Abstract
Many social animals acquire social behaviours during development, and social experience during development can be vital for acquiring necessary social behaviours in adulthood. We investigated the development of a distinctive tactile interaction behaviour in Bronze Cory catfish, in which adults interact with one another tactilely during foraging and during group responses to threats. We found that larvae respond to applied tactile stimulation with a flight response significantly less often as larvae matured. This habituation to tactile stimulation is consistent with developing appropriate adult social behaviour. We also found that social exposure affects the larval response to tactile interactions with conspecifics, and that isolation in early life leads to a greater likelihood of responding to tactile interactions with conspecifics with a flight response. This suggests that social exposure is important for developing social tactile interaction behaviour and underscores the particular importance of early experience in social development.
1. Introduction
Group living confers a number of advantages to social animals, including a reduced risk of predation (Magurran, 1990) and increased foraging efficiency (Pitcher et al., 1982). In order for social animals to reap the benefits of group living, individuals must coordinate their behaviour to their group-mates’. While the coordination of behaviours is necessary for social groups to function, this presents a developmental challenge, as most social animals begin their lives as socially undeveloped juveniles without the behavioural repertoire to interact with group-mates in ways that are appropriate for group structure and function (Mason, 1979). In order for these juveniles to become socially functional adults, juvenile social behaviour must emerge over time alongside physical maturation (Skinner, 1966).
Many behaviours necessary for group coordination, including social attraction (which leads to group cohesion), develop in groups alongside physical development in individuals. In the catfish Corydoras paleatus, the development of group cohesion and aggregative behaviours is highly correlated to anatomical developmental stage (Rodríguez-Ithurralde et al., 2014). The development of cohesive group behaviour with age has also been documented in zebrafish (Danio rerio), which begin to form increasingly cohesive shoals as individuals age (Buske & Gerlai, 2011). In Florida scrub jays (Aphelocoma coerulescens), mobbing behaviour, a group behaviour used to deter predators, first appears in the fledgling stage and gradually develops into adult mobbing behaviour over about three months (Francis et al., 1989).
While time is necessary for all developmental changes, many factors affect how juveniles grow into adults. For the development of social behaviours, the social environment an animal experiences during development can have a profound impact on the social behaviours it exhibits as an adult (Slagsvold et al., 2002). This is apparent in species that imprint, such as when very early experiences determine mate preferences in great tits (Slagsvold et al., 2002). In zebrafish, a preference to shoal with conspecifics with particular coloration patterns is highly influenced by the patterns of the group-mates that an individual was raised with (Spence & Smith, 2007). The way social behaviour can be determined by early experiences highlights how the mechanisms of development can affect behaviour later in life. In rats (Rattus norvegicus), for example, individuals who experience early social isolation subsequently develop a behavioural condition which shares core features with schizophrenia (Fone & Porkess, 2008). Furthermore, many species have ‘critical periods’ in which developing individuals must experience specific stimuli in order to develop adult behaviours. Examples include language development in humans, in which individuals must be exposed to language within a window of development to fully develop language abilities (Kuhl et al., 2005) and song tuition in zebra finches (Taeniopygia guttata), which must occur within a specific period of development for a young bird to properly learn to sing (George et al., 1995).

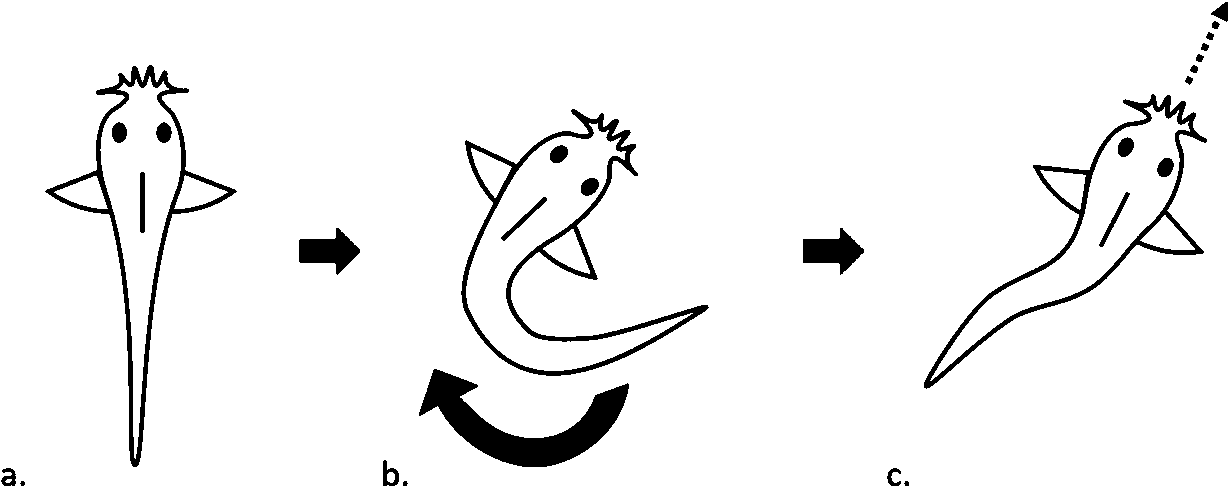
C-start Schematic Diagram, dorsal view. (a) Larva pre C-start; (b) Stage 1, larva adopts ‘C shape’ during the fast phase; (c) Stage 2, larva straightens body and is propelled forwards in an escape trajectory.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
The development of communication in social species is of particular interest, as communication can be a driving factor of group coordination (Conradt & Roper, 2005). Adults of our study species, Corydoras aeneus (Bronze Cory catfish), utilize tactile interactions that seem to facilitate group coordination (Riley et al., 2019b) and mediate flight responses from a potential threat (Riley et al., 2019a). The usage of tactile interactions in Bronze Cory catfish is unusual, as tactile stimulation often triggers a stereotyped threat response in fish called a ‘c-start’ that is deployed involuntarily when an individual perceives an urgent potential threat (Kimmel et al., 1980). The c-start threat response was first reported in detail by (Weihs, 1973) in trout (Salmo trutta) and pike (Esox lucius) and is characterized by a two stage motor pattern (Figure 1). Stage 1 lasts 15–40 ms and is characterized by the ipsilateral contraction of axial muscle on one side of the body (Eaton & Emberley, 1991). The fish orientates away from the threat and resembles a ‘C shape’ from above at the end of stage one. The head and caudal fin flap lie in the same direction. Stage 2 is characterized by the straightening of the axial skeleton and acceleration caused by the lateral side of the fish displacing water (Eaton et al., 1988). This allows the fish to be propelled forwards in an escape trajectory. The underlying neural command appears to be ballistic, and the reflex is unaffected by sensory information once it is initiated (Eaton and Emberley, 1991). The c-start is particularly important for larval fish, and larvae and adults of a related species, Corydoras paleateus, exhibit well-developed Mauthner cells and circuits, which trigger the c-start response (Rodríguez-Ithurralde et al., 2014). We have also observed adult Bronze Cory catfish undergoing a c-start response while responding to a threat stimulus (Riley et al., 2019a). The fact that Bronze Cory catfish respond to potential threats with a c-start response, but also interact tactilely with conspecifics without a c-start response, suggests that Bronze Cory catfish may develop a tolerance to tactile stimulation during development. Developing this tolerance would allow larvae to respond to tactile stimulation as a potential threat during early development (while larvae are most vulnerable and have not developed social cohesion) and eventually interact tactilely with conspecifics later in development as social aggregation behaviours develop (Buske & Gerlai, 2011; Rodríguez-Ithurralde et al., 2014).
Furthermore, Bronze Cory catfish provides a unique study system for investigating the development of social interaction, as patterns of tactile interactions can be observed and responses to tactile interactions can be clearly recorded. This allows us to ask questions about the way individual larvae respond to tactile interactions during development, and about the role of social exposure in developing a tolerance to tactile stimulation. How do individuals develop the ability to interact and coordinate with others, and how do their experiences with conspecifics during early development affect their responses to tactile interactions later on?
We conducted two experiments to investigate the development of communication and sociality in larval Bronze Cory catfish. Our previous work on adult Bronze Cory catfish describes an unusual tactile interaction behaviour, which we termed nudges, that individuals use during coordinated movements (Riley et al., 2019b) and while coordinating a group flight response (Riley et al., 2019a). This behaviour has several advantages for the study of group coordination, as nudges are discrete, easily quantifiable, and directed interactions between individuals: for each nudge, we can identify the initiator (i.e., the individual whose approach resulted in physical contact) and the recipient of the interaction. This behaviour is associated with increased coordination, with higher rates of nudging when fish are coordinating with one another when compared to fish that are in close proximity but not coordinating their movements (Riley et al., 2019b). Wild fish were observed utilizing this behaviour in several small streams in the Madre de Dios locality of the Peruvian Amazon in 2011 and 2013, particularly when fleeing the observer (RJR, pers. obs.), and captive individuals can frequently be seen nudging one another (RJR, BRG, TPR, AM, pers. obs.).
To investigate the manifestation of this behaviour during development, we first investigated the ontogeny of the response to tactile stimuli by testing individuals within stable groups over regular intervals in the developmental period. If the adult response to tactile stimulation (i.e. a social response and not a c-start) is the result of developmental processes, we predicted that older larvae would respond to the tactile stimulation with a c-start less often than younger larvae, demonstrating the effect of social development. We also predicted that individuals that did respond to experimental tactile stimulation with a flight response would be more likely to initiate tactile interactions with group-mates later in development as larvae developed the ability to utilize nudging behaviour to communicate about threats, as is seen in adult Bronze Cory catfish (Riley et al., 2019a). Second, we investigated the effect of isolation on sociality and communication in larvae. We predicted that isolated larvae would spend less time together, participate in fewer tactile communication interactions and be more likely to respond to tactile interactions with conspecifics with a c-start, demonstrating that they were not responding to tactile interactions socially.
2. Methods
2.1. Study system
Corydoras is a genus of highly social neotropical catfish widely available in the aquarium trade. In the wild, Bronze Cory catfish forage socially in mixed groups of males, females and juveniles (Lambourne, 1995), but their life histories are not fully known. In captivity, they have lifespans from 10–15 years (Lambourne, 1995). Corydoras tend to be benthic fish that live in slow moving, shallow water; they are known for their marked sociality and robust shoaling behaviour (Nijssen in Lambourne, 1995). Because they are bottom-dwelling, they shoal along the substrate in two dimensions and their social behaviour can be accurately recorded from above. Bronze Cory catfish have a subtle sexual dimorphism, and females are generally larger and thicker-bodied than males.
2.2. Study subjects: breeding Bronze Cory catfish
We obtained larval Bronze Cory catfish for use in these experiments by rearing eggs laid by our laboratory adult stock. The adult stock was originally obtained from three local pet shops in Cambridgeshire: Maidenhead Aquatics Cambridge, Pet Paks, and Ely Aquatics and Reptiles. Fish were maintained in reverse osmosis (RO) water purified to 15 or less total dissolved solids (TDS) and re-mineralized to 105–110 ppm TDS using a commercially prepared RO re-mineralizing mix (Tropic Marin Re-mineral Tropic). The fish experienced a 12:12 light:dark cycle under fluorescent ambient room lighting at a temperature of 23 ± 1°C. These adults were the parents of the larvae used in all experiments.
Adults were maintained under these conditions for at least one year before we collected eggs. We can be certain that all eggs from the same clutch are full siblings due to the Bronze Cory catfish’s unique copulation mechanism. During copulation, females drink sperm from one male which then fertilizes their entire clutch, which is laid in small clusters of full sibling eggs (Kohda et al., 1995). We collected the eggs within 24 h of spawning and removed them from the adult tanks to hatch and be reared under experimental conditions. Experiments occurred eight months apart, and different larvae were used in each experiment (although all larvae were the offspring of our adult stock population). Experiment-specific details of housing and husbandry are provided below.
2.3. Experiment 1: development of response to tactile stimulation
2.3.1. Experiment 1 husbandry
This experiment was performed in two batches, with two weeks in between batches. Larvae from each batch were taken from the same three adult stock tanks. For each batch, six groups, each composed of five full siblings 12 days post hatching (dph), were placed in a transparent, 20-l plastic fish tank. Each tank had a small air driven aquarium filter for biological filtration. Larvae were fed “Interpet Liquifry Number 1” and “Interpet Liquifry Number 3” twice daily. Aufwuchs, the biologically rich film that grows on the surface of aquariums and consists of microorganisms that are nutritionally beneficial for larvae, from adult Bronze Cory catfish tanks was introduced for larvae to graze on. In addition, larvae were fed twice a week with live Panagrellus redivivus.
2.3.2. Tactile stimulation procedure
Larvae were stimulated tactility on 14, 24 and 31 dph, and their responses were filmed from a dorsal perspective (i.e., from above). At 14 dph, our larvae were approximately 10–11 mm; at 24 dph, our larvae were approximately 14–15 mm; at 31 dph, our larvae were approximately 16–17 mm. These sizes are generally in line with the growth curves presented in (Huysentruyt et al., 2009). These videos were then scored to analyse larval behaviour. Larvae were not fed prior to tactile stimulation (but were fed afterwards). The order in which the groups were filmed was randomly determined, and larval tanks were physically moved to a filming set-up. To optimize filming, before tactile stimulation commenced, the water level in the tanks was reduced to 11 cm, and the air driven filter was removed as was any detritus on the bottom of the aquarium. Following these adjustments, the larvae were left in the filming set-up for 15 min to allow them to acclimatize.
After the acclimatization period, a 25-cm-long glass rod which tapered to a 1 mm blunt end was used to tactilely stimulate the larvae. Tactile stimulation was applied by RJR and TPR, and stimulation was performed in rounds. Each of the five fish in a group was stimulated in turn during a round before the next round commenced, except when a fish was previously nudged by a group-mate who had been stimulated earlier in the round and responded to the stimulation with a flight response. Individuals could be distinguished within tactile stimulation rounds, but due to larval movement following stimulation, could not always be individually identified between rounds. Rounds were separated by a period of at least 60 s to avoid stressing the larvae excessively. Stimulation was standardized through practice on a test cohort, which was used to develop the tactile stimulation protocol and from which no data were collected, before the experiment began. This ensured that the tactile stimulations were consistent and reproducible when data were collected. Tactile stimulation was delivered to the anterior half of the body.
Each group of larvae underwent a session of 10 or 11 rounds of tactile stimulation; each individual received tactile stimulation at maximum once per round. In some cases, after we tactilely stimulated a larva, it proceeded to initiate a nudge with one of its group-mates; in this case, we did not apply tactile stimulation to the receiver of the nudge during that round of tactile stimulation (and resumed the following round). This was to ensure that receiving a nudge from a group-mate did not bias the larva’s response to the experimental tactile stimulation. In sessions where one or more rounds resulted in only one to three individuals receiving artificial stimulation, an 11th round was performed. We aimed to stimulate all individuals as consistently as possible (and at least three times per session), but because larvae could not be distinguished definitively between rounds, we cannot be certain we reached this criteria.
Stock tanks containing full siblings for all the experimental groups were maintained under identical conditions so that any deceased larvae could be replaced with full siblings. Over the course of the experiment, 11 larvae (out of the 60 that began the experiment) died (of either apparent deformities, or no obvious cause) in seven different groups, and were replaced with full siblings from the same clutch at least 24 h before tactile stimulation was applied. The mortality in the larvae taking part in this experiment was very similar to the mortality of full siblings not participating in the experiment.
To determine if any developmental effects we observed in larvae were also present in individuals who had never experienced our tactile stimulation procedure, we also applied a modified tactile stimulation procedure to naïve adults from our laboratory stock population. Because adults were significantly bigger than even 31 dph larvae, we tested 11 groups of three adults in a 45.5 × 25 × 21 cm aquarium. We applied tactile stimulation with the rounded end of a 25-cm glass rod.
2.3.3. Tactile stimulation scoring procedure
We scored the immediate and delayed response of each larva to tactile stimulation. We defined the immediate response as occurring within the first second after stimulation, and each immediate response was the larvae (1) ‘ignoring’, (2) c-starting, or (3) performing a non c-start flight response. A response was scored as ‘ignored’ if the larva being stimulated remained static; larvae with an immediate ‘ignoring’ response did not have a further delayed response. A c-start was scored if a clear c-shaped body morphology was observed in conjunction with rapid movement. A non-c-start flight response was scored if the larva did not remain static in response to tactile stimulation but also did not undergo a c-start. For adults, we could only score the ‘immediate’ response to the tactile stimulation due to the relatively much greater size of the adults relative to the testing aquarium.
For larvae, only larvae with an immediate response of (2) or (3) could have a delayed response, and we defined the delayed response as occurring within 1–5 s of stimulation; larvae that ignored the tactile stimulus remained still and hence did not undergo a delayed response. The delayed response could be either (1) nudging another larvae or (2) movement around the aquarium that did not involve nudging another larvae. We recorded this measure to ascertain if flight responses tended to lead to nudges more frequently as larvae developed socially.
Videos were scored by RJR and TPR, and scoring protocols were developed to ensure consistency between researchers. Both researchers observed one full session of tactile stimulation (10 rounds of stimulation) together to develop the scoring protocol. Four training videos were then scored independently by both researchers. Scores were compared, and inconsistencies resolved by referring to the scoring protocol and reaching a consensus about each interaction. A further two videos were scored independently and the scores of each measure were compared. All measures (for both immediate response and delayed response) were within 90% agreement between both scorers and so the scoring was assessed as being consistent. The remaining videos were randomly divided up between the two researchers and were scored independently. Due to a filming error, two groups were excluded from the first batch, and therefore actual analysis was carried out on 10 groups total.
2.4. Experiment 2: effect of isolation on sociality and communication
2.4.1. Experiment 2 husbandry
To investigate the effect of social isolation on sociality and communication, larvae were reared in groups of 3–5 or in isolation from hatching until testing. All larvae were raised in identical mesh enclosures of roughly 9.5 cm diameter suspended in 20-l aquariums equipped with an Interpet MINI filters. Each 20-l aquarium held a maximum of six mesh enclosures. Larvae were fed twice daily with “Interpet Liquifry Number 1” and “Interpet Liquifry Number 3” daily. Aufwuchs from adult Bronze Cory catfish tanks was introduced for larvae to graze on.
2.4.2. Filming procedure
After being reared either socially or in isolation, larvae in the age range of 14–21 dph were tested in groups of three; seven groups of socially-housed larvae were tested, and eight groups of isolation-housed larvae were tested. During filming, all larvae were moved from their mesh enclosures and placed in a standard breeding net (TRIXIE brand, 16 × 13 × 12 cm), which consisted of a plastic frame fitted with fine white mesh that could safely contain larvae on a more visible background closer to the camera. Larvae in the social-housing condition were placed with the other larvae in their mesh enclosures; larvae in the isolation-housing condition were placed with other isolation-housed larvae from the same 20-l aquarium. Due to their similar and diminutive size, larvae could not be individually recognized. After a 30-min acclimatization period, larval behaviour was filmed dorsally for a 1-h period. The camera was fixed 30 cm above the water surface. Larvae were not fed prior to filming (but were fed after). Following filming, larvae were removed from the breeding nets and placed in larger (30 l) communal housing tanks.
2.4.3. Video scoring
We recorded measures of cohesion and nudging from each 1-h video; details are below.
Larvae were scored as ‘together’ if they were within 2 body lengths of each other using the body length of the smallest fish in that group. Time was recorded as soon as two fish were within this distance and were on the bottom. Periods of togetherness (i.e., cohesion) were terminated if one fish left the bottom of the breeding net and began swimming up and down along the sides, as it is not possible to tell how high the larvae were swimming and, consequently, the distance between larvae. As a result, larvae could only be classified as being ‘together’ if they were on the bottom of the breeding net. If a fish briefly moved more than two body lengths away from its partner(s) but returned to within 2 body lengths in less than three seconds, then this was scored as one continuous period of togetherness without a break for the brief separation period. Periods of togetherness were only scored if individuals were together for at least 3 s.
We defined a ‘nudge’ as an event when one fish physically touches another (Riley et al., 2019b). As with cohesion, nudges were only scored if both fish were on the bottom of the enclosure. Each nudge had an initiator and a receiver. An initiator is the individual whose movement resulted in the interaction. A receiver is the individual who was touched by the movement of the initiator. We recorded the response of initiators and receivers to tactile interactions. Responses were classified as either (1) a c-start or (2) not a c-start. A c-start was scored if a clear c-shaped body morphology was observed in conjunction with rapid movement, as is characteristic of the fast phase of the c-start response.
2.4.4. Other measures
In addition to initiator and receiver c-starts, we also scored spontaneous c-starts. A spontaneous c-start was defined as any time an individual exhibited a c-start response (identified by the clear c-shaped body morphology and rapid movement) without any apparent contact from another larvae.
2.4.5. Assessing the scoring protocol
To assess the consistency of scoring protocol, we developed an ethogram to score the interactions between group-members. Videos where scored by RJR and TPR. Scoring always began 30 min after the start of filming. Both researchers observed an hour of randomly selected footage together to develop scoring protocol for an ethogram. Three training videos were then scored independently by both researchers. Scores were compared, and inconsistencies resolved by referring to the scoring protocol and reaching a consensus about each interaction. A further three videos were scored independently and the scores of each measure were compared. All measures (total time together, total number of interactions, total initiator c-starts, total receiver c-starts, total spontaneous c-starts) were within 90% agreement between both scorers and so the scoring was assessed as being consistent. The remaining videos were randomly divided between the two researchers and were scored independently. Videos were scored blind (i.e., the researchers did not know if a given group was from the social or isolation housing condition).
2.4.6. Data analyses
All statistical analyses were carried out in R version 3.2.2 (R core developer team). Data distributions were initially assessed visually and model diagnostics were subsequently checked to assure appropriate fits.
We investigated the development of tactile stimulation by comparing the proportion of tactile stimulation events that resulted in a c-start, in ignoring responses, and in social interactions among different age groups. Binomial and betabinomial Generalized Linear Mixed-effects Models (GLMM) were used for proportion data as groups were tested three times at different ages; we fitted binomial GLMMs using the lme4 package (Bates et al., 2015) and betabinomial distributions using the glmmTMB package where binomial GLMMs were overdispersed (Brooks et al., 2017). We used single-term deletions with
We fitted a betabinomial GLMM with the proportion of c-starts (the number of tactile stimulation events resulting in a c-start in each group in a session divided by the total number of tactile stimulation events in that group in a session) as the response variable, age (14, 24 and 31 dph) as the fixed effect, and group ID was a random effect. We modelled the proportion of non-response (“ignoring” responses) with a betabinomial GLMM with the proportion of ignoring responses (the number of tactile stimulation events resulting in the fish not responding to the stimulation in a group per session divided by the total number of tactile stimulation events in a group per session) as the response variable, age (14, 24 and 31 dph) as the fixed effect, and group ID as a random effect. Similarly, we used a GLMM with betabinomial error structure to assess if individuals were more likely to nudge conspecifics during a flight response as they aged. The proportion of tactile stimulation events where an individual responded to the stimulus by interacting tactilely with a group-mate(s) (i.e., the number of tactile stimulation events where the individual being stimulated initiated a nudge with a group-mate following stimulation in a group per session, divided by the number of tactile stimulation events where the individual moved in response to tactile stimulation in a group per session) was our response variable, age (14, 24 and 31 dph) was the fixed effect, and group ID was a random effect. We performed a Tukey post-hoc test on all three of these models (using the eemeans package in R) to confirm which ages differed in c-start and ignoring responses as well as the likelihood of initiating nudges during flight responses.
To assess if any development patterns observed were also present in individuals who had never been exposed to the tactile stimulation procedure, we used a nonparametric Wilcoxon signed-rank test (as the data were not normally distributed) to compare the proportion of c-starts between 31-dph larvae and naïve adult fish. We used a two-sample t-test to compare the proportion of ignoring responses between 31-dph larvae and naïve adult fish.
For the social isolation experiment, we fitted linear models (LMs) for normally distributed data, GLMs with Poisson error distribution for count data, GLMs with negative binomial error distributions where Poisson models had been overdispersed (Bolker et al., 2009), and GLMs with quasibinomial error distribution for proportion data.
To assess whether socially- and isolation-housed groups differed in the number of total tactile interactions each group underwent, we fitted a GLM with a Poisson error structure: the total number of interactions between group members was the response variable and social housing status and age of the larvae were explanatory variables. We used a linear model (LM) to assess whether socially-housed and isolation-housed groups differed in the amount of time individuals were in proximity to one another: time together was the response variable, and social housing status and age of the larvae were explanatory variables.
To assess whether social isolation affected how individuals responded to initiating and receiving nudges, we used two quasibinomial GLMs. First, the proportion of interactions in which the initiator c-started following the interactions (the number of interactions where the initiator c-started following the interaction divided by the total number of interactions the group underwent) was the response variable and age and social housing status were the explanatory variables. Second, the proportion of interactions in which the receiver c-started following the interaction (the number of interactions where the receiver c-started following the interaction divided by the total number of interactions the group underwent) was the response variable and age and social housing status were the explanatory variables.
Finally, to assess if social housing status affected the likelihood of c-starting outside of social interactions, we ran a negative binomial GLM with the number of spontaneous c-starts as the response variable and social housing status and age as the explanatory variables.
2.5. Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. This study was approved via a non-regulated use of animals in scientific procedures application (consistent with UK’s animal welfare legislation ASPA) through the University of Cambridge. It was approved through the University of Cambridge’s Ethical Review Process; it was approved and presented by the Named Veterinary Surgeon and the Named Animal Care and Welfare Officer (NACWO) for the Zoology Department.
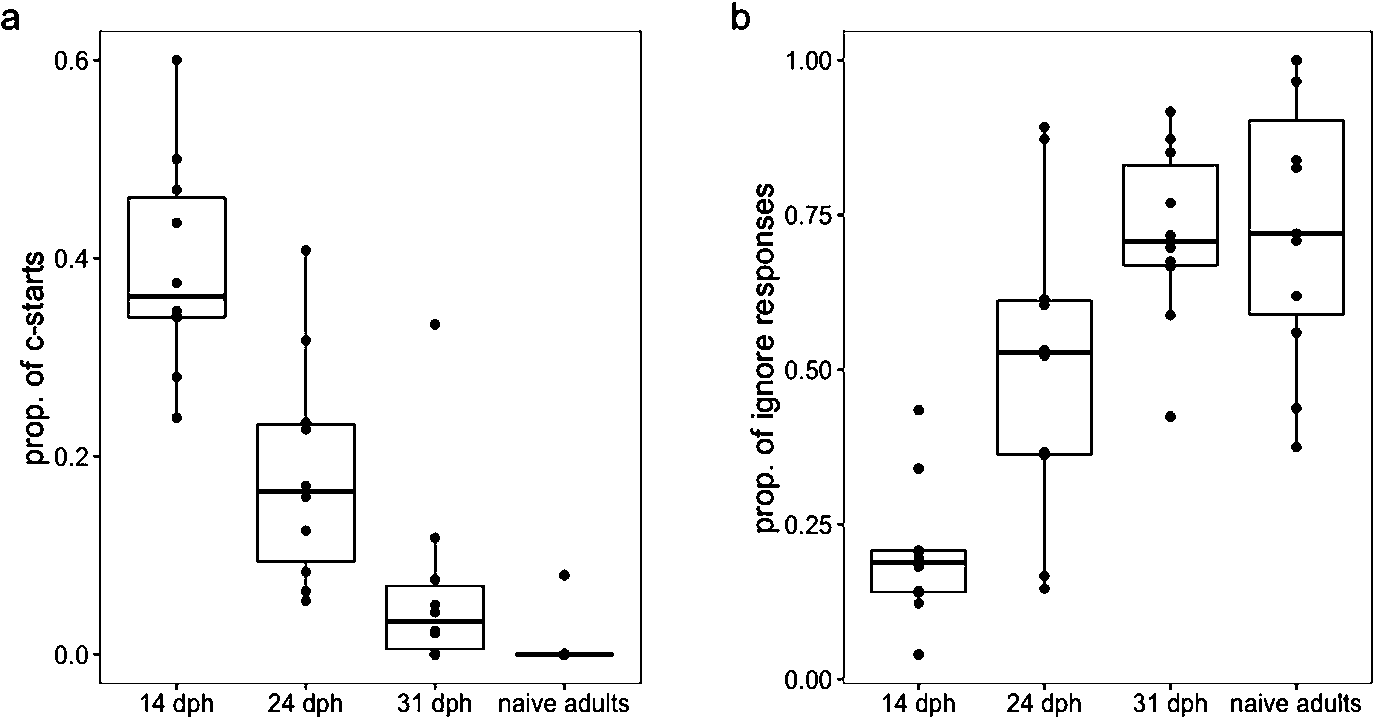
(a) The proportion of tactile stimulation events that resulted in a c-start response in larvae days 14, 24 and 31 dph; (b) the proportion of tactile stimulation events that results in an ignoring response on days 14, 24 and 31 dph.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
3. Results
3.1. Development of response to tactile stimulation
Larval responses to tactile stimulation varied by age as detailed below. The proportion of responses to tactile stimulation that were not c-starts or ignoring responses (i.e., the number of non c-start responses to tactile stimulation events divided by the total number of tactile stimulation events) were 0.41, 0.31 and 0.22 in larvae ages 21, 24 and 31 dph, respectively. The remainder of our analysis focused on the proportion of ignoring and c-start responses.
Larvae responded to tactile stimulation with a c-start response at lower rates as they mature (betabinomial GLMM: Likelihood Ratio Test (LRT) = 34.5,
As larvae mature, they are more likely to initiate a nudge with their group-mates when they do respond to a tactile stimulation event with a flight response (betabinomial GLMM: LRT = 19.2,
The proportion of responses involving one or more nudges (number of tactile stimulation events in which a larva nudged a group-mate divided by the number of tactile stimulation events in which larvae responded to the tactile stimulation with a c-start or non c-start flight response) in larvae 14, 24 and 31 days post hatching.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
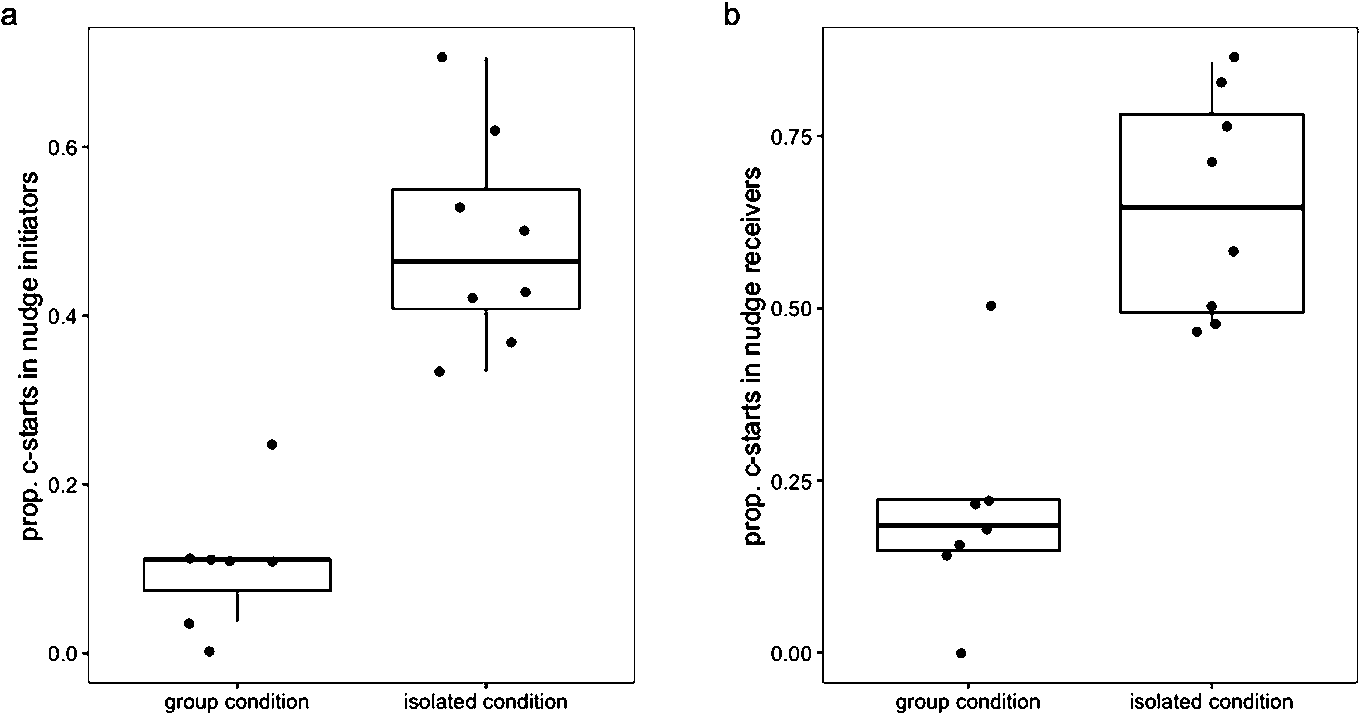
(a) Total nudges in groups of three larvae reared socially and larvae reared in isolation; (b) time together in larvae reared socially and larvae reared in isolation.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
3.2. The role of social exposure in responding to tactile interactions with group-mates
Housing condition (socially housed or isolation housed) was a significant predictor of how many nudges were initiated within groups, with isolation housed groups initiating fewer nudges (Poisson GLM: LRT = 12.9,
Larvae raised in isolation are also significantly more like to respond to a nudge with a c-start both when initiating (quasibinomial GLM:
(a) Proportion of total interactions in which the initiator c-started following a nudge; (b) proportion of total interactions in which the receiver c-started following a nudge.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
However, it does not seem that individuals in the isolation condition were merely more reactive generally: the social- and isolation-reared groups performed a similar number of spontaneous c-starts (negative binomial GLM: LRT = 0.36,
The number of spontaneous c-starts in socially- and isolation-reared groups.
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
4. Discussion
Our results show that Bronze Cory catfish larvae are more likely to c-start in response to tactile stimulation when in early developmental stages, and increasingly tolerate tactile stimulation as they develop. Our results also suggest that more developed larvae tend to initiate tactile interactions with conspecifics when disturbed by a tactile stimulation event. Our results also show that social isolation during development leads to fewer tactile interactions and increased likelihood of responding to tactile interactions with conspecifics with a flight response. This is consistent with development of social interaction behaviour in other taxa, even though the Bronze Cory catfish’s tactile mode of social interaction seems to be less commonly reported than visual, auditory, or olfactory social interactions; consequently, this study also adds to the body of literature on social development by demonstrating commonalities across taxa and sensory modalities.
Increased toleration of tactile stimulation imply that the larvae may have perceived the stimulation as a threat less often with age, may have already developed the ‘freeze’ threat response common in adult threat responses, or their morphological development could have prevented them forming the c-shape necessary for a c-start to be scored. Morphological constraints seem unlikely, as there were a few c-starts in the adult fish stimulated in this study, as well as in adult fish in our previous work (Riley et al., 2019). It also seems unlikely that larvae would adopt a freeze threat response before the development of prominent anterior pectoral fin rays or armoured scutes (which occurs at >22 mm in Corydoras arcuatus, a related species of similar size (Sire, 1993)) because they have very little protection from predation (Rodríguez-Ithurralde et al., 2014) and would rely on camouflage alone. Therefore, it seems that the most likely explanation for the change in response to tactile stimulation is that older larvae were less likely to perceive tactile stimulation as a threat. The ability to control and modulate c-start responses has been observed in archer fish (Toxotes jaculatrix) in the context of feeding (Wöhl & Schuster, 2007) and it may be that the modulation of the c-start reflex occurs in Bronze Cory catfish so that tactile interaction is tolerated.
The fact that older larvae tend to initiate more tactile interactions with group-mates when they do respond to tactile stimuli is likely to be due, at least in part, to having greater social exposure with more advanced development. This is consistent with the results of the isolation experiment, in which larvae housed in groups (and which therefore have ample opportunities to engage in tactile social interactions with group-mates) were far less likely to respond to a tactile interaction with a group-mate with a c-start response, assuming our results primarily reflect the effect of isolation. An alternative explanation for these results is the differing degrees of social familiarity, as individuals housed socially were tested with their social housing group-mates. Isolation housed individuals, however, were always tested with other isolated individuals from their 21 l aquarium, which allowed for constant exposure to olfactory cues based on shared habitat and diet. As these olfactory cues have shown to be a key factor in the formation of familiarity in fish (Ward et al., 2004), and larvae had visual access to others through the mesh, our methodology allowed for familiarity based on diet, habitat, and chemical cues to form, and only kept isolation housed larvae in tactile isolation from one another. For this reason, it seems likely that our results are not based on familiarity or lack thereof, but rather the effect of social tactile exposure and social tactile isolation.
Social exposure has been found to influence and ultimately weaken responses to potential threats in paradise fish (Macropodus opercularis) larvae (Miklósi et al., 1997), as we saw with the developing larvae in our tactile stimulation experiment. It seems likely that paradise fish larvae became habituated to the continuous presence of larval conspecifics and generalized the experience of conspecifics with exposure to potential predators, leading to a weakened response to predators with increasing social exposure (Miklósi et al., 1997). We observed a similar decrease in response to a potential threat with age in Bronze Cory catfish larvae, which was also likely a result of increased social exposure. It may be that desensitization to tactile stimulation during early development in Bronze Cory catfish may only occur in the presence of conspecifics to ensure that individuals do not erroneously ignore a potential threat. Nonetheless, the fact that older Bronze Cory catfish larvae were more actively social (i.e. initiated more nudges with group-mates) when they did mount a flight response to experimentally applied tactile stimulation implies that the effect of social exposure in Bronze Cory catfish is not just limited to the downregulation of a threat response, as it appears to be in paradise fish larvae.
Although our experiment cannot elucidate the long-term effects of social isolation, the isolated larvae’s increased likelihood of misinterpreting an interaction with a conspecific has parallels in other systems. The behavioural and neurological effects of social isolation early in development are well documented in rodent models (Einon & Morgan, 1977; Fone & Porkess, 2008; Makinodan et al., 2012). Impaired sensorimotor gaiting was a symptom of early social isolation in rats (Fone & Porkess, 2008) and this is one possible explanation for the higher c-start frequency we observed in isolated larvae. In many animals, including rodents (Fone & Porkess, 2008), social isolation early in development permanently alters an individual’s social behaviour, rendering individuals incapable of normal social behaviour or cognition. This effect is seen in invertebrates as well: in mites, early isolation impairs social competency and mate choice selectivity (Schausberger et al., 2017), and social isolation during development leads to the inability to perform the essential social task of facial recognition in paper wasps (Tibbetts et al., 2019). In mice, social isolation in early development has a lasting effect on brain development, but only if mice are isolated during their critical period in development (Makinodan et al., 2012). Because critical periods are so fundamental to the development of social behaviour in many group-living animals, the study of social isolation and its effects in more diverse taxa may have the potential to improve our understanding of some psychological and behavioural disorders, and the role social exposure plays in their aetiology. Indeed, in humans, social isolation is as significant a risk factor as smoking or obesity for morbidity and mortality, effects that have also been observed in animal models (Cacioppo et al., 2011).
With social exposure playing a role in the development of adult-like behaviour in Bronze Cory catfish larvae, this implies that there may be a critical period during which individual social behaviour develops through social exposure. In the tactile stimulation experiment, we found the decrease in c-start frequency seemed to occur gradually over time, with 14 dph larvae c-starting frequently, 24 dph larvae c-starting significantly less frequently, and 31 dph larvae c-starting very seldomly at all. This progression seems gradual over this time period, without a developmental ‘switch’ that very rapidly modifies behaviour, but this window of time may represent a critical period for social skill acquisition. We also note that this gradual decrease in frequency of c-starting response and increase of ignoring response could be the result of larval acclimation to the tactile stimulus, and that larvae may have adjusted their alarm responses as they habituated to the tactile stimulation procedure. However, our result that naïve adult fish ignore the stimulus at similar rates to 31 dph larvae, and c-start even less frequently than 31 dph (indicating continued development of desensitization to tactile stimuli), suggests that the developmental progression we observed was not solely due to habituation to our tactile stimulation procedure. Furthermore, our result that isolated larvae in the isolation experiment exhibited significantly more c-starts in response to nudges from conspecifics strongly suggests that sustained social exposure leads to the development of toleration for tactile stimulation. These results show the potential of nudging in Corydoras catfish as model for social development.
Our isolation experiment provides evidence that nudging requires social exposure to properly develop, although it is unclear whether or not there is a critical period of social exposure for the development of nudging. In our isolation experiment, the isolation of larvae was confounded when they formed groups of three for filming and the development of isolated individuals was not followed post filming due to the difficulty of tagging larvae. Future work may aim to determine if the effect of social isolation we observed in isolated larvae can be overcome by social exposure later in life or if Bronze Cory catfish larvae show a critical period for social exposure. Social behaviour may be plastic throughout a fish’s life, or it could be determined by the early developmental environment. If there is a critical period for development of sociality in Bronze Cory catfish then it is ecologically important for an individual to have social exposure and interact with other larvae during that critical period. If a larva does not, then it may become unable to interact effectively with the rest of the shoal later in life, potentially misinterpreting the social interactions of conspecifics as a threat.
Their readily observable tactile interaction behaviour and response paradigm make Bronze Cory catfish a compelling model system for investigating the development of social behaviour and its consequences in later life. Our results emphasize the importance of social experience during development in adult social behaviour, and further work on the Bronze Cory catfish can further elucidate the ecological and social consequences of early social experiences for social behaviour in adulthood.
Corresponding author’s e-mail address: rjriley@post.harvard.edu
Acknowledgements
We would like to acknowledge Jim Allen for his logistical support and comments on the manuscript, as well as Arne Jungwirth for his statistical advice and comments on the manuscript.
References
Bates, D., Mächler, M., Bolker, B. & Walker, S. (2015). Fitting linear mixed-effects models using lme4. — J. Stat. Softw. 67: 1-48. DOI:10.18637/jss.v067.i01.
Bolker, B.M., Brooks, M.E., Clark, C.J., Geange, S.W., Poulsen, J.R., Stevens, M.H.H. & White, J.-S.S. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. — Trends Ecol. Evol. 24: 127-135. DOI:10.1016/j.tree.2008.10.008.
Brooks, M.E., Kristensen, K., van Benthem, K.J., Magnusson, A., Berg, C.W., Nielsen, A., Skaug, H.J., Machler, M. & Bolker, B.M. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. — R J. 9: 378-400. DOI:10.3929/ethz-b-000240890.
Buske, C. & Gerlai, R. (2011). Shoaling develops with age in Zebrafish (Danio rerio). — Prog. Neuropsychopharmacol. Biol. Psych. 35: 1409-1415. DOI:10.1016/j.pnpbp.2010.09.003.
Cacioppo, J.T., Hawkley, L.C., Norman, G.J. & Berntson, G.G. (2011). Social isolation. — Ann. N.Y. Acad. Sci. 1231: 17-22. DOI:10.1111/j.1749-6632.2011.06028.x.
Conradt, L. & Roper, T.J. (2005). Consensus decision making in animals. — Trends Ecol. Evol. 20: 449-456. DOI:10.1016/j.tree.2005.05.008.
Eaton, R.C., DiDomenico, R. & Nissanov, J. (1988). Flexible body dynamics of the goldfish C-start: implications for reticulospinal command mechanisms. — J. Neurosci. 8: 2758-2768.
Eaton, R.C. & Emberley, D.S. (1991). How stimulus direction determines the trajectory of the Mauthner-initiated escape response in a teleost fish. — J. Exp. Biol. 161: 469-487.
Einon, D.F. & Morgan, M.J. (1977). A critical period for social isolation in the rat. — Dev. Psychobiol. 10: 123-132. DOI:10.1002/dev.420100205.
Fone, K.C.F. & Porkess, M.V. (2008). Behavioural and neurochemical effects of post-weaning social isolation in rodents — relevance to developmental neuropsychiatric disorders. — Neurosci. Biobehav. Rev. 32: 1087-1102. DOI:10.1016/j.neubiorev.2008.03.003.
Francis, A.M., Hailman, J.P. & Woolfenden, G.E. (1989). Mobbing by Florida scrub jays: behaviour, sexual asymmetry, role of helpers and ontogeny. — Anim. Behav. 38: 795-816. DOI:10.1016/S0003-3472(89)80112-5.
George, J.M., Jin, H., Woods, W.S. & Clayton, D.F. (1995). Characterization of a novel protein regulated during the critical period for song learning in the zebra finch. — Neuron 15: 361-372. DOI:10.1016/0896-6273(95)90040-3.
Huysentruyt, F., Moerkerke, B., deVaere, S. & Adriaens, D. (2009). Early development and allometric growth in the armoured catfish Corydoras aeneus (Gill: 1858). — Hydrobiologia 627: 45-54. DOI:10.1007/s10750-009-9714-z.
Kimmel, C.B., Eaton, R.C. & Powell, S.L. (1980). Decreased fast-start performance of zebrafish larvae lacking mauthner neurons. — J. Comp. Physiol. 140: 343-350. DOI:10.1007/BF00606274.
Kohda, M., Tanimura, M., Kikue-Nakamura, M. & Yamagishi, S. (1995). Sperm drinking by female catfishes: a novel mode of insemination. — Environ. Biol. Fishes 42: 1-6. DOI:10.1007/BF00002344.
Kuhl, P.K., Conboy, B.T., Padden, D., Nelson, T. & Pruitt, J. (2005). Early speech perception and later language development: implications for the “critical period. — Lang. Learn. Dev. 1: 237-264. DOI:10.1080/15475441.2005.9671948.
Lambourne, D. (1995). Corydoras catfish: an aquarist’s handbook. — Blandford, London.
Magurran, A.E. (1990). The adaptive significance of schooling as an anti-predator defence in fish. — Ann. Zool. Fenn. 27: 51-66.
Makinodan, M., Rosen, K.M., Ito, S. & Corfas, G. (2012). A critical period for social experience-dependent oligodendrocyte maturation and myelination. — Science 337: 1357-1360. DOI:10.1126/science.1220845.
Mason, W. (1979). Ontogeny of social behavior. — In: Social behavior and communication (Marler, P. & Vandenbergh, J.G., eds). Springer, Boston, MA, p. 1-28. DOI:10.1007/978-1-4615-9116-0_1.
Miklósi, Á., Pongrácz, P. & Csányi, V. (1997). The ontogeny of antipredator behavior in paradise fish larvae ( Macropodus opercularis ) IV. The effect of exposure to siblings. — Dev. Psychobiol. 30: 283-291. DOI:10.1002/(sici)1098-2302(199705)30:4<283::aid-dev2>3.3.co;2-2.
Pitcher, T.J., Magurran, A.E. & Winfield, I.J. (1982). Fish in larger shoals find food faster. — Behav. Ecol. Sociobiol. 10: 149-151. DOI:10.1007/BF00300175.
Riley, R.J., Gillie, E.R., Horswill, C., Johnstone, R.A., Boogert, N.J. & Manica, A. (2019a). Coping with strangers: how familiarity and active interactions shape group coordination in Corydoras aeneus. — Roy. Soc. Open Sci. 6: 190587. DOI:10.1098/rsos.190587.
Riley, R.J., Gillie, E.R., Savage, J.L., Boogert, N.J., Manica, A. & Jungwirth, A. (2019b). The role of tactile interactions in flight responses in the Bronze Cory catfish (Corydoras aeneus). — Ethology 125: 810-820. DOI:10.1111/eth.12935.
Rodríguez-Ithurralde, D., del Puerto, G. & Fernández-Bornia, F. (2014). Morphological development of Corydoras aff. paleatus (Siluriformes, Callichthyidae) and correlation with the emergence of motor and social behaviors. — Iheringia Sér. Zool. 104: 189-199. DOI:10.1590/1678-476620141042189199.
Schausberger, P., Gratzer, M. & Strodl, M.A. (2017). Early social isolation impairs development, mate choice and grouping behaviour of predatory mites. — Anim. Behav. 127: 15-21. DOI:10.1016/j.anbehav.2017.02.024.
Sire, J.-Y. (1993). Development and fine structure of the bony scutes in Corydoras arcuatus (Siluriformes, Callichthyidae). — J. Morphol. 215: 225-244. DOI:10.1002/jmor.1052150305.
Skinner, B.F. (1966). The phylogeny and ontogeny of behavior. — Science 153: 1205-1213.
Slagsvold, T., Hansen, B.T., Johannessen, L.E. & Lifjeld, J.T. (2002). Mate choice and imprinting in birds studied by cross-fostering in the wild. — Proc. R. Soc. Lond. B Biol. Sci. 269: 1449-1455. DOI:10.1098/rspb.2002.2045.
Spence, R. & Smith, C. (2007). The role of early learning in determining shoaling preferences based on visual cues in the zebrafish, Danio rerio. — Ethology 113: 62-67. DOI:10.1111/j.1439-0310.2006.01295.x.
Tibbetts, E.A., Desjardins, E., Kou, N. & Wellman, L. (2019). Social isolation prevents the development of individual face recognition in paper wasps. — Anim. Behav. 152: 71-77. DOI:10.1016/j.anbehav.2019.04.009.
Ward, A.J.W., Hart, P.J.B. & Krause, J. (2004). The effects of habitat- and diet-based cues on association preferences in three-spined sticklebacks. — Behav. Ecol. 15: 925-929. DOI:10.1093/beheco/arh097.
Weihs, D. (1973). The mechanism of rapid starting of slender fish. — Biorheology 10: 343-350.
Wöhl, S. & Schuster, S. (2007). The predictive start of hunting archer fish: a flexible and precise motor pattern performed with the kinematics of an escape C-start. — J. Exp. Biol. 210: 311-324.
Appendix
A.1. Post-hoc test details
Below are tables with the full outputs of all post-hoc tests performed. Results are given as they were in R.
For the proportion of c-starts (Tukey post-hoc using the emmeans package).
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
For the proportion of ignoring responses (Tukey post-hoc using the emmeans package).
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011
For the proportion of tactile stimulation events where an individual responded to the stimulus by interacting tactilely with a group-mate(s).
Citation: Behaviour 157, 6 (2020) ; 10.1163/1568539X-bja10011







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}