miR-146a eventually repressed monocyte chemoattractant protein-1 (MCP-1) secretion, which is a chemokine that promotes inflammatory cell infiltration. These outcomes confirm that LPS stimulates miR-146a expression through the TLR4/MyD88/NF-κB-Akt pathway, and the miR-146a subsequently regulates inflammatory processes. Interestingly, upregulated expression of miR-146a was found in acute colitis-induced mice, but not in a chronic inflammation-induced model. These results imply that the degree of intestinal barrier damage, which is more severe in the acute model, also plays a role in the appropriate response of miRNA suppression of inflammation. On the other hand, the regulatory function of miR-146a may be impaired in the setting of chronic inflammation (Anzola et al., 2018). The dysfunction of anti-inflammatory miRNA may be one of the causes of chronic inflammatory diseases and, thus, has the potential to be a novel approach to disease prevention and therapy. Besides, since LPS stimulated the anti-inflammatory miR-146a expression, the importance of gut homeostasis should be highlighted.
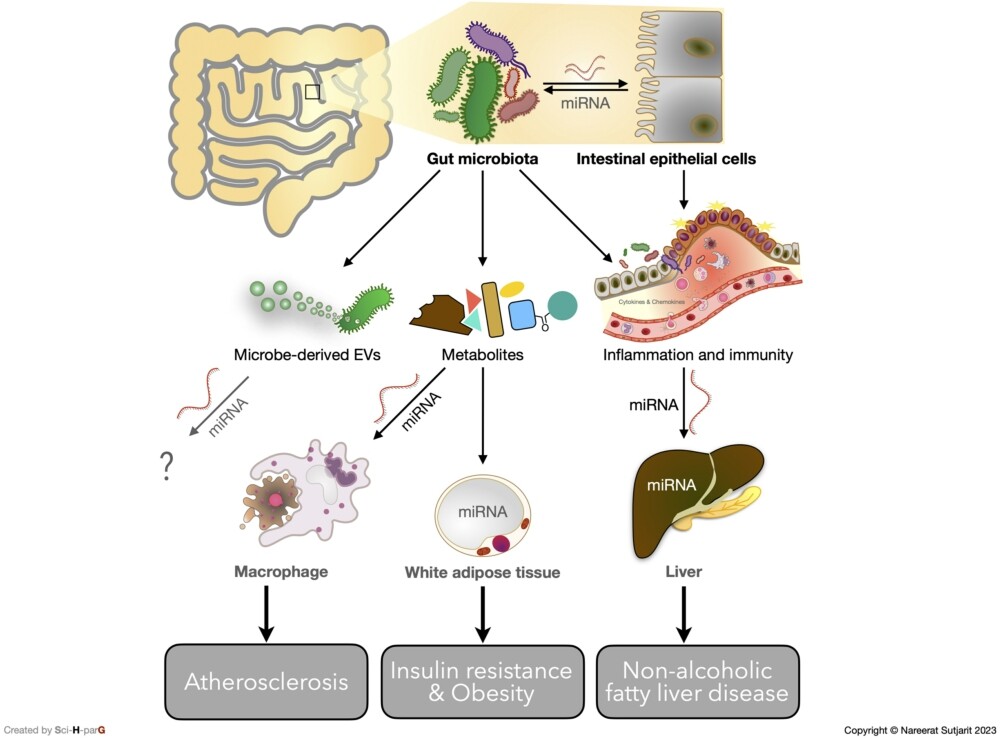
The link between gut microbiota composition, impaired intestinal barrier, circulating miRNA, and hepatic inflammation in the development of NAFLD was observed in Sprague Dawley rats. The bacterial phyla Tenericutes and Cyanobacteria were observed to be more abundant in the control group, while Bacteroidetes and Proteobacteria, which are both gram-negative bacteria containing LPS, were more prominent in the NAFLD group. Accordingly, genes involved with tight junction proteins, including occludin (Ocln) and F11 receptor (F11r), were markedly downregulated in the NAFLD-induced rats. Furthermore, the expression of hepatic TLR4 and MyD88 were significantly increased in the NAFLD group, which was consistent with the increased expression of tumour necrosis factor (TNF)-α gene and hepatic concentration. These outcomes demonstrate a potential role for gut dysbiosis-derived LPS in hepatic inflammation, which was also correlated with NAFLD progression. Although relationships between NAFLD and circulating miR-122 have been observed, there was no mention of a link to inflammation in this study (Jia et al., 2018). Comparable with the results from the NAFLD rat model, higher serum and plasma miR-122 were found in human subjects with MetS and T2DM (Willeit et al., 2017). This evidence proposed a promising role of miR-122 as a predictor of MetS and T2DM development. Willeit et al. (2017) also confirmed the effect of miR-122 in an inhibited miR-122 mice model using antagomiR-122. After treatment with antagomiR-122, the expression of genes involved in lipid biosynthesis, including ATP citrate lyase (Acly) and sterol regulatory element-binding protein 1 (Srebp1), and triglyceride transportation, including microsomal triglyceride transfer protein (Mttp), were all declined. As expected, a substantially reduced total cholesterol level was observed in mice that had been treated with antagomiR-122 (Willeit et al., 2017). Taken together, the results suggest that there may be a connection between gut dysbiosis-derived systemic LPS, IEC impairment, and miR-122 upregulation that ultimately promotes lipid metabolism disturbance and NAFLD progression. The potential interplay between host miRNAs and gut microbiota on metabolic disorders via inflammatory response and gut microbiota modulation is summarised in Figure 1.
The potential interplay between host miRNAs and gut microbiota on metabolic disorders via inflammatory response and gut microbiota modulation.
Citation: Beneficial Microbes 15, 1 (2024) ; 10.1163/18762891-20230103
Blasco-Baque et al. also reported evidence of LPS specifically stimulating hepatic miRNA expression in primary hepatocytes of Cd14 knockout mice. LPS was found to induce miR-181a, miR-666, and miR-21 expression. However, only miR-666 and miR-21 were statistically negatively correlated with hepatic triglyceride concentration. In this model, while Firmicutes were positively associated with liver triglyceride level, Bacteroides acidifaciens showed the strongest negative relationship with hepatic triglyceride concentration. In accordance with these observed correlations, both miR-666 and miR-21 had an inverse association with Firmicutes and a positive relationship with B. acidifaciens. These findings not only suggested a link between gut microbiota, miRNA, and hepatic triglycerides, but also emphasised the potential impact of LPS on gut dysbiosis-hepatic miRNA communication (Blasco-Baque et al., 2017). In addition, miR-21 in a macrophage-like cell line was also found to be elevated with LPS incubation in a dose- and time-dependent manner. The upregulated expression of miR-21 in macrophages inhibited both fat accumulation and oxidised-LDL (ox-LDL) uptake in the macrophages. As a result of the overexpression of miR-21 induced by LPS, foam cell formation, a hallmark of atherosclerosis progression, was inhibited. Furthermore, the augmentation of miR-21 suppressed the TLR4/NF-κB signalling pathway in macrophage, leading to a reduction in the production of the pro-inflammatory cytokine interleukin (IL)-6, and an increase in production of the anti-inflammatory cytokine IL-10. The beneficial effects of miR-21 on preventing inflammation were also supported by the results of anti-miR-21 inhibitor transfection in another experiment (Feng et al., 2014). Hence, miR-21 may play a role in atherosclerosis progression, which is a primary consequence of metabolic disorders related to chronic inflammation (Hasheminasabgorji and Jha, 2021). Nonetheless, the positive effects of miR-21 were contrary to the aforementioned evidences (Nakata et al., 2017; Santos et al., 2020; Wang et al., 2020; Wang et al., 2022). The functions of miRNA in each type of cell may be various. Prior to clinical trials, a further investigation for a more in-depth mechanism is required.
Microbial metabolites
The alteration of gut microbiota is not only related to inflammation, but it also influences microbial metabolite shifting, resulting in metabolic disturbance. The abundance of SCFA producers, Prevotella and Bifidobacterium, were reported to be higher in miR-21-deficient mice (Johnston et al., 2018). Since the depletion of these genera was also detected in T2DM patients (Cunningham et al., 2021), it is possible that miR-21 may influence host metabolism via gut microbiota modulation altering microbial metabolites in these patients. Nevertheless, the causal relationship between miR-21 and these beneficial microbes should be confirmed in vitro study. Interestingly, synthetic human miR-21 inhibited the growth of probiotic Lactobacillus spp. in an in vitro study, and this probiotic was also found to be prominent in miR-21-knockout mice (Santos et al., 2020). This probiotic has been extensively reported on for its positive effect in liver diseases, including NAFLD and liver fibrosis (Ritze et al., 2014; Santos et al., 2020). Unsurprisingly, an improvement in hepatic inflammation was observed in miR-21-deficient mice (Santos et al., 2020). D-lactate, the main product of Lactobacillus spp. metabolised by D-lactate dehydrogenase (D-LDH), can either be a fuel for cellular metabolism or a substrate for other SCFAs production (Ewaschuk et al., 2005; Santos et al., 2020). Although serum D-lactate was not detected, the hepatic D-LDH mRNA was significantly upregulated in mice lacking miR-21. Santos et al., thereby, suggested that the suppression of Lactobacillus spp. growth and subsequent reduction of D-lactate caused by miR-21 aggravate liver injury (Santos et al., 2020). Although, the potential benefits of D-lactate on liver fibrosis was mentioned in this study, several recent studies observed the positive correlations between D-lactate on chronic liver diseases and impaired intestinal barrier (Peng et al., 2018; Zhang et al., 2023). The favourable effects of miR-21 deficiency promoting Lactobacillus spp. growth may due to other microbial metabolites, e.g. acetate, propionate, and butyrate (Markowiak-Kopeć and Śliżewska, 2020) (Figure 1).
Butyrate, one of the most well-known microbial metabolite products, is absorbed by enterocytes and subsequently binds to GPR43 on multiple target organs, modulating host metabolism (Coppola et al., 2021). Although Pant et al. reported a beneficial role of butyrate in liver cancer through sirtuin 1 (SIRT1) inhibition, which was found to be mediated by upregulation of miR-22 expression (Pant et al., 2017), another study showed that the overexpression of miR-22 promoted hepatic steatosis via inhibition of fibroblast growth factor 21 (FGF21) (Hu et al., 2020). Furthermore, butyrate-mediated suppression of histone deacetylase 3 (HDAC3) resulting in increased expression of FGF21, a peptide hormone regulating glucose and lipid metabolism, has also been reported (Li et al., 2012). These studies denote the action of butyrate on different epigenetic modulations under distinct circumstances. Interestingly, a recent study reported evidence of butyrate interfering with a post-transcriptional regulator of miRNA expression (Das et al., 2022). After the adenylate and uridylate (AU)-rich element binding factor 1 (AUF1) interacts with AU-rich elements (AREs) on the 3′-UTR of target mRNA, the mRNA becomes unstable, resulting in diminished mRNA translation and even mRNA degradation. Dicer1 mRNA segments can be targets for AUF1. Since Dicer1 mRNA encodes the dicer enzyme, which is critical for miRNA maturation, lower overall miRNA levels were reported after AUF1 overexpression (Abdelmohsen et al., 2012). Das et al. observed that butyrate has a role in AUF1 upregulation, leading to Dicer1 downregulation and ultimately repressing hepatic miR-122 expression. Lipid biogenesis-related genes, such as HMG CoA reductase, 7-dehydrocholesterol reductase, and HMG CoA synthase 1, have been reported to be targets of miR-122; hence, lower cellular and serum cholesterol levels were observed in Huh7 cells and in mice fed with a HFD, respectively (Das et al., 2022).
IPA, a tryptophan-derived microbial metabolite, has been well-recognised as an inflammation suppressor, intestinal barrier protector, and host metabolic modulator (Sehgal et al., 2022). A negative correlation between IPA level and CVD risk has been reported, and it was also found that the gut microbiota producing IPA, especially Clostridium and Peptostreptococcus, were reduced in patients with coronary artery disease compared to healthy individuals (Xue et al., 2022). The depletion of these bacterial genera also reduced the serum IPA level by more than 50% in mice fed a Western diet. Treatment with IPA attenuated fat accumulation in both human and mouse macrophages in a dose-dependent manner by promoting cholesterol efflux to ApoA1. ATP-binding cassette transporter A1 (ABCA1), the primary lipid transporter in macrophages, was observed to be the main channel of cholesterol efflux induced by IPA. Interestingly, the expression of transcriptional factor SPI1 was reduced after IPA treatment. Under normal circumstances, this transcriptional factor upregulates miR-142-5p expression, which interacts with ABCA1. Taken together, it is thought that IPA treatment enhanced macrophage cholesterol efflux by inhibiting SPI1, downregulating miR-142-5p, and ultimately promoting ABCA1 expression. The alleviation of atherosclerotic plaques was confirmed in an animal study, corresponding with the above mechanism (Xue et al., 2022). Moreover, another tryptophan-derived metabolite, indole, was found to specifically impact miR-181 expression in WAT without LPS involvement (Virtue et al., 2019). It was also found that miR-181-knockout in macrophages and hepatocytes had no impact on weight gain and insulin resistance. Likewise, after receiving a HFD, increased expression of miR-181 was only detected in adipocytes of obese mice, thereby implying that miR-181 specifically acts on adipocytes in obesity and insulin resistance. Additionally, downregulation of miR-181 in WAT was revealed only in the presence of Escherichia coli strains carrying the enzyme tryptophanase (TnaA), which catalyses the conversion of tryptophan into indole. Correspondingly, mice fed with a HFD and colonised with TnaA-knockout E. coli strains had aggravated weight gain and impaired glucose tolerance compared with the control group (Virtue et al., 2019). These results denote a link between gut microbiota-derived metabolites, host miRNA, and metabolic homeostasis.
Not all microbial metabolites yield benefits for host metabolism. Trimethylamine N-oxide (TMAO), a primary metabolite derived from animal products, has been recognised as a pro-CVD factor (Canyelles et al., 2023). TMAO was found to promote the expression of miR-17-92 cluster in various cell lines (Dı́ez-Ricote et al., 2022). Serpin Family E Member 1 (SERPINE1), one of the mRNA targets of miR-17-92 cluster, has been reported to have a positive correlation with atherogenesis (Kubota et al., 2021). Although the expression of SERPINE1 and IL-12A were upregulated with TMAO treatment, in the same way as miR-17-92 cluster, there was no further investigation to prove the direct influence of miR-17-92 cluster on SERPINE1 and IL-12A expression (Dı́ez-Ricote et al., 2022). Since animal products are a primary source of high biological value proteins, which are necessary for human development (Giromini and Givens, 2022), further study should be done to confirm the role of TMAO on miRNA, expanding the prevention perspective on TMAO-related metabolic diseases.
Apart from the above well-known microbial metabolites, compounds produced by fermentation of phytochemicals by gut microbiota have also been shown to be involved in the regulation of metabolic homeostasis and inflammation. Urolithin A, a bioactive compound from ellagitannins and ellagic acid, significantly decreased miR-10, miR-99b, miR-146a, and miR-155 expression after LPS stimulation and inhibited NF-kB activation (Abdelazeem et al., 2021). The repression of IκBα phosphorylation, mitogen activated protein kinase (MAPK), and Akt/mechanistic target of rapamycin (mTOR) signalling pathways were found to be the underlying mechanisms of urolithin alleviating inflammation (Abdelazeem et al., 2021). Since miR-146a is strongly associated with LPS-induced gut inflammation through the TLR4/MyD88/NF-κB-Akt pathway as mentioned above (Anzola et al., 2018), the anti-inflammatory role of urolithin A via miRNA regulation was indicated as it suppressed miR-146a expression (Abdelazeem et al., 2021). In addition to urolithin A, another bioactive compound derived from anthocyanins and metabolised by gut microbiota, protocatechuic acid, has an impact on metabolism and atherogenesis by enhancing macrophage cholesterol efflux (Wang et al., 2012) similar to the study with IPA mentioned above from Xue et al. (2022). However, in the case of protocatechuic acid, the active miRNA was distinct. Protocatechuic acid reduced miR-10b expression and directly targeted ABCA1 and ATP Binding Cassette Subfamily G Member 1. The upregulation of these genes enhanced cholesterol efflux from macrophages and thus, alleviated foam cell formation (Wang et al., 2012).
Microbial-derived extracellular vesicles
Similar to human extracellular vesicles (EVs), microbial-derived extracellular vesicles (MEVs) contain genetic information, e.g. miRNA or mRNA, and carry their genetic cargo to the target cell and subsequently initiate communication by either ligand-receptor interaction, specific surface antigen-receptor interaction, or membrane fusion (Martellucci et al., 2020; Nik Mohamed Kamal and Shahidan, 2020). Although the research into the disruption of metabolic homeostasis induced by miRNA in gut MEVs is scarce, there are a few reports of the effects of oral microbiota-derived EVs on host health (Choi et al., 2017; Han et al., 2019). Like IBD, chronic periodontitis can interfere with host health via oral microbial dysbiosis, barrier impairment, and chronic systemic inflammation (Byrd and Gulati, 2021). The release of miRNA-size small RNA (msRNA) from outer-membrane vesicles (OMVs) derived from immuno-interfering periodontal pathogens, including Aggregatibacter actinomycetemcomitans (Aa), Porphyromonas gingivalis, and Treponema denticola, has been observed. As expected, several predicted gene targets of these msRNAs are involved with the immune system. Moreover, the expression of IL-5, IL-13, and IL-15 was substantially decreased after synthetic msRNA transfection of T-cells (Choi et al., 2017). Research by Han et al. (2019) also found that the presence of OMVs derived from Aa significantly increased TNF-α secretion from macrophages, while the presence of lysed OMVs showed no significant change. This study suggests the importance of OMVs on bacterial genetic protection and the presence of an inter-kingdom communication channel by direct uptake to the macrophage. After entering the macrophage, the msRNA of Aa triggered TLR8 and activated the NF-κB signalling pathway and the expression of TNF-α protein was subsequently elevated. In addition, the msRNA was able to interact with host Ago2 and form RISC. Therefore, the msRNA from periodontal pathogens may be able to impact host gene expression through interaction with Ago2 and RISC formation. OMVs from Aa were also found to be able to cross the blood-brain barrier and were able to eventually promote TNF-α in the brain (Han et al., 2019).
Even though a large number of studies have been carried out in recent years on the impacts of MEVs on host metabolism, the majority of research has focused on the presence of MEVs, LPS, and microbial DNA (Castaño et al., 2023). Consistent with the above mechanisms, MEVs derived from beneficial gut microbiota, such as Akkermansia muciniphila, Lactobacillus spp., and Propionibacterium freudenreichii, ameliorated intestinal inflammation (Choi et al., 2020; Rodovalho et al., 2020), regulated IEC tight junctions (Chelakkot et al., 2018), and, ultimately, deferred the development of obesity in mice fed with a HFD (Ashrafian et al., 2021). Interestingly, although the abundance of Pseudomonas panacis and Pseudomonas cedrina were similar between mice fed a HFD and the regular diet group, the MEVs derived from these two pathogens were significantly different. MEVs released from P. panacis, which were more prominent in HFD-fed mice, contained more LPS than MEVs from P. cedrina. Furthermore, the P. panacis-derived MEVs promoted insulin resistance by inhibiting Akt signalling pathway in adipocytes and suppressed GLUT-4 translocation in skeletal muscle (Choi et al., 2015). This result corresponded with the study by Nah et al., as there was a correlation between faecal, blood, and urine MEVs in patients with T2DM (Nah et al., 2019). Collectively, these findings support the importance of MEVs derived from gut microbiota in host metabolic homeostasis.
4 Conclusions
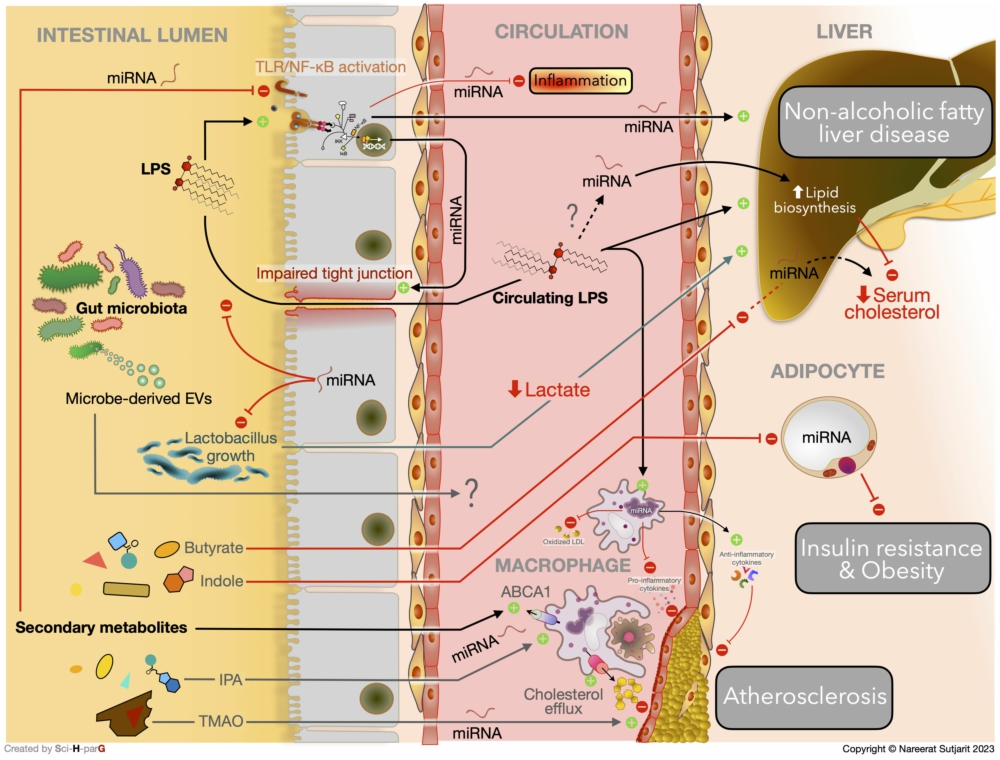
According to the current evidence, gut microbiota not only influence the host metabolism through the immune system, microbial metabolite, gut hormone, or nervous system (Cook and Mansuy-Aubert, 2022), they can also modulate host metabolic homeostasis via post-transcriptional regulation of a gene in target organs. The signalling messengers of gut microbiota, including LPS, MEVs, and gut microbiota-derived metabolites, play a key role in this epigenetic control (Figure 2). The discovery of gut microbiota specifically interfering with host miRNA in each metabolic organ may provide a novel solution for managing metabolic disorders. Nevertheless, further investigation is needed to provide more evidence for this novel therapeutic strategy.
Current evidence of molecular mechanisms underlying gut microbiota-miRNA interaction in host metabolic homeostasis.
Citation: Beneficial Microbes 15, 1 (2024) ; 10.1163/18762891-20230103
Corresponding author; e-mail: nareerat.sut@mahidol.ac.th
Authors’ contribution
Conceptualisation: PP and NS; writing-original draft preparation: PP; writing-review and editing: PP, NS, and JS. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
The authors declare no conflict of interest.
Funding
This research received no external funding.
References
Abdelazeem, K.N.M., Kalo, M.Z., Beer-Hammer, S. and Lang, F., 2021. The gut microbiota metabolite urolithin A inhibits NF-κB activation in LPS stimulated BMDMs. Scientific Reports 11: 7117. https://doi.org/10.1038/s41598-021-86514-6
Abdelmohsen, K., Tominaga-Yamanaka, K., Srikantan, S., Yoon, J.H., Kang, M.J. and Gorospe, M., 2012. RNA-binding protein AUF1 represses Dicer expression. Nucleic Acids Research 40: 11531-11544. https://doi.org/10.1093/nar/gks930
Al-Shehri, H.A., Al-Asmari, A.K., Khan, H.A., Horaib, G.B., Al-Buraidi, A., Al-Sharif, A.A., Kadasah, S.G., Al-Omani, S., Mohammed, F.S., Abbasmanthiri, R. and Osman, N.M., 2021. Recent trends of metabolic syndrome and its components in military recruits from Saudi Arabia. Medicines 8: 65.
Anzola, A., González, R., Gámez-Belmonte, R., Ocón, B., Aranda, C.J., Martı́nez-Moya, P., López-Posadas, R., Hernández-Chirlaque, C., Sánchez de Medina, F. and Martı́nez-Augustin, O., 2018. miR-146a regulates the crosstalk between intestinal epithelial cells, microbial components and inflammatory stimuli. Scientific Reports 8: 17350. https://doi.org/10.1038/s41598-018-35338-y
Ashrafian, F., Keshavarz Azizi Raftar, S., Lari, A., Shahryari, A., Abdollahiyan, S., Moradi, H.R., Masoumi, M., Davari, M., Khatami, S., Omrani, M.D., Vaziri, F., Masotti, A. and Siadat, S.D., 2021. Extracellular vesicles and pasteurized cells derived from Akkermansia muciniphila protect against high-fat induced obesity in mice. Microbial Cell Factories 20: 219. https://doi.org/10.1186/s12934-021-01709-w
Assmann, T.S., Cuevas-Sierra, A., Riezu-Boj, J.I., Milagro, F.I. and Martı́nez, J.A., 2020. Comprehensive analysis reveals novel interactions between circulating MicroRNAs and gut microbiota composition in human obesity. International Journal of Molecular Sciences 21: 9509. https://doi.org/10.3390/ijms21249509
Baier, S.R., Nguyen, C., Xie, F., Wood, J.R. and Zempleni, J., 2014. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. Journal of Nutrition 144: 1495-1500. https://doi.org/10.3945/jn.114.196436
Behrouzi, A., Ashrafian, F., Mazaheri, H., Lari, A., Nouri, M., Riazi Rad, F., Hoseini Tavassol, Z. and Siadat, S.D., 2020. The importance of interaction between MicroRNAs and gut microbiota in several pathways. Microbial Pathogenesis 144: 104200. https://doi.org/10.1016/j.micpath.2020.104200
Bi, K., Zhang, X., Chen, W. and Diao, H., 2020. MicroRNAs regulate intestinal immunity and gut microbiota for gastrointestinal health: a comprehensive review. Genes 11: 1057. https://doi.org/10.3390/genes11091075
Blasco-Baque, V., Coupé, B., Fabre, A., Handgraaf, S., Gourdy, P., Arnal, J.F., Courtney, M., Schuster-Klein, C., Guardiola, B., Tercé, F., Burcelin, R. and Serino, M., 2017. Associations between hepatic miRNA expression, liver triacylglycerols and gut microbiota during metabolic adaptation to high-fat diet in mice. Diabetologia 60: 690-700. https://doi.org/10.1007/s00125-017-4209-3
Byrd, K.M. and Gulati, A.S., 2021. The ‘gum-gut’ axis in inflammatory bowel diseases: a hypothesis-driven review of associations and advances. Frontiers in Immunology 12: 620124. https://doi.org/10.3389/fimmu.2021.620124
Canyelles, M., Borràs, C., Rotllan, N., Tondo, M., Escolà-Gil, J.C. and Blanco-Vaca, F., 2023. Gut microbiota-derived TMAO: a causal factor promoting atherosclerotic cardiovascular disease? International Journal of Molecular Sciences 24: 1940.
Castaño, C., Novials, A. and Párrizas, M., 2023. An overview of inter-tissue and inter-kingdom communication mediated by extracellular vesicles in the regulation of mammalian metabolism. International Journal of Molecular Sciences 24: 2071. https://doi.org/10.3390/ijms24032071
Chelakkot, C., Choi, Y., Kim, D.K., Park, H.T., Ghim, J., Kwon, Y., Jeon, J., Kim, M.S., Jee, Y.K., Gho, Y.S., Park, H.S., Kim, Y.K. and Ryu, S.H., 2018. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Experimental and Molecular Medicine 50: e450. https://doi.org/10.1038/emm.2017.282
Choi, J.H., Moon, C.M., Shin, T.S., Kim, E.K., McDowell, A., Jo, M.K., Joo, Y.H., Kim, S.E., Jung, H.K., Shim, K.N., Jung, S.A. and Kim, Y.K., 2020. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Experimental and Molecular Medicine 52: 423-437. https://doi.org/10.1038/s12276-019-0359-3
Choi, J.W., Kim, S.C., Hong, S.H. and Lee, H.J., 2017. Secretable small RNAs via outer membrane vesicles in periodontal pathogens. Journal of Dental Research 96: 458-466. https://doi.org/10.1177/0022034516685071
Choi, Y., Kwon, Y., Kim, D.K., Jeon, J., Jang, S.C., Wang, T., Ban, M., Kim, M.H., Jeon, S.G., Kim, M.S., Choi, C.S., Jee, Y.K., Gho, Y.S., Ryu, S.H. and Kim, Y.K., 2015. Gut microbe-derived extracellular vesicles induce insulin resistance, thereby impairing glucose metabolism in skeletal muscle. Scientific Reports 5: 15878. https://doi.org/10.1038/srep15878
Cook, T.M. and Mansuy-Aubert, V., 2022. Communication between the gut microbiota and peripheral nervous system in health and chronic disease. Gut Microbes 14: 2068365. https://doi.org/10.1080/19490976.2022.2068365
Coppola, S., Avagliano, C., Calignano, A. and Berni Canani, R., 2021. The protective role of butyrate against obesity and obesity-related diseases. Molecules 26: 682.
Cunningham, A.L., Stephens, J.W. and Harris, D.A., 2021. Gut microbiota influence in type 2 diabetes mellitus (T2DM). Gut Pathogens 13: 50. https://doi.org/10.1186/s13099-021-00446-0
Dalmasso, G., Nguyen, H.T., Yan, Y., Laroui, H., Charania, M.A., Ayyadurai, S., Sitaraman, S.V. and Merlin, D., 2011. Microbiota modulate host gene expression via microRNAs. PLoS ONE 6: e19293. https://doi.org/10.1371/journal.pone.0019293
Das, O., Kundu, J., Ghosh, A., Gautam, A., Ghosh, S., Chakraborty, M., Masid, A., Gauri, S.S., Mitra, D., Dutta, M., Mukherjee, B., Sinha, S. and Bhaumik, M., 2022. AUF-1 knockdown in mice undermines gut microbial butyrate-driven hypocholesterolemia through AUF-1-Dicer-1-mir-122 hierarchy. Frontiers in Cellular and Infection Microbiology 12: 1011386. https://doi.org/10.3389/fcimb.2022.1011386
Desgagné, V., Bouchard, L. and Guérin, R., 2017. MicroRNAs in lipoprotein and lipid metabolism: from biological function to clinical application. Clinical Chemistry and Laboratory Medicine 55: 667-686. https://doi.org/10.1515/cclm-2016-0575
Dı́ez-Ricote, L., Ruiz-Valderrey, P., Micó, V., Blanco, R., Tomé-Carneiro, J., Dávalos, A., Ordovás, J.M. and Daimiel, L., 2022. TMAO upregulates members of the miR-17/92 cluster and impacts targets associated with atherosclerosis. International Journal of Molecular Sciences 23: 2107. https://doi.org/10.3390/ijms232012107
Dong, J., Gu, W., Yang, X., Zeng, L., Wang, X., Mu, J., Wang, Y., Li, F., Yang, M. and Yu, J., 2021. Crosstalk between Polygonatum kingianum, the miRNA, and gut microbiota in the regulation of lipid metabolism. Frontiers in Pharmacology 12: 740528. https://doi.org/10.3389/fphar.2021.740528
Dragasevic, S., Stankovic, B., Kotur, N., Sokic-Milutinovic, A., Milovanovic, T., Lukic, S., Milosavljevic, T., Srzentic Drazilov, S., Klaassen, K., Pavlovic, S. and Popovic, D., 2020. Metabolic syndrome in inflammatory bowel disease: association with genetic markers of obesity and inflammation. Metabolic Syndrome and Related Disorders 18: 31-38. https://doi.org/10.1089/met.2019.0090
Du, H., Zhao, Y., Li, H., Wang, D.W. and Chen, C., 2021a. Roles of microRNAs in glucose and lipid metabolism in the heart. Frontiers in Cardiovascular Medicine 8: 716213. https://doi.org/10.3389/fcvm.2021.716213
Du, X., Ley, R. and Buck, A.H., 2021b. MicroRNAs and extracellular vesicles in the gut: new host modulators of the microbiome? microLife 2: uqab010. https://doi.org/10.1093/femsml/uqab010
Ewaschuk, J.B., Naylor, J.M. and Zello, G.A., 2005. D-lactate in human and ruminant metabolism. Journal of Nutrition 135: 1619-1625. https://doi.org/10.1093/jn/135.7.1619
Feng, J., Li, A., Deng, J., Yang, Y., Dang, L., Ye, Y., Li, Y. and Zhang, W., 2014. miR-21 attenuates lipopolysaccharide-induced lipid accumulation and inflammatory response: potential role in cerebrovascular disease. Lipids in Health and Disease 13: 27. https://doi.org/10.1186/1476-511x-13-27
Foulkes, W.D., Priest, J.R. and Duchaine, T.F., 2014. DICER1: mutations, microRNAs and mechanisms. Nature Reviews Cancer 14: 662-672. https://doi.org/10.1038/nrc3802
Giromini, C. and Givens, D.I., 2022. Benefits and risks associated with meat consumption during key life processes and in relation to the risk of chronic diseases. Foods 11. https://doi.org/10.3390/foods11142063
Han, E.C., Choi, S.Y., Lee, Y., Park, J.W., Hong, S.H. and Lee, H.J., 2019. Extracellular RNAs in periodontopathogenic outer membrane vesicles promote TNF-α production in human macrophages and cross the blood-brain barrier in mice. FASEB Journal 33: 13412-13422. https://doi.org/10.1096/fj.201901575R
Hasheminasabgorji, E. and Jha, J.C., 2021. Dyslipidemia, diabetes and atherosclerosis: role of inflammation and ROS-redox-sensitive factors. Biomedicines 9: 1602.
Hirode, G. and Wong, R.J., 2020. Trends in the prevalence of metabolic syndrome in the United States, 2011-2016. JAMA 323: 2526-2528. https://doi.org/10.1001/jama.2020.4501
Hu, Y., Liu, H.X., Jena, P.K., Sheng, L., Ali, M.R. and Wan, Y.Y., 2020. miR-22 inhibition reduces hepatic steatosis via FGF21 and FGFR1 induction. JHEP Report 2: 100093. https://doi.org/10.1016/j.jhepr.2020.100093
Jia, N., Lin, X., Ma, S., Ge, S., Mu, S., Yang, C., Shi, S., Gao, L., Xu, J., Bo, T. and Zhao, J., 2018. Amelioration of hepatic steatosis is associated with modulation of gut microbiota and suppression of hepatic miR-34a in Gynostemma pentaphylla (Thunb.) Makino treated mice. Nutrition and Metabolism 15: 86. https://doi.org/10.1186/s12986-018-0323-6
Johnston, D.G.W., Williams, M.A., Thaiss, C.A., Cabrera-Rubio, R., Raverdeau, M., McEntee, C., Cotter, P.D., Elinav, E., O’Neill, L.A.J. and Corr, S.C., 2018. Loss of microRNA-21 influences the gut microbiota, causing reduced susceptibility in a murine model of colitis. Journal of Crohn’s and Colitis 12: 835-848. https://doi.org/10.1093/ecco-jcc/jjy038
Kang, S.Y., Kim, Y.J. and Park, H.S., 2020. Trends in the prevalence of non-alcoholic fatty liver disease and its future predictions in Korean men, 1998-2035. Journal of Clinical Medicine 9: 2626. https://doi.org/10.3390/jcm9082626
Kubota, M., Yoshida, Y., Kobayashi, E., Matsutani, T., Li, S.-Y., Zhang, B.-S., Mine, S., Machida, T., Takizawa, H., Hiwasa, T. and Iwadate, Y., 2021. Serum anti-SERPINE1 antibody as a potential biomarker of acute cerebral infarction. Scientific Reports 11: 21772. https://doi.org/10.1038/s41598-021-01176-8
Kupec, T., Bleilevens, A., Iborra, S., Najjari, L., Wittenborn, J., Maurer, J. and Stickeler, E., 2022. Stability of circulating microRNAs in serum. PLoS ONE 17: e0268958. https://doi.org/10.1371/journal.pone.0268958
Lawrie, C.H., Gal, S., Dunlop, H.M., Pushkaran, B., Liggins, A.P., Pulford, K., Banham, A.H., Pezzella, F., Boultwood, J., Wainscoat, J.S., Hatton, C.S.R. and Harris, A.L., 2008. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. British Journal of Haematology 141: 672-675. https://doi.org/10.1111/j.1365-2141.2008.07077.x
Lee, R.C., Feinbaum, R.L. and Ambros, V., 1993. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75: 843-854. https://doi.org/10.1016/0092-8674(93)90529-y
León Aguilera, X.E., Manzano, A., Pirela, D. and Bermúdez, V., 2022. Probiotics and gut microbiota in obesity: myths and realities of a new health revolution. Journal of Personalized Medicine 12: 1282.
Li, H., Gao, Z., Zhang, J., Ye, X., Xu, A., Ye, J. and Jia, W., 2012. Sodium butyrate stimulates expression of fibroblast growth factor 21 in liver by inhibition of histone deacetylase 3. Diabetes 61: 797-806. https://doi.org/10.2337/db11-0846
Li, K., Zhao, B., Wei, D., Wang, W., Cui, Y., Qian, L. and Liu, G., 2020a. miR-146a improves hepatic lipid and glucose metabolism by targeting MED1. International Journal of Molecular Medicine 45: 543-555. https://doi.org/10.3892/ijmm.2019.4443
Li, L., Li, C., Lv, M., Hu, Q., Guo, L. and Xiong, D., 2020b. Correlation between alterations of gut microbiota and miR-122-5p expression in patients with type 2 diabetes mellitus. Ann Transl Med 8: 1481. https://doi.org/10.21037/atm-20-6717
Li, M., Chen, W.D. and Wang, Y.D., 2020c. The roles of the gut microbiota-miRNA interaction in the host pathophysiology. Molecular Medicine 26: 101. https://doi.org/10.1186/s10020-020-00234-7
Liu, L., Hu, J., Wang, Y., Lei, H. and Xu, D., 2021. The role and research progress of the balance and interaction between regulatory T cells and other immune cells in obesity with insulin resistance. Adipocyte 10: 66-79. https://doi.org/10.1080/21623945.2021.1876375
Markowiak-Kopeć, P. and Śliżewska, K., 2020. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 12: 1107. https://doi.org/10.3390/nu12041107
Martellucci, S., Orefice, N.S., Angelucci, A., Luce, A., Caraglia, M. and Zappavigna, S., 2020. Extracellular vesicles: new endogenous shuttles for miRNAs in cancer diagnosis and therapy? International Journal of Molecular Sciences 21. https://doi.org/10.3390/ijms21186486
Marzec, M., 2020. New insights into the function of mammalian Argonaute2. PLoS Genetics 16: e1009058. https://doi.org/10.1371/journal.pgen.1009058
Matsuyama, H. and Suzuki, H.I., 2019. Systems and synthetic microRNA biology: from biogenesis to disease pathogenesis. International Journal of Molecular Sciences 21: 132. https://doi.org/10.3390/ijms21010132
McKenna, L.B., Schug, J., Vourekas, A., McKenna, J.B., Bramswig, N.C., Friedman, J.R. and Kaestner, K.H., 2010. MicroRNAs control intestinal epithelial differentiation, architecture, and barrier function. Gastroenterology 139: 1654-1664.e1. https://doi.org/10.1053/j.gastro.2010.07.040
Mitchell, P.S., Parkin, R.K., Kroh, E.M., Fritz, B.R., Wyman, S.K., Pogosova-Agadjanyan, E.L., Peterson, A., Noteboom, J., O’Briant, K.C., Allen, A., Lin, D.W., Urban, N., Drescher, C.W., Knudsen, B.S., Stirewalt, D.L., Gentleman, R., Vessella, R.L., Nelson, P.S., Martin, D.B. and Tewari, M., 2008. Circulating microRNAs as stable blood-based markers for cancer detection. Proceedings of the National Academy of Sciences of the USA 105: 10513-10518. https://doi.org/10.1073/pnas.0804549105
Mori, M.A., Ludwig, R.G., Garcia-Martin, R., Brandão, B.B. and Kahn, C.R., 2019. Extracellular miRNAs: from biomarkers to mediators of physiology and disease. Cell Metabolism 30: 656-673. https://doi.org/10.1016/j.cmet.2019.07.011
Mutalub, Y.B., Abdulwahab, M., Mohammed, A., Yahkub, A.M., Al-Mhanna, S.B., Yusof, W., Tang, S.P., Rasool, A.H.G. and Mokhtar, S.S., 2022. Gut microbiota modulation as a novel therapeutic strategy in cardiometabolic diseases. Foods 11: 2575.
Nah, G., Park, S.C., Kim, K., Kim, S., Park, J., Lee, S. and Won, S., 2019. Type-2 diabetics reduces spatial variation of microbiome based on extracellular vesicles from gut microbes across human body. Scientific Reports 9: 20136. https://doi.org/10.1038/s41598-019-56662-x
Nakata, K., Sugi, Y., Narabayashi, H., Kobayakawa, T., Nakanishi, Y., Tsuda, M., Hosono, A., Kaminogawa, S., Hanazawa, S. and Takahashi, K., 2017. Commensal microbiota-induced microRNA modulates intestinal epithelial permeability through the small GTPase ARF4. Journal of Biological Chemistry 292: 15426-15433. https://doi.org/10.1074/jbc.M117.788596
Nik Mohamed Kamal, N.N.S.B. and Shahidan, W.N.S., 2020. Non-exosomal and exosomal circulatory micrornas: which are more valid as biomarkers? Frontiers in Pharmacology 10: 1500. https://doi.org/10.3389/fphar.2019.01500
Nowbar, A.N., Gitto, M., Howard, J.P., Francis, D.P. and Al-Lamee, R., 2019. Mortality from ischemic heart disease. Circulation: Cardiovascular Quality and Outcomes 12: e005375. https://doi.org/10.1161/CIRCOUTCOMES.118.005375
O’Brien, J., Hayder, H., Zayed, Y. and Peng, C., 2018. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Frontiers in Endocrinology 9: 402. https://doi.org/10.3389/fendo.2018.00402
Pant, K., Yadav, A.K., Gupta, P., Islam, R., Saraya, A. and Venugopal, S.K., 2017. Butyrate induces ROS-mediated apoptosis by modulating miR-22/SIRT-1 pathway in hepatic cancer cells. Redox Biology 12: 340-349. https://doi.org/10.1016/j.redox.2017.03.006
Peng, J.H., Leng, J., Tian, H.J., Yang, T., Fang, Y., Feng, Q., Zhao, Y. and Hu, Y.Y., 2018. Geniposide and chlorogenic acid combination ameliorates non-alcoholic steatohepatitis involving the protection on the gut barrier function in mouse induced by high-fat diet. Frontiers in Pharmacology 9: 1399. https://doi.org/10.3389/fphar.2018.01399
Ritze, Y., Bárdos, G., Claus, A., Ehrmann, V., Bergheim, I., Schwiertz, A. and Bischoff, S.C., 2014. Lactobacillus rhamnosus GG protects against non-alcoholic fatty liver disease in mice. PLoS ONE 9: e80169. https://doi.org/10.1371/journal.pone.0080169
Rodovalho, V.R., da Luz, B.S.R., Rabah, H., do Carmo, F.L.R., Folador, E.L., Nicolas, A., Jardin, J., Briard-Bion, V., Blottière, H., Lapaque, N., Jan, G., Le Loir, Y., de Carvalho Azevedo, V.A. and Guédon, E., 2020. Extracellular vesicles produced by the probiotic Propionibacterium freudenreichii CIRM-BIA 129 mitigate inflammation by modulating the NF-κB pathway. Frontiers in Microbiology 11: 1544. https://doi.org/10.3389/fmicb.2020.01544
Runtsch, M.C., Nelson, M.C., Lee, S.H., Voth, W., Alexander, M., Hu, R., Wallace, J., Petersen, C., Panic, V., Villanueva, C.J., Evason, K.J., Bauer, K.M., Mosbruger, T., Boudina, S., Bronner, M., Round, J.L., Drummond, M.J. and O’Connell, R.M., 2019. Anti-inflammatory microRNA-146a protects mice from diet-induced metabolic disease. PLoS Genetics 15: e1007970. https://doi.org/10.1371/journal.pgen.1007970
Sabbaghian, A., Mussack, V., Kirchner, B., Bui, M.L.U., Kalani, M.R., Pfaffl, M.W. and Golalipour, M., 2022. A panel of blood-derived miRNAs with a stable expression pattern as a potential pan-cancer detection signature. Frontiers in Molecular Bioscience 9: 1030749. https://doi.org/10.3389/fmolb.2022.1030749
Salazar, J., Angarita, L., Morillo, V., Navarro, C., Martı́nez, M.S., Chacı́n, M., Torres, W., Rajotia, A., Rojas, M., Cano, C., Añez, R., Rojas, J. and Bermudez, V., 2020. Microbiota and diabetes mellitus: role of lipid mediators. Nutrients 12: 3039.
Santos, A.A., Afonso, M.B., Ramiro, R.S., Pires, D., Pimentel, M., Castro, R.E. and Rodrigues, C.M.P., 2020. Host miRNA-21 promotes liver dysfunction by targeting small intestinal Lactobacillus in mice. Gut Microbes 12: 1840766. https://doi.org/10.1080/19490976.2020.1840766
Sehgal, R., de Mello, V.D., Männistö, V., Lindström, J., Tuomilehto, J., Pihlajamäki, J. and Uusitupa, M., 2022. Indolepropionic acid, a gut bacteria-produced tryptophan metabolite and the risk of type 2 diabetes and non-alcoholic fatty liver disease. Nutrients 14: 4695.
Sheedy, F.J., Palsson-McDermott, E., Hennessy, E.J., Martin, C., O’Leary, J.J., Ruan, Q., Johnson, D.S., Chen, Y. and O’Neill, L.A.J., 2010. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nature Immunology 11: 141-147. https://doi.org/10.1038/ni.1828
Sikalidis, A.K. and Maykish, A., 2020. The gut microbiome and type 2 diabetes mellitus: discussing a complex relationship. Biomedicines 8: 8.
Singh, N., Shirdel, E.A., Waldron, L., Zhang, R.H., Jurisica, I. and Comelli, E.M., 2012. The murine caecal microRNA signature depends on the presence of the endogenous microbiota. International Journal of Biological Sciences 8: 171-186. https://doi.org/10.7150/ijbs.8.171
Song, K., Park, G., Lee, H.S., Lee, M., Lee, H.I., Ahn, J., Lee, E., Choi, H.S., Suh, J., Kwon, A., Kim, H.-S. and Chae, H.W., 2022. Trends in prediabetes and non-alcoholic fatty liver disease associated with abdominal obesity among Korean children and adolescents: based on the Korea National Health and Nutrition Examination Survey between 2009 and 2018. Biomedicines 10: 584.
Teng, Y., Ren, Y., Sayed, M., Hu, X., Lei, C., Kumar, A., Hutchins, E., Mu, J., Deng, Z., Luo, C., Sundaram, K., Sriwastva, M.K., Zhang, L., Hsieh, M., Reiman, R., Haribabu, B., Yan, J., Jala, V.R., Miller, D.M., Van Keuren-Jensen, K., Merchant, M.L., McClain, C.J., Park, J.W., Egilmez, N.K. and Zhang, H.G., 2018. Plant-derived exosomal microRNAs shape the gut microbiota. Cell Host Microbe 24: 637-652, e1-e8. https://doi.org/10.1016/j.chom.2018.10.001
Virtue, A.T., McCright, S.J., Wright, J.M., Jimenez, M.T., Mowel, W.K., Kotzin, J.J., Joannas, L., Basavappa, M.G., Spencer, S.P., Clark, M.L., Eisennagel, S.H., Williams, A., Levy, M., Manne, S., Henrickson, S.E., Wherry, E.J., Thaiss, C.A., Elinav, E. and Henao-Mejia, J., 2019. The gut microbiota regulates white adipose tissue inflammation and obesity via a family of microRNAs. Science Translational Medicine 11: eaav1892. https://doi.org/10.1126/scitranslmed.aav1892
Wang, D., Xia, M., Yan, X., Li, D., Wang, L., Xu, Y., Jin, T. and Ling, W., 2012. Gut microbiota metabolism of anthocyanin promotes reverse cholesterol transport in mice via repressing miRNA-10b. Circulation Research 111: 967-981. https://doi.org/10.1161/circresaha.112.266502
Wang, Y., Yang, L.-Z., Yang, D.-G., Zhang, Q.-Y., Deng, Z.-N., Wang, K. and Mao, X.-J., 2020. miR-21 antagomir improves insulin resistance and lipid metabolism disorder in streptozotocin-induced type 2 diabetes mellitus rats. Annals of Palliative Medicine 9: 394-404.
Wang, Z., Zhou, H., Cheng, F., Zhang, Z. and Long, S., 2022. miR-21 negatively regulates the PTEN-PI3K-Akt-mtor signaling pathway in Crohn’s disease by altering immune tolerance and epithelial-mesenchymal transition. Discovery Medicine 34: 45-58.
Willeit, P., Skroblin, P., Moschen, A.R., Yin, X., Kaudewitz, D., Zampetaki, A., Barwari, T., Whitehead, M., Ramı́rez, C.M., Goedeke, L., Rotllan, N., Bonora, E., Hughes, A.D., Santer, P., Fernández-Hernando, C., Tilg, H., Willeit, J., Kiechl, S. and Mayr, M., 2017. Circulating microRNA-122 is associated with the risk of new-onset metabolic syndrome and type 2 diabetes. Diabetes 66: 347-357. https://doi.org/10.2337/db16-0731
Wortelboer, K., Bakker, G.J., Winkelmeijer, M., van Riel, N., Levin, E., Nieuwdorp, M., Herrema, H. and Davids, M., 2022. Fecal microbiota transplantation as tool to study the interrelation between microbiota composition and miRNA expression. Microbiological Research 257: 126972. https://doi.org/10.1016/j.micres.2022.126972
Wu, H., Tremaroli, V., Schmidt, C., Lundqvist, A., Olsson, L.M., Krämer, M., Gummesson, A., Perkins, R., Bergström, G. and Bäckhed, F., 2020. The gut microbiota in prediabetes and diabetes: a population-based cross-sectional study. Cell Metabolism 32: 379-390, e1-e3. https://doi.org/10.1016/j.cmet.2020.06.011
Wu, Y., Wang, C.Z., Wan, J.Y., Yao, H. and Yuan, C.S., 2021. Dissecting the interplay mechanism between epigenetics and gut microbiota: health maintenance and disease prevention. International Journal of Molecular Sciences 22: 6933. https://doi.org/10.3390/ijms22136933
Xue, H., Chen, X., Yu, C., Deng, Y., Zhang, Y., Chen, S., Chen, X., Chen, K., Yang, Y. and Ling, W., 2022. Gut microbially produced indole-3-propionic acid inhibits atherosclerosis by promoting reverse cholesterol transport and its deficiency is causally related to atherosclerotic cardiovascular disease. Circulation Research 131: 404-420. https://doi.org/10.1161/circresaha.122.321253
Yan, H., Zhang, X. and Xu, Y., 2020. Aberrant expression of miR-21 in patients with inflammatory bowel disease: a protocol for systematic review and meta analysis. Medicine 99: e19693. https://doi.org/10.1097/md.0000000000019693
Yan, X.Y., Yao, J.P., Li, Y.Q., Zhang, W., Xi, M.H., Chen, M. and Li, Y., 2022. Global trends in research on miRNA-microbiome interaction from 2011 to 2021: a bibliometric analysis. Frontiers in Pharmacology 13: 974741. https://doi.org/10.3389/fphar.2022.974741
Zhang, L., Hou, D., Chen, X., Li, D., Zhu, L., Zhang, Y., Li, J., Bian, Z., Liang, X., Cai, X., Yin, Y., Wang, C., Zhang, T., Zhu, D., Zhang, D., Xu, J., Chen, Q., Ba, Y., Liu, J., Wang, Q., Chen, J., Wang, J., Wang, M., Zhang, Q., Zhang, J., Zen, K. and Zhang, C.Y., 2012. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Research 22: 107-126. https://doi.org/10.1038/cr.2011.158
Zhang, R., Chen, Y.-N., Zhang, J. and Liu, J., 2023. Elevated serum levels of diamine oxidase, D-lactate and lipopolysaccharides are associated with metabolic-associated fatty liver disease. European Journal of Gastroenterology and Hepatology 35: 94-101. https://doi.org/10.1097/meg.0000000000002456
Zhang, T., Yang, Z., Kusumanchi, P., Han, S. and Liangpunsakul, S., 2020. Critical role of microRNA-21 in the pathogenesis of liver diseases. Frontiers in Medicine 7: 7. https://doi.org/10.3389/fmed.2020.00007
Zhou, X., Zhang, X., Zhao, N., Zhang, L., Qiu, W., Song, C., Chai, J., Cai, S. and Chen, W., 2023. Gut microbiota deficiency exacerbates liver injury in bile duct ligated mice via inflammation and lipid metabolism. International Journal of Molecular Sciences 24: 3180.


{kind=link}
{kind=link}
{kind=link}