Abstract
The larvae of black soldier fly (Hermetia illucens) (BSFL) can grow on a wide range of wastes. BSFL are known for their capacity to reduce some pathogens, while their ability to eliminate spore-forming bacteria is less documented. Five waste types were studied, three pure plant materials, and two complex waste containing animal proteins. Each diet consisted of 3 g of wheat bran, added to 7 g of humid waste. We evaluated the effects of these diets on BSFL growth, as well as the natural contamination of these wastes by two spore-forming pathogens: aerobic Bacillus cereus group bacteria, including Bacillus thuringiensis or anaerobic, Clostridioides difficile. Next, we assessed the capacity of BSFL to reduce B. thuringiensis or C. difficile inoculated at a concentration of 106 spores/g diet. After 3 and 7 days, larval weight, spore counts in larvae and frass, and the humidity and pH were assessed. Only a few wastes naturally contained B. cereus bacteria (105 CFU/g), while C. difficile was not detected. All diets, allowed healthy larval growth but larval weight significantly depended on the waste. During the challenge tests, B. thuringiensis and C. difficile were predominantly recovered as spores, and at similar concentrations across all diets and frass, while in the larvae a small fraction was recorded as vegetative B. thuringiensis. Therefore, no significant spore reduction was observed during bioconversion by BSFL. These findings underscore the importance of monitoring spore-forming pathogens in both rearing diets and final larval products to ensure the microbial safety of BSFL-based feed additives.
1 Introduction
Large quantities of organic waste are generated by human activities such as agriculture and industry. In this context, some insect species have gained interest, for their ability to thrive on low-value wastes (van Huis, 2020). One prominent example is the larva of Hermetia illucens (L.) (Diptera: Stratiomyidae), commonly known as the black soldier fly larva (BSFL). Thanks to their unique physiology, BSFL can consume and develop on a wide range of organic substrates, including agricultural by-products, supermarket leftovers, food-wastes and even manure (Wu et al., 2024). This bioconversion process transforms waste materials into high-value larval biomass which serves as a sustainable protein source to feed animals, particularly pigs, chickens and fishes (Gold et al., 2018). Beyond their role as a feed additive, the residual material left after BSFL rearing, known as frass, can also be purposed as an organic fertilizer, further contributing to circular bioeconomy practices (Lopes et al., 2022).
In Europe, regulations mandate that insects reared for feed or food production must be raised on plant-based materials. The use of animal by-products such as catering waste, manure, or other leftovers containing animal proteins is strictly prohibited under Regulation (EC) No. 1069/200. Despite this, further research is needed to investigate alternative substrates for insect rearing, supporting the development of a circular bioeconomy where waste from one process becomes a resource for another one (Slorach et al., 2019). However, microbial risk, must be carefully monitored as organic waste-streams, authorized or not, are known to host a wide variety of microorganisms including potential pathogens. In addition, the hygienic quality of the substrates used to feed insects plays a crucial role in determining the microbiological safety of the resulting products (van der Spiegel et al., 2013). For this reason, proper hygiene and robust monitoring procedures are essential to prevent the introduction of foodborne pathogens, or more broadly any contaminants harmful to humans or animals into the feed and food chain via the insect larvae. The ability of BSFL to develop on waste is influenced by the nutritional composition and stability of the substrate during rearing. Indeed, nutritional stability, in particular, is closely linked to the microbial activity within these waste streams (De Smet et al., 2018; Luparelli et al., 2022). Several studies have explored the different nutritional value of waste streams for BSFL growth (Barragan-Fonseca, 2017; Gold et al., 2020), while others have focused on food safety aspects, specifically assessing BSFL’s capacity to reduce human bacterial foodborne pathogens.
The growth of BSFL has been studied across a variety of waste streams, with a focus on the balance between an optimal growth, healthy insects, and the use of low value leftovers. BSFL requires a nutritionally balanced feed starter, several studies referring to chicken feed as a reliable option (Pinotti and Ottoboni, 2021). After the initial feeding phase, one-week-old larvae can be transferred to other food sources, but maintaining an appropriate carbon-to-protein ratio around 2:1 (50%:25% of dry matter) is critical to achieve maximum larval weight (Barragán-Fonseca et al., 2021). Likewise, the ideal substrate moisture rate should range between 40 and 80% (Nayak et al., 2024). Because organic waste streams and food leftovers differ greatly across regions, due to variation in climate and local availability, the optimal larval diet composition must be regularly adjusted. For instance, fruits like apples or apricots may require supplementation with protein and fibre, while protein-rich feeds must be balanced with additional carbon and fibre.
The microbial risk assessments of edible insect naturally contaminated with pathogens (Aguilar-Toalá et al., 2022; Brulé et al., 2024; Garofalo et al., 2019; Vandeweyer et al., 2021) or artificially inoculated (Erickson et al., 2004) have mainly focused on the yellow mealworm (Tenebrio molitor), the silkworm (Bombyx mori), the cricket (Acheta domesticus) and the grasshopper (Locusta migratoria), all of which are authorized for human consumption in Europe. In contrast, BSFL is not yet authorized for this purpose. BSFL have nevertheless a significant advantage over these species for waste bioconversion, they are able to reduce the level of several human pathogens like Escherichia coli, Staphylococcus aureus, Salmonella and Enterococcus species in their feeding substrates (Erickson et al., 2004; Gorrens et al., 2021; Lalander et al., 2013). However, reviews by Brulé et al. (2024) and Vandeweyer et al. (2021) highlight that spore-forming pathogens, such as Bacillus cereus group bacteria and Clostridium perfringens, are frequently detected in authorized processed insect products. The counts of these pathogens in the substrates remained stable or even increased, as demonstrated for B. cereus (Moyet et al., 2023). Moreover, the spores of these pathogens are highly resistant and therefore difficult to eliminate through conventional treatments used in sanitation, like heating, freezing or drying (Pakdel et al., 2023).
Most studies addressing pathogen fate have been conducted in laboratory settings, by monitoring the growth or persistence of a specific pathogen inoculated into the insect rearing substrate (Gorrens et al., 2021; Rehman et al., 2023). Such challenge-tests have mainly been performed using non-spore-forming pathogens (Erickson et al., 2004; Gorrens et al., 2021; Lalander et al., 2013). Moreover, evaluating the potential of non-authorized waste streams is essential, to assess their efficiency as a feed source for BSFL growth and to ensure microbial safety. Therefore, further research on these aspects at multiple levels is required which we partly address in the present study
Two spore-forming pathogens relevant to microbial safety were chosen: a member of B. cereus group and C. difficile. Indeed, the B. cereus group, includes several closely related and facultative aerobic species capable of causing food poisoning. Some B. cereus sensu stricto strains are of particular concern, as they have caused foodborne-poisoning outbreaks in the community due to toxins targeting the intestinal barriers (Stenfors Arnesen et al., 2008; Walia et al., 2018). Some B. cereus strains also produce cereulide, a small and highly stable toxin responsible for vomiting and more severe complications in immune compromised individuals (Yang et al., 2023). The B. cereus group also encompasses the insect pathogen Bacillus thuringiensis, which can be used as a Bio-larvicide to control agricultural insect pest (Sanahuja et al., 2011). The latter can therefore be found not only in treated fruits or vegetables but also naturally in a non-treated environment (Byun et al., 2024), moreover it has been incriminated in food poisoning (Bonis et al., 2021).
We also studied C. difficile as a strict spore-forming anaerobe and a major and orally acquired entero-pathogen of both humans and animals. Long known as a cause of healthcare-associated and post-antibiotics diarrhoea and colitis (Smits et al., 2016), C. difficile is now recognized as a One Health issue (Lim et al., 2020; Rodriguez Diaz et al., 2018; Warriner et al., 2017). It can cause infection in a wide range of animals, including wild, farm, companion animals and horses (Petry et al., 2025; Rodriguez Diaz et al., 2018). Moreover, the fecal shedding of C. difficile spores, combined with the use of manure as fertilizer, contributes to the wide contamination of the environment (Rodriguez Diaz et al., 2018; Warriner et al., 2017). C. difficile has been detected in raw vegetables, salads (Bakri et al., 2009) (Eckert et al., 2013) and potatoes (Tkalec et al., 2022) making them, as foods or feeds, potential sources of transmission (Bolton and Marcos, 2023; Borji et al., 2023; Candel-Pérez et al., 2019; Rodriguez Diaz et al., 2018; Warriner et al., 2017). C. difficile vectors include invertebrates notably insects such as houseflies, drain flies and yellow mealworms Tenebrio molitor (Burt et al., 2012; Davies et al., 2016, 2017). To date however, no study has examined the contamination of BSFL by C. difficile.
In this study, we chose five wastes: three agricultural food products (carrots, apricots and salad) and two biowastes from supermarkets (GMS) or school canteens (OMR). We assessed their ability to promote BSFL growth and the bacterial persistence of inoculated spores. As spore-forming opportunistic pathogens we focused on B. cereus/B. thuringiensis and C. difficile. We first evaluated their presence as natural contaminants in each waste. Then we performed challenge tests, to evaluate the ability of BSFL to reduce the load of spores initially spiked into each waste. This research is of novelty as it combines in a single study the evaluation of BSFL potential for both the bioconversion of several waste types and the reduction of two spore-forming, human and animal opportunistic pathogens in these wastes.
2 Material and methods
Insect rearing and assay conditions
This study utilized black soldier fly Hermetia illucens (Diptera: Stratiomyidae) larvae (BSFL) reared at BioMiMetiC (Boulbon, France; https://www.biomimetic.fr/uk/index.html). The larvae were delivered to the INRAE laboratory at Jouy-en-Josas at the juvenile stage housed in a container containing chicken feed. For growth assays and the bacterial challenge tests, larvae weighing 5 mg each were selected. Experimental conditions included incubation at 28 °C in darkness with a relative humidity (RH) of approximately 70%.

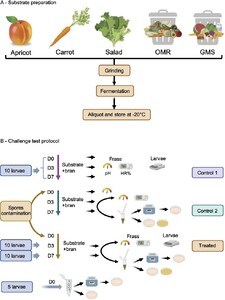
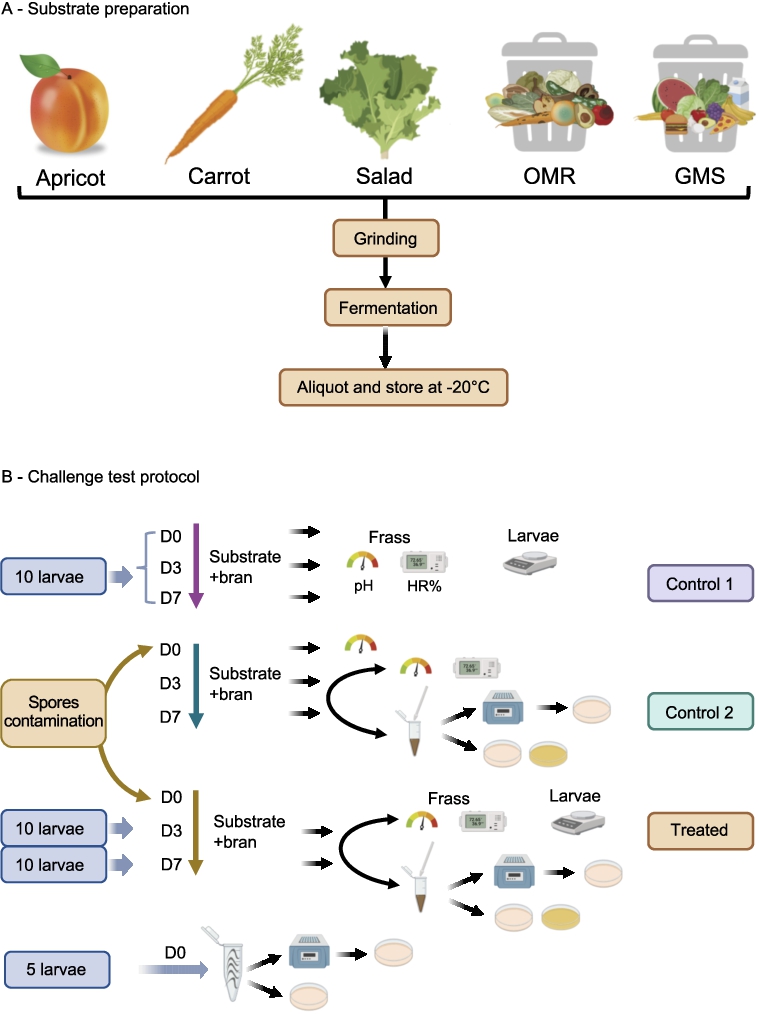
Schematic representation of waste substrate preparation (A) and experimental trial set-up and sampling method for pH, humidity, larval growth and bacterial counts in challenge tests with or without B. thuringiensis and C. difficile inoculated into waste-based diets (B).
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
Preparation of tested wastes
Five different waste-substrates, were selected as BSFL feed sources (Figure 1A). Three were “left-over agricultural production” bio-wastes: apricot, raw carrots and salad, all of which may have been exposed to soil microbes or to Bacillus thuringiensis-based bio-insecticides. They were obtained through the company BioMiMetiC. Two additional complex bio-wastes, made of mixed food ingredients and originating from supermarkets waste (GMS) and school canteens leftovers (OMR) were provided by PAPREC (Avignon, France). A standard wheat bran diet was used as a reference diet and as a supplement to the tested wastes in order to optimize nutritional balance and structural quality of the diet for BSFL rearing, as per BioMimetiC’s recommendations. To prepare the substrates, kernels were removed from apricots and all wastes were homogenized and incubated for seven days at 28 °C to allow spontaneous fermentation by their endogenous microbiota. This step was recommended in order to stabilize the substrate through partial consumption of the substrate by the natural microbiota (Quan et al., 2023). Fermentation occurred in 1-litre plastic boxes closed with a lid in which was inserted a Pasteur glass pipette to ensure gas removal. Following fermentation, 30 grams aliquots were frozen at −20 °C until used in the experimental trials (Figure 1A). To ensure a humidity level of 60-70% RH in the final diets, each substrate was adjusted to 95% humidity of which seven grams were mixed with three grams of dry (13% RH) wheat bran. Substrates after fermentation are referred to as “substrates” and those supplemented with 3 gram of wheat bran, as “diets”.
Preparation of Bacillus thuringiensis and Clostridioides difficile spores and detection methods
Bacillus thuringiensis serovar Kurstaki strain HD73 (Bt HD73-Cry), which is acrystalliferous, was chosen as a representative strain of the Bacillus cereus group. This strain has been cured for the cry-toxin gene carrying pHT73 plasmid (Wilcks et al., 1998) and transformed by electroporation with plasmid pHT315Ωerm, conferring erythromycin resistance (Arantes and Lereclus, 1991). This allows to select the strain on Erythromycin 15 μg/ml LB agar plates (LBE15). In this study referred to as BtHD73.
Spore preparation involved culturing the strain in 50 ml of HCT liquid medium, in a 500 ml Erlen flask, containing 0.3% glucose and 15 μg/ml erythromycin for 68 hours at 30 °C and 200 rpm until the sporulation was achieved; the final OD600 nm of the culture was 6.1, measured using a 1:10 dilution. After centrifugation at 6000 rpm, the supernatant was discarded and the pellet was resuspended in 5 ml of water. The suspension was then heated for 12 min at 80 °C, centrifuged for 10 min at 6000 rpm and the spore pellet was washed twice with sterile water and finally resuspended in 1.5 ml of sterile water and stored at 4 °C. The measured spore concentration was 3.3 × 108 CFU/ml.
C. difficile strain CdE-18 is non-toxigenic and had been isolated from the caecum of a necropsied foal (Petry et al., 2025). CdE-18 was grown at 37 °C under anaerobic conditions (90% N2 5% H2 5% CO2) in a rich BHIS medium (BHI supplemented with yeast extract and cysteine). Spores were prepared as previously described (Edwards and McBride, 2016). Briefly, CdE-18 was first grown over night in liquid 70:30 broth (70% BHIS and 30% sporulation medium) (Edwards and McBride, 2016). An aliquot of this culture (100 μl) was spread onto a 70:30 agar plate and grown for 10 days. Bacteria were recovered, resuspended in 500 μl of PBS (1×), and heated for 30 min at 65 °C to eliminate vegetative cells. Spores were washed and resuspended in PBS twice, and finally stored at 4 °C. The concentration of CdE-18 spores was determined to be of 9.0 × 108 CFU/ml by plating serial dilutions on either BHIS agar supplemented with taurocholate (1 g/l) or ChromID (Biomérieux, Lyon, France).
Initial detection of B. thuringiensis/B. cereus and C. difficile in substrates and larvae
The natural microbiota of fermented substrates, wheat bran and juvenile larvae were analysed for the presence of B. thuringiensis/B. cereus or Cd using a selective medium specific for each species. Substrates (2.5 g of each) were first crushed, resuspended in 25 ml of physiological water, heated or not for 10 min at 80 °C, and then serially diluted. For larval analysis, five larvae (5 mg each) were surface cleaned in water, cooled on ice and homogenized with a pestle in an Eppendorf tube of 1.5 ml containing 500 μl of cold physiological water, and finally serially diluted (1:10 dilutions) to detect B. thuringiensis/B. cereus, which were numerated on either LB-agar supplemented with E15 μg/ml or COMPASS® plates (Bacillus Plus Agar, BIOKAR, Grosseron Couëron, France).
To identify for Cd, aliquots of homogenized suspensions of larvae or substrates, were plated onto ChromID (Biomérieux, Lyon, France), but no Cd colonies could be detected in any of three 100 μl samples of these suspensions. As Cd might be present in very low amounts, we then tried to improve recovery either by spore concentration or by an enrichment step in liquid culture. For spore concentration, each sample was first transferred into a special bag, diluted in PBS and homogenized (BagMixer, Interscience) to collect the liquid phase, from which spores were concentrated by ethanol precipitation. For the enrichment step, an aliquot of crushed larvae or substrates was inoculated into a rich liquid medium: BHIS supplemented with taurocholate at 1 g/l (to promote spore germination), and d-cycloserine at 250 μg/ml and cefoxitin at 8 μg/ml (OXOID) (to counter-select many enteric bacteria), and incubated under anaerobic conditions for several days to allow vegetative growth. An aliquot of this culture was then plated onto ChromID.
Molecular identification of some endogenous species from substrates or larval microbiota
After growth of substrates or larvae aliquots on LBE15 plates under aerobic conditions or on ChromID plates under anaerobic conditions, we observed colonies whose phenotypes (morphology, colour, growth rate) differed from prototypal colonies of B. cereus or Cd strains. For non-Cd growth colony on ChromID, for each diet and for larvae, we chose a representative colony of each phenotype for molecular identification. After DNA extraction using InstaGene® Matrix (Bio-Rad) as recommended, the conserved V3-V4 region of their 16S rRNA intergenic region was amplified using the following primers: 5′-CTTTCCCTACACGACGCTCTTCCGATCTACGGRAGGCAGCAG-3′ and 5′-GGAGTTCAGACGTGTGCTCTTCCGATCTTACCAGGGTATCTAATCCT-3′, and sequenced as described in Poirier et al. (2018). A blastn search (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome) was subsequently performed to identify the corresponding species.
Relative humidity and pH of substrates
Prior to the larval feeding experiment, the moisture content of each substrate was adjusted to 95% RH by adding sterile water before wheat bran supplementation to achieve a final RH of 60-70%. The final substrates/diets precise relative humidity (%RH) was determined gravimetrically by measuring the gravimetric weight loss after drying at 28 °C until stabilization. For this purpose, one gram of diet (substrate + wheat bran) at D0 or frass (D3 and D7) was weighted from each cup and maintained at 28 °C in a half-open Petri dish for approximately 4 days until no variation of weight was observed. The pH was determined directly in ten grams of diet or frass using a calibrated pH-meter (Hanna 210) and a probe (Mettler Toledo LoT406-M6-DXK-S7/25).
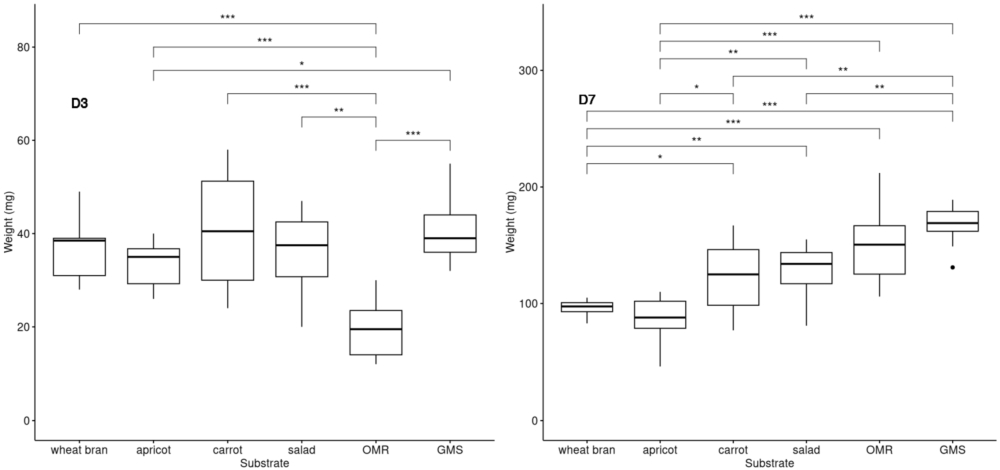
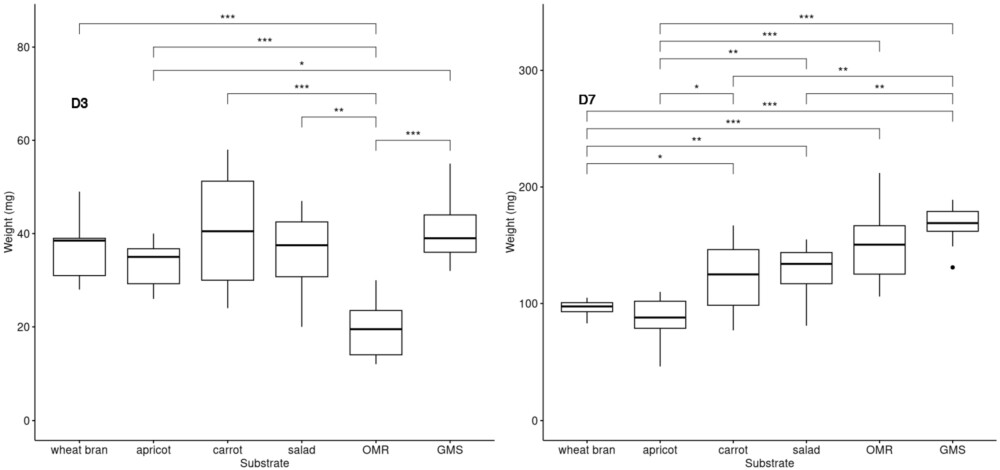
BSFL growth on different diets. The individual weights in mg of ten larvae were recorded at day 3 and day 7 for each diet (
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
Challenge test set-up for the monitoring of larval growth and of Bt HD73/CdE-18 persistence
The experimental protocol setup (Figure 1B) was designed to measure different parameters in a single run. These included the weight of BSFL after growth on the different diets and the persistence of the two pathogens (Bt HD73 and CdE-18) within the larvae, their frass and in the diets without larva (control 2). The five waste substrates combined with wheat bran (diets) without contamination were also subjected to identical experimental conditions (control 1) for comparison.
Growth and weight gain of BSFL without or with added spores
Sterile plastic cups (60 ml, Qualibact®, Labelians, France) were prepared by drilling five 2-mm-diameter holes in the lids to facilitate air circulation. Each cup was filled with 7 g of substrate (95% RH), prepared immediately after thawing, and 3 g of wheat bran (13% RH), and thoroughly mixed with a spatula to obtain a total of 10 g of diet. The initial humidity of all diets including the control (wheat bran alone) was adjusted to 70%. Ten larvae, previously fed on chicken feed until Day 0 (D0), and each weighing 5 mg, were placed in each vial containing 10 grams of one the different diets or the control. All vials were placed in an incubator set at 28 °C. To evaluate the influence of the five diets on larval growth (Figure 2), two replicate vials were prepared per diet and used for weighing the larvae: one sampled on Day 3 (D3) and the other on Day 7 (D7). To asses weight gain, larvae were recovered from the diets, rinsed with sterile water to remove any frass sticking to the teguments, dried on absorbent paper and individually weighed. The control diet consisted of wheat bran only.
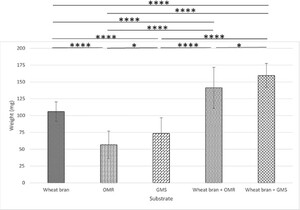
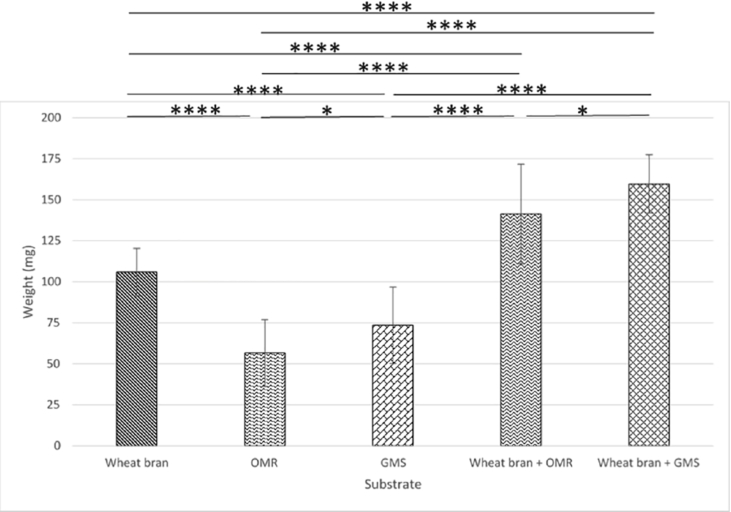
Role of wheat bran addition to OMR and GMS waste substrates. Bar plots depicting BSFL weight (in mg) after 7 days with or without 30% wheat bran, compared to a 100% wheat bran control diet. The experiment was performed in duplicate and the data presented reflect the measurements of the individual larval weight of 20 larvae (
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
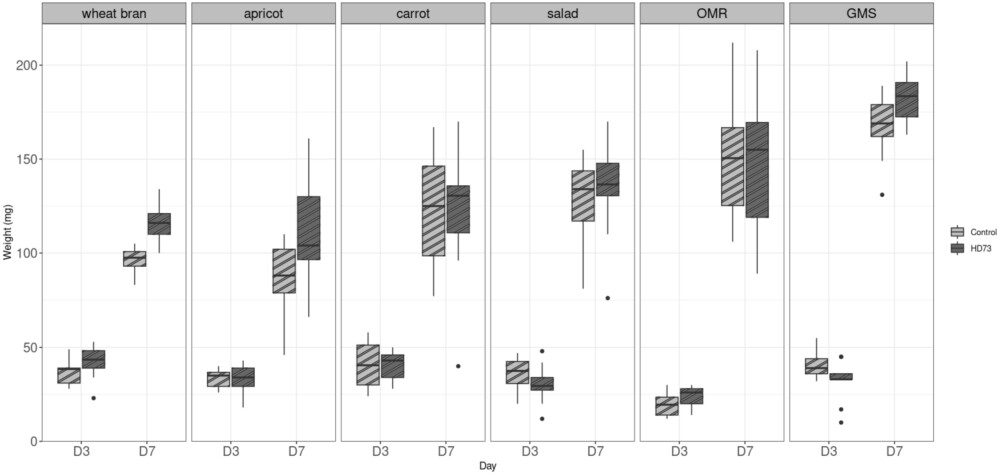
Impact on larval growth by the presence of Bt HD73. Box and whiskers plots of BSFL weights recorded from six different diets, with or without inoculation with Bt HD73, at Day 3 and Day 7. 10 larvae were weighted at each condition, (
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
Evaluation of the effect of supplementing OMR and GMS with wheat bran
Ten larvae, each weighing 5 mg, were reared for 7 days in vials containing 10 g of one of the following diets: 100% wheat bran (control), pure OMR or GMS substrates or OMR or GMS substrates supplemented with 30% wheat bran. (Figure 3)
Effects of B. thuringiensis (Bt HD73) or C. difficile (CdE-18) on larval growth
Ten 5-days-old BSFL, each weighing 5 mg, were placed at Day 0 (D0) on vials containing 10 g of food (7 g of a given substrate + 3 g of wheat bran) contaminated or not with spores (Figures 1 and 4). Spore preparations (see below) were mixed thoroughly into the diets to ensure even distribution and achieve target final spore concentration of about 106 CFU/g of any diet. The exact final concentration of the inoculum, for each experiment, was verified by plating on selective media. At Day 0 (D0) the Bt HD73 or CdE-18 spore counts were consistently close to the targeted level, at approximately (3-5) × 106 CFU/g in diets. Two vials per condition were incubated at 28 °C and larvae were weighed on Day 3 (vial 1) and Day 7 (vial 2) as for the vials without spores Wheat bran alone served as the control.
Inoculation of BtHD73 and CdE-18 spores into BSFL diets for challenge tests
For the challenge tests, the initial suspensions of Bt HD73 (3.3 × 108 CFU/ml) or CdE-18 spores (9.0 × 108 CFU/ml) were diluted 1:10 in physiological water and an aliquot of this dilution (300 μl) was added to 10 g of each diet and thoroughly mixed with a spatula. Bt HD73 spores were inoculated into all five diets at a final concentration of 3.5 × 106 CFU/g, exceeding the initial contamination levels (approx. 105 CFU/g) of B. cereus group bacteria initially found in fermented GMS and OMR substrates (see results below). CdE-18 spores were added to carrot and OMR diets at a similar final concentration (measured to be of 2.8 × 106 CFU/g carrots).
Monitoring of BtHD73 and CdE-18 in substrates, frass and larvae
The total viable counts (vegetative cells + spores) and spore counts were determined before and after heat treatment (10 min at 80 °C) of the diets, frass or crushed larvae samples. Bacterial counts (CFUs) were determined through serial 10-fold dilution and plating. For diet and frass, 500 mg (in triplicate) of each diet/frass sample was mixed with 500 μl of physiological water. For larvae, groups of three larvae were cooled on ice, surface sterilized with 70% ethanol, rinsed with sterile water. Larvae were then cut in small pieces with fine scissors, placed into 1.5-ml Eppendorf tubes containing either 500 μl (D3) or 800 μl (D7) cold physiological water and homogenized with a plastic pestle. The final volumes and the weight of the larvae were taking into account to determine the dilution rate and CFU per gram. Three replicates were performed for each sample type (diet, frass) and each group of three larvae (per diet) at each time point.
Each suspension (from diets, frass and larvae) was analysed in two ways: first, serial dilutions of the unheated suspensions, were plated to determine total CFU counts (spores + vegetative bacteria) then the same tubes were heat-treated (80 °C for 10 min) to quantify spores only. The serial dilutions were plated on different selective media: LB E15 and COMPASS® Bacillus Plus Agar (BIOKAR) for Bt HD73 and ChromID for CdE-18. COMPASS® plates enabled detection of B. cereus group bacteria, which appeared as blue/green colonies with a specific morphology.
Data analyses
Larval weight gain of BSFL across different diets was analysed using the Kruskal-Wallis test to assess overall differences and evaluate whether the larval weight from one diet was different from the weight in another diet. Diets were tested with either 10 larvae (as shown in Figures 2 and S1) or 20 larvae (Figure 3). Weights were recorded at D0, D3, and D7. For comparisons between diets, the Mann-Whitney pairwise test was employed. Graphical data visualizations were generated using RStudio software with the ggplot2, ggpubr and ggpatern libraries. Data are represented as box and whiskers plots, showing the minimum, first quartile, median, third quartile, and maximum larval weight values (in mg). To monitor Bt HD73 or CdE-18 contamination levels in diets, frass or larvae over time, three replicates of each sample type (three replicates of 500 mg of diet or frass and three replicates of three larvae (
3 Results
Measurement of the substrate pH and humidity variation during the experiment
BSFL can tolerate high relative humidity (RH) conditions, with optimal development occurring between 60 and 70% RH (Barragan-Fonseca, 2017). In this study, the initial RH of the five diets (7 g waste and 3 g wheat bran) and of the control substrate (wheat bran alone) was adjusted to between 57 and 67 RH% at day 0 (D0). The rearing temperature was maintained at 28 ± 2 °C. Throughout the experiment, pH and RH% were measured on days 0, 3 and 7 for all substrates.
pH evolution over time
At D0, all substrates, except for wheat-bran (pH 5.9) and salad (pH 5.2) had initial low pH levels. Apricot and OMR exhibited the most acidic conditions with initial pH values of 4.2 and 4.5, respectively. By day 7, in the presence of the larvae, pH values increased substantially exceeding 7.0 and up to 9.1 for wheat bran at day 7 in all the diets/frass, indicating that the larval activity had an alkalizing effect across all substrates (Table S1). In contrast, substrates maintained without larvae displayed minimal pH increases, highlighting the role of larval metabolism in modifying the chemical environment (Table S1).
Humidity monitoring
Moisture content of the diets and frass was measured by incubating three 500 mg samples of each diet (or frass when larvae were present) at D0, D3 and D7 and at 28 °C with circulating air until no further weight variation was observed. The final dry matter content was used to calculate the diets/frass RH. The results showed that the moisture content remained relative stable throughout the experiment, consistently above 56% RH in all cases. In some diets, RH slightly increased to as high as 75-77% by day 7, particularly in diets with larvae (Table S2) suggesting that this increase may be attributed to larval activity and substrate mixing which may help to maintain or even slightly elevate moisture levels in the tested conditions (cups with caps/small holes). In contrast, diets without larvae maintained constant RH throughout the trial.
Impact of the five waste diets on BSFL growth
This experiment aimed to evaluate the influence of the five substrates on larval growth by measuring the weight of BSFL reared on these substrates. Larval weight served as an indicator of development and substrate suitability for BSFL rearing. Three substrates were composed of unsold fruit and vegetables (apricots, carrots and salad), while the other two consisted of food leftovers from school canteens (OMR) and supermarket (GMS). Day 0 (D0) marked the start of the experiment when the larvae were placed on diets. To minimize disturbance, the vials and their content were not opened or manipulated before to be analysed. The results showed that the larvae grew well regardless of the diet, as their initial weight of 5 mg larvae increased significantly over the course of the experiment. At day 3 (D3) the average medium weight of a larva was from 20-45 mg and between 101-166 mg at day 7, depending on the diet. Statistical differences (p values) observed between them are shown in Figure 2. Larvae reared on diets containing animal protein (OMR and GMS substrates) exhibited significantly higher weights by D7 compared to those on other diets, especially with respect to apricot.
Impact of adding wheat bran to OMR and GMS substrates on BSFL growth
This experiment evaluated the effect of supplementing OMR and GMS waste-stream substrates with 30% wheat bran on BSFL growth. Larvae were reared for 7 days on the five diets compositions: control diets of 7 g pure wheat bran, OMR, GMS (adjusted to ≈70% humidity) or OMR (95 RH%) + 3 g wheat bran, or GMS (95 RH%) + 3 g wheat bran. The results presented in Figure 3 indicate that larvae fed with pure OMR or GMS substrates showed significantly reduced growth performance compared to the wheat bran control. Specifically, larval biomass was reduced by 47 and 31% for OMR and GMS, respectively. Among all tested diets, OMR yielded the poorest results in terms of larval biomass production. On the other hand, adding three grams of wheat bran to the OMR and GMS substrates significantly improved larval weight. Larvae reared on the supplemented diets achieved a biomass increase of 40% (OMR + wheat bran) and 58% (GMS + wheat bran) compared to the wheat bran control. These findings align with the outcomes of the growth assays (Figure 2), reinforcing the beneficial impact of wheat bran supplementation in enhancing larval growth.
Impact of diets spiked with Bacillus thuringiensis (BtHD73) on BSFL growth
Organic waste streams, including the authorized ones, can potentially harbour a wide range of harmful microorganisms. To investigate the behaviour and effects of a spore-forming bacterium during bioconversion of contaminated substrates by BSFL, we inoculated the five diets with B. thuringiensis. Larvae fed on those diets were monitored to determine whether the pathogen impacted BSFL growth as compared to the uncontaminated control. An overview of these experimental conditions is provided in Figure 1B. For this experiment, and for each diet, two different rearing conditions were tested: diet with larvae but without pathogen, and (ii) diet inoculated with the pathogen and with larvae.
The results, presented in Figure 4, indicate that BSFL weight gain was not affected by the presence of B. thuringiensis. This suggests that the presence of Bt HD73 did not have a significant negative or positive effect on BSFL growth characteristics. However, larval weights at Day 7 (D7) were significantly higher (
Search for B. cereus and C. difficile in BSFL intestinal microbiota or in substrates
As a control before the challenge tests with BtHD73 and CdE-18 in BSFL, we examined whether the microbiota of juvenile BSFL and waste substrates contained B. cereus group bacteria or Cd or other species able to grow under the same conditions as those chosen for the challenge tests: (i) on LBE15 or COMPASS® under aerobiosis for B. thuringiensis and (ii) on ChromID under anaerobiosis for C. difficile.
The natural presence of B. cereus group bacteria in juvenile BSFL was investigated by spreading crushed larvae on LBE15 agar plates. The results revealed the presence of endogenous aerobic, erythromycin-resistant bacteria at a concentration of 108 CFU/g. After heat treatment, the bacterial concentration dropped to 104 CFU/g, indicating the presence of both erythromycin and heat-resistant colonies, which might be spore-formers. However, none of them were B. cereus/B. thuringiensis colonies based on their colony morphology and their lack of growth or coloration on the selective COMPASS® Bacillus Plus Agar medium. To the opposite none of the five tested fermented substrates (apricot, carrot, salad, OMR or GMS) contained erythromycin resistant colonies, no growth on LB E15. When plated on the selective COMPASS® medium, Bt/Bc-like colonies were detected at a concentration of 105 CFU/g substrate for OMR and GMS but not recorded in the other substrates even in 100 μl of a 10−1 dilution of the suspension on heated sample. In addition, other small colonies with bluish centers and a with a morphology different from BtHD73 were detected on COMPASS® plates No further characterization of non-B. cereus colonies was performed.
We also tested whether endogenous C. difficile or other species with similar growth requirements could belong to BSFL or diet microbiota. No C. difficile colonies, which are characterized by an irregular shape, a tar-like smell and in most cases, a black color on ChromID, could be identified in any case, even when 100 μl of each concentrated sample (by ethanol precipitation, see materials and methods) were directly plated on ChromID. This indicated that Cd was not present or undetectable (10−1-100 CFU/g diet). Yet, black colonies of regular shapes and of two different sizes in the case of OMR could be detected at a level of approx. 100-101 CFU/g OMR or GMS and of approx. 104 CFU/g salad. After having sequenced the conserved V3-V4 region of their chromosomal 16S rRNA intergenic region, they were identified as: (i) Clostridium sartagoforme in larvae, (ii) Clostridium butyricum in salads, GMS and OMR samples, and (iii) Paenibacillus spp in OMR samples (small colonies of delayed growth), these species are spore-forming anaerobes, and specifically strict or facultative anaerobes for the first two and for the latter, respectively.
Monitoring and behavior of BtHD73 in diets, larvae and frass following spiking of the diets
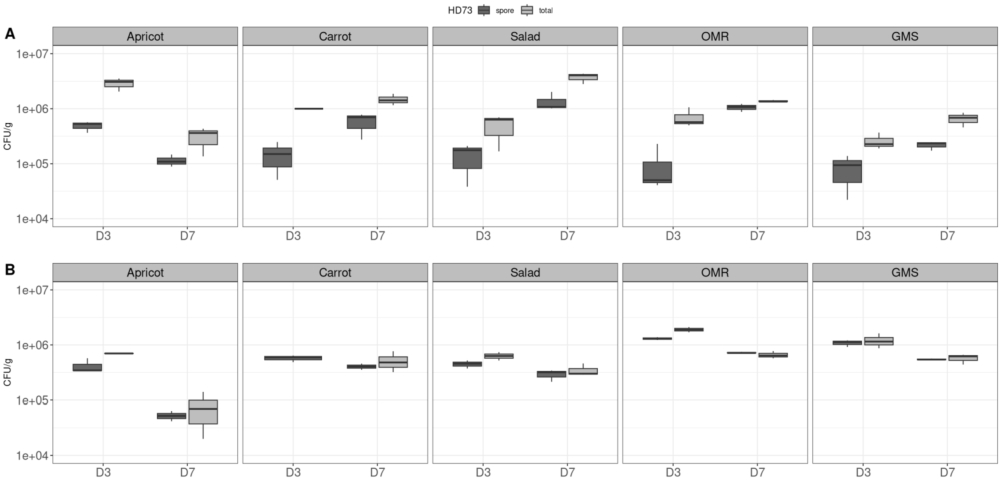
Next, we investigated the persistence and behaviour of BtHD73 spores, in the five diets during rearing and under a high contamination scenario. Our aim was to determine whether the Bt HD73 strain introduced at D0 persisted as spores (heat-resistant forms) or transitioned to a metabolically active vegetative cells after 3 and 7 days. This assessment was carried out for diets both in the absence of larvae and in vials containing larvae. In the latter case, we analysed the diet, the larvae, and the frass (a mixture of leftover substrate, exuviae, and faeces). The results of these measurements are presented in Figure S2. Although having only three values per condition (each diet) the statistical tests were performed but no significant differences between spore and total count was found in any of the diets during the experiments in both larvae, diet, and frass, because of the low number of samples per diet. Meanwhile, when data from the five diets were grouped, meaning 15 values per diet/ per day (D3 or D7), significant differences (pairwise Wilcoxon rank test) were found for some. In the case of larvae, p-values of 0.0023 (significant,
Dynamics of BtHD73 in larvae and in the frass on D3 and D7. Black boxes, total HD73 (vegetative cells+ spores); grey boxes, HD73 spores For each diet and time point, three replicates of three larvae (A) or three replicates of frass (B) were performed. The y-axis represents CFU/g diet on a logarithmic scale ranging from 4 log to 7 log. Box and whisker plots, presenting the minimum, first quartile, the median, the third quartile, and the maximum values (CFU/g), are shown. With only three values by condition (diet/day) the statistical Wilcoxon rank (Mann-Whitney test, showed no significant differences between total counts and spores only.
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
Growth of BSFL and monitoring of CdE-18 in larvae and diets
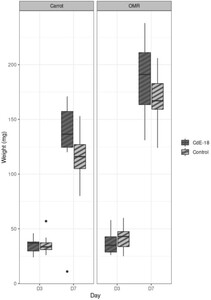
We monitored the growth of juvenile larvae on two diets, carrots or OMR, after their inoculation or not by CdE-18 spores, at a level similar to that used for B. thuringiensis spores (approx. 106 CFU/g). There was no significant difference in BSFL final weight after growth on any diet, irrespective of whether it had initially been inoculated or not by CdE-18 spores, and at any final time point (D3 and D7) (Figure 6). We, then inoculated carrots with CdE-18 spores and studied their fate. We measured CdE-18 levels at the two time points in the larvae and frass for the assays of BSFL reared on carrots, and in the carrot diet for the control without larvae. The CdE-18 level was of the same order of magnitude at D3 and D7 in the inoculated carrots (substrate without larvae) (Figure S4A) and in the frass (Figure S4B). In larvae reared on carrots, the CdE-18 level did not decrease as a function of time. A similar result was obtained with BtHD73 spores in all diets except apricot (Figure 5A).
BSFL weight after growth on carrot or OMR diets inoculated or not by C. difficile. Carrot (left) or OMR (right) diets, which had initially been inoculated (dark grey) or not (light grey) by CdE-18 spores at 106 CFU/gram diet, were used to rear BSFL as previously described in Figure 4. The weight of BSFL was measured at two time points, day 3 (D3) and day 7 (D7). Whiskers plots, presenting the minimum, first quartile, the median, the third quartile, and the maximum values in mg are shown. Pairwise comparison using the Wilcoxon rank test (Mann-Whitney test) indicated that there were no statistically significant differences between diet with or without CdE-18.
Citation: Journal of Insects as Food and Feed 12, 6 (2026) ; 10.1163/23524588-bja10317
4 Discussion
The utilization of black solder fly larvae (BSFL) for bioconversion of organic wastes is a promising example of a sustainable and circular approach in the feed and food production chain. However, organic wastes often contain a wide range of diverse microbes (bacteria, viruses and parasites), some of which may be potentially pathogenic to humans or insects. These contaminants can impact larval growth, health and the microbial safety of the final larvae-derived products. This study evaluated BSFL’s efficiency to convert low-value waste streams into biomass and examined the fate of spore-forming pathogenic bacteria during the rearing process in these substrates. In doing so, this study addressed a knowledge gap regarding BSFL’s capacity to reduce endospore-forming bacterial pathogens and whether the waste type could influence on that. Our study utilized five different waste substrates including three plant-based products and two animal-protein-containing substrates and a control wheat bran standard diet.
Suitability of waste-based diets for BSFL growth
We first determined whether the five pre-fermented substrates were suitable for BSLF growth. BSFL reared for seven days on diets composed of these waste-substrates, each supplemented with wheat bran, exhibited no mortality, indicating that none of the substrates contained harmful contaminants and that they supported larval growth and development. However, larval weight differed depending on the substrate nutritional composition (Figure 2). High-protein diets like OMR and GMS significantly improved larval growth compared to apricot (
However, it should be noted that the nutritional composition of animal protein-based substrates can also vary significantly seasonally or from batch-to-batch, which may impact both nutrient content and larval growth. Although we did not analyse larval nutrient value composition and chemical contamination in this study, future work should investigate these parameters to understand their impact on the nutrient quality of the larvae as feed. Regarding the initial pH of the diets, we observed no significant differences in final larval weights between acidic (pH < 5) or nearly neutral (pH > 6) substrates, indicating BSFL’s tolerance for acidic substrates, are consistent with prior reports showing growth at a pH as low as 4 (Ma et al., 2018). Of note, in a small-scale growth assay (data not shown), young 5-day old larvae placed on frass recovered at Day 7 exhibited minimal growth, suggesting that the frass at Day7 had low residual nutritional value. This observation supports the conclusion that our experimental setup (10 BSFL individuals/10 g humid feed diet) was close to optimal and comparable to what is found in large rearing batches on fermented and stabilized waste, as communicated by BioMimetic, a commercial BSFL producer company, who advised on the setup.
Natural contamination of substrates or larvae by B. cereus and C. difficile
B. cereus/B. thuringiensis was not detected in juvenile larvae, while its levels reached 105 CFU/g in the plant-based substrates and in GMS and OMR substrates. The initial pH was low, due to the pre-fermentation, which has also modified the microbial composition as compared to the initial fresh non fermented waste. Fermentation is commonly used for food preservation, as it inhibits microbial growth of several pathogens or of food-alteration microbes. Bacillus cereus can tolerate environments with a pH as low as 4.6 (Le Marc et al., 2021). The pH increased over time in the frass, likely due to ammonia production (Parodi et al., 2020), potentially altering microbial dynamics and larval interactions with the pathogens in later growth stages. Further studies are also needed to examine factors such as humidity, which could affect BSFL growth and their interactions with pathogens (Nayak et al., 2024). Cd was not detected in the batch of carrots studied here. Indeed, in systematic studies performed in Slovenia and in Europe, and dealing with a similar root vegetable, reported 28 and 22% prevalence for Cd in the tested batches (Tkalec et al., 2022). Similarly, Cd was not detected in juvenile BSFL. Although Cd had previously been found in several insects such as houseflies, drain flies and even edible yellow mealworms Tenebrio molitor (Burt et al., 2012; Davies et al., 2016, 2017), Cd has not yet been detected in insect-based food/feed products. In contrast, B. cereus was the most prevalent foodborne pathogen in these products (present in 58% of insect-based feed products), followed by C. perfringens (16%) and Salmonella enterica (1%) (Frentzel et al., 2022). Of note, in this study, we could detect two strict anaerobic species: C. sartagoforme in BSFL and C. butyricum in plant-derived substrates and in animal wastes, some of which can contain strains pathogenic for humans.
Impact of BtHD73 and CdE-18 on BSFL Growth
By rearing BSFL on five diets inoculated or not with spores of BtHD73, we found no significant effect on larval viability (not shown) or larval growth and final weight (Figure 4). Similarly, CdE-18 inoculation of carrots or OMR diets showed no significant impact (Figure 6). Previous studies have reported that Bacillus subtilis supplementation to chicken manure may improve BSFL growth, possibly due to enzymatic degradation enhancing substrate digestibility (Xiao et al., 2018), but such effects were not observed in this study with BtHD73 or CdE-18 supplementation.
Persistence of Bt HD73 and CdE-18, spores during BSFL rearing
To evaluate the ability of BSFL’s to reduce pathogen spore loads, diets were inoculated with BtHD73 and CdE-18 spores. No significant reduction in spore concentrations of BtHD73 was observed, regardless of the presence of larvae (Figure 5B, Figure S2), although the vegetative form was found inside the larvae but not or less in frass from all diets; indicating that there may be some spore germination at least in the gut. These findings align with prior studies showing that spore-forming bacteria persist in processed insect products (Brulé et al., 2024; Vandeweyer et al., 2021). The low pH of some substrates may have impeded spore germination and bacterial growth (Vandeweyer et al., 2020). Likewise, BSFL was unable to reduce the load of Cd spores in carrots. Similar results were previously been reported in Musca domestica larvae, which were able to acquire and internalize C. difficile spores during feeding and to retain them for a few days without reducing them (Davies et al., 2017). Therefore, neither M. domestica larvae nor BSFL, both dipteran larvae, were able to eliminate Cd spores under continuous exposure via spiked diet. These results are consistent with other studies, which reported that Bacillus or other Clostridium spores frequently persist in both processed and unprocessed edible insects (Brulé et al., 2024; Vandeweyer et al., 2021). Our findings also align with a previous report, indicating that B. cereus was not reduced in the presence of BSFL (Moyet et al., 2023). In that study, mashed and sterilized potatoes were inoculated with an 18-h culture of B. cereus, although it was not specified whether the culture used for spiking contained spores or only vegetative cells or both. In our study, total CFU counts in the larvae, showed greater variability, particularly at day three, where they exceeded the spore counts, suggesting the presence of vegetative cells, which was significantly shown following analysis of grouped data from all diet (Figure S4). This was especially apparent in BSFL reared on apricot and carrot substrates. However, it remains unclear whether the BtHD73 spores were unable to germinate in the diet or whether any resulting vegetative forms were readily eliminated by the acidic substrate or by the acidic environment of the larval gut (Bonelli et al., 2019), explaining why mainly spores were found in the frass. Importantly, no increase in the total CFU or spore counts was recorded, supporting the conclusion that no major germination nor proliferation occurred under the tested conditions. Additionally, in a supplementary inoculation experiment using the vegetative form of BtHD73 in apricot and GMS, no bacterial growth was observed in either the inoculated diets or the larvae that fed on them (data not shown). This suggests that the vegetative forms of the BtHD73 strain were unable to survive or multiply within these substrates themselves. These findings are consistent with a recent study reporting that when two B. cereus strains were inoculated into wheat bran diet as either vegetative or spores, only the spores survived following BSFL bioconversion (van Kessel et al., 2024). Further research is needed to better understand the ecology of these bacteria in the BSFL rearing environment, and more specifically the factors that inhibit spore germination and those reducing the survival of vegetative cells in these substrates, in the presence or absence of larvae.
Implications for Food Safety and Regulation
Current European regulations permit only the utilization of plant-based diets for BSFL rearing, excluding animal-protein-containing wastes. However, our results demonstrate that all tested substrates, including those not authorized for BSFL rearing, are suitable for larval development. Nevertheless, the spores of the two bacteria we added to the diets were also found in the larvae, indicating that contamination occurred, at least when they were collected also directly from contaminated feed. As a result, these spores may remain viable in the final product after insect processing, posing potential food safety concerns (Fasolato et al., 2018; Vandeweyer et al., 2020). Future research should prioritize developing innovative methods to inactivate spore-forming pathogens and exploring new strategies for controlling spore-forming pathogens contamination during BSFL production. Likewise, it is important to evaluate spore persistence in the larvae after removal from contaminated substrates, as has been done for the bacteria Serratia in Tenebrio molitor (Dupriez et al., 2022). Our findings can also guide regulatory decision-making on the nutritional benefits of animal-derived substrates, and helps determining which rearing, processing, and decontamination methods should be employed for the safe large-scale production of BSFL.
5 Conclusion
This study underscores the suitability of diverse organic waste streams for BSFL rearing and confirms the persistence of spore-forming pathogens throughout the entomo-conversion process. While BSFL did not significantly reduce pathogen spore loads, they effectively converted waste into biomass without adverse effects on larval growth. Moving forward, regulatory and technological developments are needed to improve microbiological food safety and facilitate a broader adoption of BSFL in sustainable waste management and feed bioconversion systems.
Corresponding author; e-mail: christina.nielsen-leroux@inrae.fr
Acknowledgements
This work was supported by the French Institut National de la Recherche pour l’Agriculture, l’Alimentation et l’Environnement (INRAE) and was carried out in the framework of the INRAE-Metaprogram BETTER project, called Fly4Waste (2021-2024), in which the French company BioMiMetiC, a producer of BSFL, provided BSFL and guidance for larval growth conditions. The authors thank Tibor Bonhomme for his contribution to the project during his internship.
References
Aguilar-Toalá, J.E., Cruz-Monterrosa, R.G. and Liceaga, A.M., 2022. Beyond human nutrition of edible insects: health benefits and safety aspects. Insects 13: 1007. https://doi.org/10.3390/insects13111007
Arantes, O. and Lereclus, D., 1991. Construction of cloning vectors for Bacillus thuringiensis. Gene 108: 115-119. https://doi.org/10.1016/0378-1119(91)90495-W
Bakri, M.M., Brown, D.J., Butcher, J.P. and Sutherland, A.D., 2009. Clostridium difficile in ready-to-eat salads, Scotland. Emerging Infectious Diseases 15: 817-818. https://doi.org/10.3201/eid1505.081186
Barragán-Fonseca, K., Dicke, M., Gort, G. and van Loon, J., 2021. Nutritional plasticity of the black soldier fly (Hermetia illucens) in response to artificial diets varying in protein and carbohydrate concentrations. Journal of Insects as Food and Feed 7: 51-61. https://doi.org/10.3920/JIFF2020.0034
Barragan-Fonseca, K.B., Dicke, M. and van Loon, J.J.A., 2017. Nutritional Value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed – a review. Journal of Insects as Food and Feed 3: 105-120. https://doi.org/10.3920/JIFF2016.0055
Bolton, D. and Marcos, P., 2023. The environment, farm animals and foods as sources of infection in humans. Foods 12: 1094. https://doi.org/10.3390/foods12051094
Bonelli, M., Bruno, D., Caccia, S., Sgambetterra, G., Cappellozza, S., Jucker, C., Tettamanti, G. and Casartelli, M., 2019. Structural and functional characterization of larval midgut. Frontiers in Physiology 10: 204. https://doi.org/10.3389/fphys.2019.00204
Bonis, M., Felten, A., Pairaud, S., Dijoux, A., Maladen, V., Mallet, L., Radomski, N., Duboisset, A., Arar, C., Sarda, X., Vial, G., Mistou, M.Y., Firmesse, O., Hennekinne, J.A. and Herbin, S., 2021. Comparative phenotypic, genotypic and genomic analyses of associated with foodborne outbreaks in France (Publication with Expression of Concern. See vol. 18, 2023). PLoS ONE 16: e0246885. https://doi.org/10.1371/journal.pone.0246885
Borji, S., Kadivarian, S., Dashtbin, S., Kooti, S., Abiri, R., Motamedi, H., Moradi, J., Rostamian, M. and Alvandi, A., 2023. Global prevalence of of Clostridioides difficile in 17,148 food samples from 2009 to 2019: a systematic review and meta-analysis. Journal of Health Population and Nutrition 42: 36. https://doi.org/10.1186/s41043-023-00369-3
Brulé, L., Misery, B., Baudouin, G., Yan, X., Guidou, C., Trespeuch, C., Foltyn, C., Anthoine, V., Moriceau, N., Federighi, M. and Boué, G., 2024. Evaluation of the microbial quality of hermetia illucens larvae for animal feed and human consumption: study of different type of rearing substrates. Foods 13: 1587. https://doi.org/10.3390/foods13101587
Burt, S.A., Siemeling, L., Kuijper, E.J. and Lipman, L.J.A., 2012. Vermin on pig farms are vectors for PCR ribotypes 078 and 045. Veterinary Microbiology 160: 256-258. https://doi.org/10.1016/j.vetmic.2012.05.014
Byun, K.H., Kang, M., Koo, M.S., Lim, M.C., Ok, G.S. and Kim, H.J., 2024. Potential risk of biofilm-forming Bacillus cereus group in fresh-cut lettuce production chain. Food Research International 191: 114692. https://doi.org/10.1016/j.foodres.2024.114692
Candel-Pérez, C., Ros-Berruezo, G. and Martı́nez-Gracı́a, C., 2019. A review of occurrence through the food chain. Food Microbiology 77: 118-129. https://doi.org/10.1016/j.fm.2018.08.012
Davies, M.P., Anderson, M. and Hilton, A.C., 2016. The housefly Musca domestica as a mechanical vector of Clostridium difficile. Journal of Hospital Infection 94: 263-267. https://doi.org/10.1016/j.jhin.2016.08.023
Davies, M.P., Anderson, M. and Hilton, A.C., 2017. Acquisition and retention of of Clostridium difficile by Musca domestica larvae and pupae during metamorphosis. Journal of Hospital Infection 95: 410-414. https://doi.org/10.1016/j.jhin.2017.01.010
De Smet, J., Wynants, E., Cos, P. and Van Campenhout, L., 2018. Microbial community dynamics during rearing of black soldier fly larvae (Hermetia illucens) and impact on exploitation potential. Applied and Environmental Microbiology 84: e02722-17. https://doi.org/10.1128/aem.02722-17
Dupriez, F., Rejasse, A., Rios, A., Lefebvre, T. and Nielsen-LeRoux, C., 2022. Impact and persistence of in larvae and feed under optimal and stressed mass rearing conditions. Insects 13: 458. https://doi.org/10.3390/insects13050458
Eckert, C., Burghoffer, B. and Barbut, F., 2013. Contamination of ready-to-eat raw vegetables with Clostridium difficile in France. Journal of Medical Microbiology 62: 1435-1438. https://doi.org/10.1099/jmm.0.056358-0
Edwards, A.N. and McBride, S.M., 2016. Isolating and purifying Clostridium difficile spores. Methods in Molecular Biology 1476: 117-128. https://doi.org/10.1007/978-1-4939-6361-4_9
Eggink, I.K.M., Donoso, I.G. and Dalsgaard, J., 2023. Optimal dietary protein to carbohydrate ratio for black soldier fly (Hermetia illucens) larvae. Journal of Insects as Food and Feed 9: 789-798. https://doi.org/10.3920/JIFF2022.0102
Erickson, M.C., Islam, M., Sheppard, C., Liao, J. and Doyle, M.P., 2004. Reduction of O157:H7 and Salmonella enterica serovar enteritidis in chicken manure by larvae of the black soldier fly. Journal of Food Protection 67: 685-690. https://doi.org/10.4315/0362-028x-67.4.685
Fasolato, L., Cardazzo, B., Carraro, L., Fontana, F., Novelli, E. and Balzan, S., 2018. Edible processed insects from e-commerce: food safety with a focus on the cereus group. Food Microbiology 76: 296-303. https://doi.org/10.1016/j.fm.2018.06.008
Frentzel, H., Kelner-Burgos, Y., Fischer, J., Heise, J., Göhler, A. and Wichmann-Schauer, H., 2022. Occurrence of selected bacterial pathogens in insect-based food products and in-depth characterisation of detected group isolates. International Journal of Food Microbiology 379: 109860. https://doi.org/10.1016/j.ijfoodmicro.2022.109860
Garofalo, C., Milanovic, V., Cardinali, F., Aquilanti, L., Clementi, F. and Osimani, A., 2019. Current knowledge on the microbiota of edible insects intended for human consumption: a state-of-the-art review. Food Research International 125: 108527. https://doi.org/10.1016/j.foodres.2019.108527
Gold, M., Tomberlin, J.K., Diener, S., Zurbrügg, C. and Mathys, A., 2018. Decomposition of biowaste macronutrients, microbes, and chemicals in black soldier fly larval treatment: A review. Waste Management 82: 302-318. https://doi.org/10.1016/j.wasman.2018.10.022
Gold, M., von Allmen, F., Zurbrügg, C., Zhang, J.B. and Mathys, A., 2020. Identification of bacteria in two food waste black soldier fly larvae rearing residues. Frontiers in Microbiology 11: 582867. https://doi.org/10.3389/fmicb.2020.582867
Gorrens, E., Van Looveren, N., Van Moll, L., Vandeweyer, D., Lachi, D., De Smet, J. and Van Campenhout, L., 2021. Staphylococcus aureus in substrates for black soldier fly larvae (Hermetia illucens) and its dynamics during rearing. Microbiology Spectrum 9: e02183-21. https://doi.org/10.1128/Spectrum.02183-21
Lalander, C., Diener, S., Magri, M.E., Zurbrügg, C., Lindström, A. and Vinnerås, B., 2013. Faecal sludge management with the larvae of the black soldier fly (Hermetia illucens) from a hygiene aspect. Science of the Total Environment 458: 312-318. https://doi.org/10.1016/j.scitotenv.2013.04.033
Lalander, C., Diener, S., Zurbrügg, C. and Vinnerås, B., 2019. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). Journal of Cleaner Production 208: 211-219. https://doi.org/10.1016/j.jclepro.2018.10.017
Le Marc, Y., Baert, L., Buss da Silva, N., Postollec, F., Huchet, V., Baranyi, J. and Ellouze, M., 2021. The effect of pH on the growth rate of Bacillus cereus sensu lato: Quantifying strain variability and modelling the combined effects of temperature and pH. International Journal of Food Microbiology 360: 109420. https://doi.org/10.1016/j.ijfoodmicro.2021.109420
Lim, S.C., Knight, D.R. and Riley, T.V., 2020. Clostridium difficile and one health. Clinical Microbiology and Infection 26: 857-863. https://doi.org/10.1016/j.cmi.2019.https://doi.org/10.023
Lopes, I.G., Yong, J.W. and Lalander, C., 2022. Frass derived from black soldier fly larvae treatment of biodegradable wastes. A critical review and future perspectives. Waste Manag 142: 65-76. https://doi.org/10.1016/j.wasman.2022.02.007
Luparelli, A.V., Saadoun, J.H., Lolli, V., Lazzi, C., Sforza, S. and Caligiani, A., 2022. Dynamic changes in molecular composition of black soldier fly prepupae and derived biomasses with microbial fermentation. Food Chem X 14: 100327. https://doi.org/10.1016/j.fochx.2022.100327
Ma, J., Lei, Y., Rehman, K.u., Yu, Z., Zhang, J., Li, W., Li, Q., Tomberlin, J.K. and Zheng, L., 2018. Dynamic Effects of Initial pH of substrate on biological growth and metamorphosis of black soldier fly (Diptera: Stratiomyidae). Environmental Entomology 47: 159-165. https://doi.org/10.1093/ee/nvx186
Moyet, M., Morrill, H., Espinal, D.L., Bernard, E. and Alyokhin, A., 2023. Early Growth patterns of Bacillus cereus on potato substrate in the presence of low densities of black soldier fly larvae. Microorganisms 11: 1284. https://doi.org/10.3390/microorganisms11051284
Nayak, A., Rühl, M. and Klüber, P., 2024. Hermetia illucens (Diptera: Stratiomyidae): Need, Potentiality, and Performance Measures. Agriculture 14: 8. https://doi.org/10.3390/agriculture14010008
Nyakeri, E.M., Ogola, H.J.O., Ayieko, M.A. and Amimo, F.A., 2017. Valorisation of organic waste material: growth performance of wild black soldier fly larvae (Hermetia illucens) reared on different organic wastes. Journal of Insects as Food and Feed 3: 193-202. https://doi.org/10.3920/JIFF2017.0004
Oonincx, D.G.A.B., van Broekhoven, S., van Huis, A. and van Loon, J.J.A., 2015. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 10: e0144601. https://doi.org/10.1371/journal.pone.0144601
Pakdel, M., Olsen, A. and Bar, E.M.S., 2023. A review of food contaminants and their pathways within food processing facilities using open food processing equipment. Journal of Food Protection 86: 100184. https://doi.org/10.1016/j.jfp.2023.100184
Parodi, A., De Boer, I.J.M., Gerrits, W.J.J., Van Loon, J.J.A., Heetkamp, M.J.W., Van Schelt, J., Bolhuis, J.E. and Van Zanten, H.H.E., 2020. Bioconversion efficiencies, greenhouse gas and ammonia emissions during black soldier fly rearing – a mass balance approach. Journal of Cleaner Production 271: 122488. https://doi.org/10.1016/j.jclepro.2020.122488
Petry, S., Tapprest, J., Maillard, K., Barbut, F., Duquesne, F., Kozak, S., Foucher, N., Bernez-Romand, M., Bridoux, L. and Poquet, I., 2025. Clostridioides difficile in Equidae necropsied in Normandie, France, in 2019-2021: common carriage of toxigenic strains and virulence of emerging ribotype 017. bioRxiv: 2024.2009.2030.615820. https://doi.org/10.1101/2024.09.30.615820
Pinotti, L. and Ottoboni, M., 2021. Substrate as insect feed for bio-mass production. Journal of Insects as Food and Feed 7: 585-596. https://doi.org/10.3920/JIFF2020.0110
Poirier, S., Rué, O., Peguilhan, R., Coeuret, G., Zagorec, M., Champomier-Vergès, M.C., Loux, V. and Chaillou, S., 2018. Deciphering intra-species bacterial diversity of meat and seafood spoilage microbiota using amplicon sequencing: A comparative analysis with 16S rDNA V3-V4 amplicon sequencing. PLoS ONE 13: e0204629. https://doi.org/10.1371/journal.pone.0204629
Quan, J., Wang, Y., Cheng, X., Li, C. and Yuan, Z., 2023. Revealing the effects of fermented food waste on the growth and intestinal microorganisms of black soldier fly (Hermetia illucens) larvae. Waste Management 171: 580-589. https://doi.org/10.1016/j.wasman.2023.https://doi.org/10.002
Rehman, K.U., Hollah, C., Wiesotzki, K., Rehman, R.U., Rehman, A.U., Zhang, J., Zheng, L., Nienaber, T., Heinz, V. and Aganovic, K., 2023. Black soldier fly, Hermetia illucens, as a potential innovative and environmentally friendly tool for organic waste management: a mini-review. Waste Management Research 41: 81-97. https://doi.org/10.1177/0734242x221105441
Rodriguez Diaz, C., Seyboldt, C. and Rupnik, M., 2018. Non-human C. difficile reservoirs and sources: animals, food, environment. In: Mastrantonio, P. and Rupnik, M. (eds.) Updates on Clostridium difficile in Europe: advances in microbiology, infectious diseases and public health, Volume 8. Springer, Cham, pp. 227-243. https://doi.org/10.1007/978-3-319-72799-8_13
Sanahuja, G., Banakar, R., Twyman, R.M., Capell, T. and Christou, P., 2011. Bacillus thuringiensis: a century of research, development and commercial applications. Plant Biotechnology Journal 9: 283-300. https://doi.org/10.1111/j.1467-7652.2011.00595.x
Slorach, P.C., Jeswani, H.K., Cuéllar-Franca, R. and Azapagic, A., 2019. Environmental and economic implications of recovering resources from food waste in a circular economy. Science of the Total Environment 693: 133516. https://doi.org/10.1016/j.scitotenv.2019.07.322
Smits, W.K., Lyras, D., Lacy, D.B., Wilcox, M.H. and Kuijper, E.J., 2016. infection. Nature Reviews Disease Primers 2: 16020. https://doi.org/10.1038/nrdp.2016.20
Stenfors Arnesen, L.P., Fagerlund, A. and Granum, P.E., 2008. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiology Reviews 32: 579-606. https://doi.org/10.1111/j.1574-6976.2008.00112.x
Tkalec, V., Viprey, V., Davis, G., Janezic, S., Sente, B., Devos, N., Wilcox, M., Davies, K., Rupnik, M. and COMBACTE-CDI consortium, 2022. Clostridioides difficile positivity rate and PCR ribotype distribution on retail potatoes in 12 European countries, January to June 2018. Euro Surveillance 27: 2100417. https://doi.org/10.2807/1560-7917.ES.2022.27.15.2100417
van der Spiegel, M., Noordam, M.Y. and van der Fels-Klerx, H.J., 2013. Safety of novel protein sources (insects, microalgae, seaweed, duckweed, and rapeseed) and legislative aspects for their application in food and feed production. Comprehensive Reviews in Food Science and Food Safety 12: 662-678. https://doi.org/10.1111/1541-4337.12032
van Huis, A., 2020. Insects as food and feed, a new emerging agricultural sector: a review. Journal of Insects as Food and Feed 6: 27-44. https://doi.org/10.3920/JIFF2019.0017
van Kessel, K., Castelijn, G., van der Voort, M. and Meijer, N., 2024. Investigation of Bacillus cereus growth and sporulation during Hermetia illucens larval rearing. Heliyon 10: e40912. https://doi.org/10.1016/j.heliyon.2024.e40912
Vandeweyer, D., De Smet, J., Van Looveren, N. and Van Campenhout, L., 2021. Biological contaminants in insects as food and feed. Journal of Insects as Food and Feed 7: 807-822. https://doi.org/10.3920/JIFF2020.0060
Vandeweyer, D., Lievens, B. and Van Campenhout, L., 2020. Identification of bacterial endospores and targeted detection of foodborne viruses in industrially reared insects for food. Nature Food 1: 511-516. https://doi.org/10.1038/s43016-020-0120-z
Walia, K., Kapoor, A. and Farber, J.M., 2018. Qualitative risk assessment of cricket powder to be used to treat undernutrition in infants and children in Cambodia. Food Control 92: 169-182. https://doi.org/10.1016/j.foodcont.2018.04.047
Warriner, K., Xu, C., Habash, M., Sultan, S. and Weese, S.J., 2017. Dissemination of Clostridium difficile in food and the environment: significant sources of C. difficile community-acquired infection? Journal of Applied Microbiology 122: 542-553. https://doi.org/10.1111/jam.13338
Wilcks, A., Jayaswal, N., Lereclus, D. and Andrup, L., 1998. Characterization of plasmid pAW63, a second self-transmissible plasmid in Bacillus thuringiensis subsp. kurstaki HD73. Microbiology 144: 1263-1270. https://doi.org/10.1099/00221287-144-5-1263
Wu, N., Ma, Y., Yu, X., Wang, X., Wang, Q., Liu, X. and Xu, X., 2024. Black soldier fly larvae bioconversion and subsequent composting promote larval frass quality during pig and chicken manure transformation process. Bioresource Technology 402: 130777. https://doi.org/10.1016/j.biortech.2024.130777
Xiao, X., Mazza, L., Yu, Y., Cai, M., Zheng, L., Tomberlin, J.K., Yu, J., van Huis, A., Yu, Z., Fasulo, S. and Zhang, J., 2018. Efficient co-conversion process of chicken manure into protein feed and organic fertilizer by Hermetia illucens L. (Diptera: Stratiomyidae) larvae and functional bacteria. Journal of Environmental Management 217: 668-676. https://doi.org/10.1016/j.jenvman.2018.03.122
Yang, S., Wang, Y., Liu, Y., Jia, K., Zhang, Z. and Dong, Q., 2023. Cereulide and emetic Bacillus cereus: characterizations, impacts and public precautions. Foods 12: 833.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}