Abstract
The black soldier fly (BSF; Hermetia illucens; Diptera: Stratiomyidae) is a novel mini-livestock species now reared in the trillions annually around the globe. As BSF farming continues to grow, research on how to protect these animals’ welfare in farmed settings is also essential. Currently, industry standard practice involves not feeding BSF adults and providing only water due to a misconception that BSF adults cannot eat. However, BSF adults have functional mouthparts and digestive systems, gravitate towards and consume food when it is provided, and live longer when provided with food. Therefore, current industry practices, where only water is provided, do not keep BSF adults free from hunger. In order to address this welfare concern, we studied the carbohydrate and protein preferences of BSF adults as well as their preferred protein to carbohydrate ratio and overall carbohydrate concentration. We assessed three carbohydrates (honey, white sugar, and molasses) and three proteins (yeast, agar, and casein). Using the maximally preferred diet (1:8 ratio of yeast to molasses, with molasses concentration set to 5%), we assessed BSF adult mating behaviour, egg mass, and sex-specific longevity on the diet vs in the water-only conditions that are currently standard in the industry. We found that BSF were observed to mate more frequently, laid more eggs on days 2-5 post-eclosion, and lived 4-6 days longer when given access to their preferred diet. Our data suggest that feeding adult BSF their preferred diet could improve animal welfare (by keeping the animals free from hunger and allowing for natural feeding-related behaviors) while also improving the number of eggs laid per day.
1 Introduction
The insects as food and feed industry is a rapidly growing industry that rears insects as a source of animal protein for human, pet, or other livestock animal consumption. Insects can contribute to more sustainable agriculture via promoting local, circular food systems; further, many insects are capable of converting waste into food, feed, and other fuels, facilitating economic and food security (Chia et al., 2019; Parodi et al., 2022; Purkayastha and Sarkar, 2022; van Huis, 2013; van Huis and Gasco, 2023; van Huis and Tomberlin, 2017). Accordingly, demand for insect protein is growing rapidly (de Jong and Nikolik, 2021). For instance, demand is projected to grow 22-fold from 2021 to 2030 in the UK (World Wildlife Fund and Tesco, 2021). Depending on which markets begin to use insect protein (fishmeal replacement, poultry feed, pet feed, etc.), trillions of additional insects are predicted to be farmed each year (Rowe, 2020; de Jong and Nikolik, 2021).
The welfare of farmed insects can be expected to be of concern if they are sentient (though see Dawkins, 2017 for reasons to consider animal welfare that do not rely on sentience). The sentience of insects, and their possible affective states, are currently topics of empirical uncertainty (Adamo, 2016, 2019; Barron and Klein, 2016; Elwood, 2016, 2023; Barrett and Fischer, 2025; Gibbons and Sarlak, 2020; Kakrada and Colombo, 2022; Klein and Barron, 2016; Lambert et al., 2021; Mikhalevich and Powell, 2020; Overgaard, 2021). However, a review of the most recent research in insect neurobiology and behaviour suggests that insect sentience is plausible at the adult life stage, especially in the Diptera (flies and mosquitoes; Gibbons et al., 2022). Adult dipterans met 6/8 criteria using the Crump et al. (2022) framework for evaluating invertebrate sentience, which is considered strong evidence for sentience. These data suggest that, from a precautionary perspective (Birch, 2017; Knutsson and Munthe, 2017; Röcklinsberg et al., 2017; van Huis, 2021), the welfare of adult insects of this order should be considered (and may be influenced by developmental factors related to larval experiences as well as adult experience; Barrett et al., 2023).
In line with this evidence, producers, legislators, academics, industry trade organizations, animal advocacy groups, the media, and ethicists have all called for consideration of the welfare of these mini-livestock during rearing and slaughter (Barrett and Adcock, 2023; Barrett and Fischer, 2023; Bear, 2019; Boppré and Vane-Wright, 2019; Council on Animal Affairs, 2018; de Goede et al., 2013; Delvendahl et al., 2022; EuroGroup for Animals, 2023; Fukada et al., 2023; IPIFF, 2019; Klobučar and Fisher, 2023; Kortsmit et al. 2023; Reynolds, 2023; Sebo and Schukraft, 2021; van Huis, 2021; Voulgari-Kokota et al., 2023). Considering insect welfare also has the potential to improve economic productivity, mitigate social risk, and create product differentiation opportunities in this emerging industry (Barrett and Adcock, 2023), providing economic as well as ethical reasons for improving farmed insect welfare.
The most popular farmed insect for food and feed is the black soldier fly (BSF; Hermetia illucens; Rumbos and Athanassiou, 2021). As of 2024, it is projected that 1.2-1.8 trillion BSF are reared annually (Shah, 2024); however, facilities are currently in construction that would farm around 600 billion to more than 1 trillion individuals each (60 000 tons of insect protein at Innovafeed (2020); see conversion method in Rowe, 2020). Most BSF are slaughtered as larvae and used for feed; however, a subset will be allowed to pupate and eclose, emerging as adult flies to be used as breeders (Dortsman et al., 2017). Adult flies mate in large cages or greenhouses on farms, using a combination of sensory cues to locate mates starting two days after eclosion (reviewed in Lemke et al., 2023; Barrett et al., 2023). Males may engage in aggressive, territorial interactions, at least in the wild (Tingle et al., 1975; Tomberlin et al., 2001), and there is some evidence of female choice in captivity (Giunti et al., 2018). Females lay eggs in large clutches, which generally contain 300-700 eggs each, starting around four days after eclosion (reviewed in Barrett et al., 2023).
BSF broodstock are adult dipterans, the life stage and order for which there is currently the most evidence for insect sentience (Gibbons et al. 2022). A recent review of BSF welfare concluded that inadequate nutrition is a common welfare concern for adult BSF in industry conditions (Barrett et al., 2023). Adult breeders are routinely not provided with food due to a misconception that likely arose from Sheppard et al., (2002), a study that was widely interpreted to mean that the adults cannot eat at the adult life stage and instead rely entirely on large stores of fat from the larval stage. Although BSF can reproduce using their energetic reserves from the larval period, adult BSF are now known to have functional mouthparts linked to a functional digestive system (Bruno et al., 2019; Oliveira et al., 2016), and feeding of adult BSF has been recommended for decades (Furman et al., 1959), as adult BSF gravitate towards carbohydrate sources when provided (Nakamura et al., 2016). Further, their longevity and reproductive output increases substantially when given carbohydrates, or certain protein-carbohydrate mixtures, in captive settings (Bertinetti et al., 2019; Bruno et al., 2019; Chia et al., 2018; Klüber et al., 2023; Macavei et al., 2020; Nakamura et al., 2016; Romano et al., 2020).
Accordingly, it is important to provide producers with empirically-driven best practices on what to feed adult BSF used as breeders in their facilities. This ensures animals are free from hunger, a key component of many farmed animal welfare frameworks like the 5 Freedoms and 5 Domains (Brambell, 1965; Mellor and Reid, 1994; IPIFF, 2019). Animals, including insects, often require both a carbohydrate and protein source in order to be adequately fed (though macronutrient needs can differ by sex, reproductive status, or age in other flies: Camus et al., 2018; Jensen et al., 2015; Lee et al., 2008). Further, to promote high welfare the feed should be delivered using a mechanism that is safe and accessible for the animals; preferably, this would also be easy to keep clean to prevent disease or contamination. Finally, an important part of animal welfare is considering what animals might want (or prefer) to eat (Stamp Dawkins, 2021).
Prior research on adult BSF nutrition fails to address most of the above points that would be useful to generating welfare guidance. Studies to date generally either (1) investigate only carbohydrate source (e.g. Nakamura et al., 2016, Macavei et al., 2020); (2) develop feed mixtures that are too gelatinous for nutrients to be continuously accessible to insects with sponging mouthparts; (3) do not assess BSF dietary preferences (Bertinetti et al., 2019); (4) do not standardize concentrations across conditions (e.g. Romano et al., 2020); or (5) fail to assess macronutrient ratios when providing both proteins and carbohydrates (e.g. Bertinetti et al., 2019, Thinn and Kainoh, 2022). Addressing each of these gaps is necessary to provide producers with greater guidance on best practices for feeding adult BSF.
Adult BSF feeding has generally been tested using white sugar or honey (Bertinetti et al., 2019; Furman et al., 1959; Klüber et al., 2023; Macavei et al., 2020; Nakamura et al., 2016; Oonincx et al., 2016; Romano et al., 2020; Thinn and Kainoh, 2022). However, molasses is frequently used for feeding other fly species (e.g. Chakraborty et al., 2011). Proteins tested to date include agar/milk mixed with sugar (Bertinetti et al., 2019), which contain proteins that are also common in other fly diets (e.g. Chakraborty et al., 2011) even if milk itself is rarely provided. Some studies have also tested Spirulina and Chlorella powder in water (Klüber et al., 2023), which appeared to have minimal effect on BSF longevity and nutrition, and are not common in fly diets. Yeast is commonly used in mass-reared fly diets (e.g. Chakraborty et al., 2011) and is also available to wild insects in the environment, serving as an important protein in many wild insect diets (Madden et al., 2018). No studies have tested yeast as a protein source for adult BSF.
Therefore, we tested BSF preferences in response to standardized concentrations of white sugar, molasses, and honey (for carbohydrates) and three proteins commonly used for rearing other fly species: casein (reducing the fat content confound associated with the use of milk itself), yeast, and agar. To test preferences, we used an industry-cage-scale version of a classic gravimetric food consumption assay (e.g. Diegelmann et al., 2017). For our initial carbohydrate preference test, we tested two ages. We chose 0-24 hour old flies for one test, as flies of this age are unmated and not yet laying eggs (Sheppard et al., 2002) and therefore represents flies earlier in the reproductive trajectory. We also tested 96-120 hour old flies (0-24 hour old flies that had been held with only water for four days), as this is the age of mating and oviposition (Sheppard et al., 2002). Testing these two ages allowed us to see if the energetic demands of these reproductive activities changed fly preferences. After determining that fly preferences were stronger at the 4-5-day-old life stage, either due to the feed withdrawal period, their mating status, and/or the effect of aging, we used 4-5-day-old flies for all future preference tests.
After determining the preferred carbohydrate (C) and protein (P), we used these preferred compounds to assess BSF preferences for different macronutrient ratios (varying from 1:2 P source: C source to 1:32 P source: C source, by weight) and different overall carbohydrate concentrations in the solution (from 5 to 20% carbohydrate source by volume). Finally, we assessed the impact of water only (industry standard practice) versus the most preferred macronutrient ratio diets (P source:C source = 1:8) on sex-specific longevity, cage-level reproductive output, and BSF mating behaviour. We use these data to make recommendations for producers interested in improving adult BSF welfare and farm productivity, describing what protein and carbohydrates to feed their flies and what macronutrient ratios to use.
2 Materials and methods
Pupae sourcing and adult rearing
Pupae were supplied in batches by Evo Conversion Systems (College Station, Texas) and Fluker Farms (Port Allen, LA, USA); each treatment received the same number of pupae from each batch. Pupae were placed in mesh cages (30 cm cubed) at 25-27 °C temperature, 75% humidity, 12:12 L:D cycles to eclose in moist potting soil to prevent desiccation before being sorted into treatment groups by sex on the day of eclosion as they were placed into the industry-sized cages (see below). All pupae used for an experiment were sourced from the same supplier.
Adults undergoing each experiment were reared in the same abiotic conditions as the pupae, but in standard industry mating cages (1 m × 1.2 m × 2.4 m, H × W × L; SPR AgTech) and with special lights designed for the adult BSF visual spectrum (SPR AgTech). All adults were in mixed-sex conditions for their entire lifespan, with equal numbers of males and females, and reared at the densities described in the sections that follow for each experiment.
Assessing dietary preferences by age
Ingredients for composing adult diets (yeast, casein, agar, dextrose, honey, and molasses) were sourced from Carolina Biological and ShopRite Supermarkets. Foods were cooked prior to loading in the feeders to ensure sterilization and yeast inactivation; this prevented growth in the feeders.
BSF adults (100 male and 100 female) were collected 0-24 hour post-eclosion and placed into breeding cage. Three cages of BSF adults were provided with the six different liquid feeds and water (300 g each; in plastic honeybee feeders) immediately, while three cages were incubated for 96 hours with water only before being provided with liquid feed. After 48 hours in with the flies, liquid food sources were removed and re-weighed to assess the combined mass lost due to feeding and evaporation. Environmental controls were used to account for evaporation for all liquid feed sources (placed directly outside of breeding cages for the same 48 hours, to exclude feeding but allow for evaporation).
For the carbohydrate preference tests the following liquid feed sources were used: water, 5% honey (ShopRite Grade A Honey) in water, 10% honey in water, 5% dextrose (Carolina Biological) in water, 10% dextrose in water, 5% molasses (Grandma’s Molasses Original) in water, and 10% molasses in water. For the protein preference tests, the liquid food sources used were water, 5% agar (Carolina Biological) in water, 10% agar in water, 5% casein (Carolina Biological) in water, 10% casein in water, 5% yeast (Red Star Dry Yeast) in water, 10% yeast in water. All ingredients were measured in w/v. Protein preference was only tested after 96 hours of incubation.
For the relative carbohydrate and protein concentration preference test, molasses was used as the carbohydrate and yeast as the protein given prior test results. 100 male and 100 female 0-24 hour old BSF adults were incubated in a breeding cage for 96 hours with water (3 replicate cages). Following incubation, BSF adults were provided six liquid food sources in honeybee feeders (300 g each; water, 1:2 ratio of protein source to carbohydrate source in water, 1:4 ratio of protein to carbohydrate in water, 1:8 ratio of protein to carbohydrate in water, 1:16 ratio of protein to carbohydrate in water, 1:32 ratio of protein to carbohydrate in water) for 48 hours. Ratios were calculated while maintaining a standard 5% (w/v) molasses concentration. After 48 hours, foods were removed and the combined effects of food consumption and evaporation were determined gravimetrically. Environmental controls were used to account for evaporation for all liquid food sources (placed directly outside of breeding cages).
For the carbohydrate percent preference test, 100 male and 100 female 0-24 hour old adults were incubated with water only for 96 hours with water (3 replicate cages). Following incubation, flies were provided a water (0%) control, and then a 5, 10, 15 and 20% molasses diet, all with a 1:8 ratio of protein source to carbohydrate source (with yeast as the protein), in honeybee feeders (300 g each). After 48 hours, diets were removed and the combined effects of food consumption and evaporation was determined gravimetrically. Environmental controls were used to account for evaporation for all liquid food sources (placed directly outside of breeding cages).
Assessing impacts of diet on longevity
BSF adults (100 male and 100 female) were collected 0-24 hour post-eclosion and placed into a breeding cage (3 replicate cages per treatment). BSF adults were immediately provided liquid food sources (either water only or their preferred yeast: molasses diet). Liquid food sources were replaced every two days. Deceased flies were counted, visually sexed using the protocol found in Julita et al., (2020), and removed from the cage daily at the same time. Tracking of survivorship was performed until all BSF adults were deceased within the breeding cage.
Assessing impacts of diet on reproduction
BSF adults (100 male and 100 female) were collected 0-24 hour post-eclosion and placed into a breeding cage (3 replicate cages per treatment). BSF larvae (supplied in a batch from Fluker Farms) were housed with food (Gainesville diet) in open shoe box containers with mesh lids to exclude adults and placed into the breeding cage; this was used as an oviposition attractant. BSF adults were immediately provided liquid food sources (either water only or their preferred yeast: molasses diet) that was placed directly over the BSF larvae housing enclosure. Standard Egg traps (Evo Conversion Systems), which are plastic frames fitted with tongue depressors, were also placed directly over the BSF larvae housing enclosure. BSF adults were immediately provided liquid food sources (either water only or their preferred yeast: molasses diet). Liquid food sources were replaced every two days.
The standard egg traps were collected and replaced daily. Collected egg traps were thoroughly cleaned to remove all eggs present. We quantified the total mass of eggs laid per day per condition in g; to obtain the number of eggs, we divided this weight by the standard average weight of a BSF egg, 0.03 mg (Dortsman et al., 2017; Macavei et al., 2020). BSF adult mating behaviour was also quantified through direct observation daily at 1100, 1200 and 1300 hours as observed mating peaks at 1200 in Julita et al. (2020).
Statistical analysis
GraphPad Prism v. 10.4.0 was used for statistical analyses. Data were checked for normality (Shapiro-Wilk normality test) prior to choosing the test to run. A one-way ANOVA was used to analyze 0-24 hour and 4-5-day-old BSF adult dietary preference data, with a Tukey’s Multiple Comparisons Test. Survival was analysed using a Log-Rank Mantel Cox test. All alpha set to 0.05. A two-way repeated measures (RM) ANOVA with Geisser-Greenhouse correction (with cage considered the repeated measure) and a Tukey’s MCT was used to analyse egg lay data. Behavioural observations (number of mating couples) were summed per cage per day; summed observation counts were then analysed using a two-way RM ANOVA (with cage considered the repeated measure) with a Geisser-Greenhouse correction and a Tukey’s MCT. Raw data can be accessed here: https://osf.io/urvjz/files/osfstorage
3 Results
Carbohydrate and protein preferences – 0-24 hours and 4-5 days old
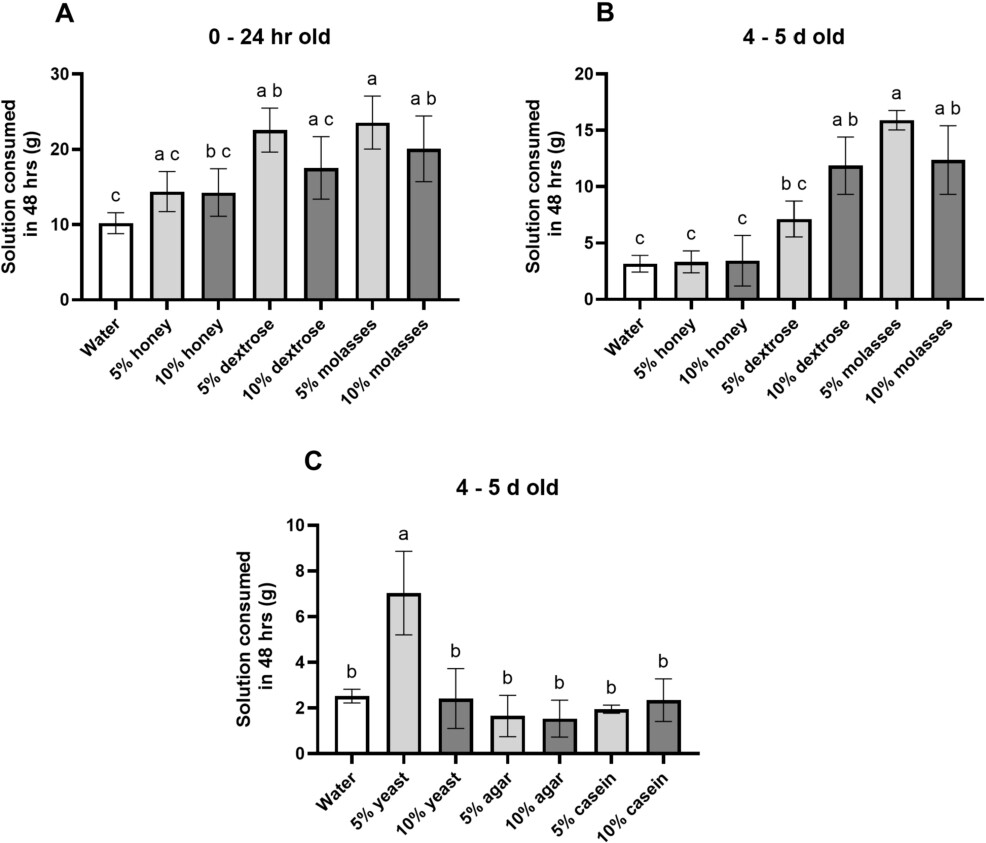
There was a significant difference in fly carbohydrate preference at both 0-24 hr old (Figure 1A; ANOVA:

Fly carbohydrate (A, B) and protein (C) preferences at 0-24 hours or 4-5 days (96-120 h) old. (A) 0-24-hour-old flies consumed more 5% dextrose, 5% molasses and 10% molasses than water over 48 h. (B) 4-5-day-old flies consumed more 10% dextrose, 5% molasses and 10% molasses than water over 48 h. Generally, dextrose and molasses were preferred over water and honey. (C) Flies consumed more 5% yeast than water or any other protein source at 4-5 days old. For (A-C) ANOVA with Tukey’s MCT; letters indicate statistically significant differences among conditions. Means with SD error bars.
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
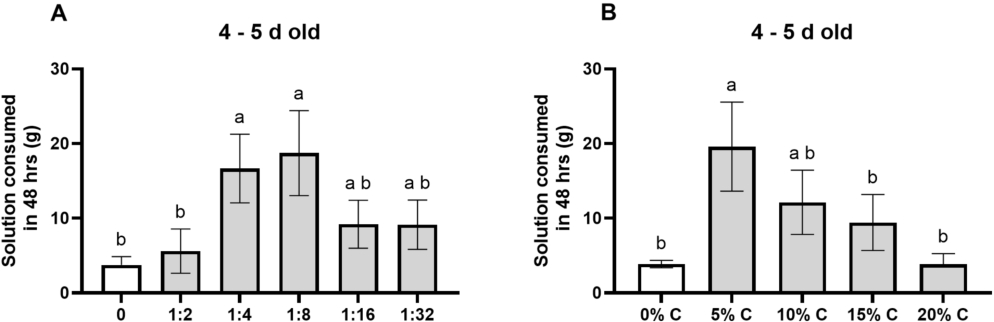
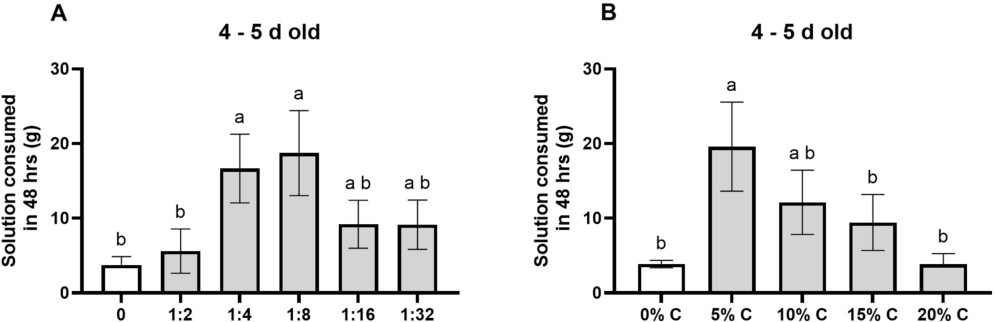
Fly preference for diets of different yeast: molasses ratios (A) and of different carbohydrate concentrations at a 1:8 yeast:molasses ratio (B). (A) Flies preferred 1:4 and 1:8 yeast: molasses ratio diets over water and 1:2 ratio diets (all
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
At 0-24 hours old, flies consumed significantly more 5% dextrose (
There was also a significant difference in fly protein consumption at 4-5 days old (Figure 1C; ANOVA:
Macronutrient ratio and diet concentration preferences
Having determined yeast was the preferred protein, and molasses the preferred carbohydrate, we next tested preferred carbohydrate and protein source ratios. There was a significant difference in fly consumption of diets of different ratios (Figure 2A; ANOVA:
As a result, we chose a 1:8 yeast: molasses diet ratio for our next test of concentration preference. There was a significant difference in fly consumption of 1:8 yeast: molasses diets based on different carbohydrate concentrations (Figure 2B; ANOVA:
Sex-specific longevity on preferred diets
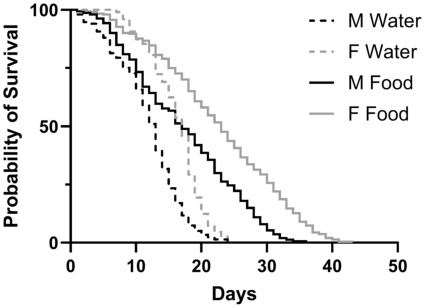
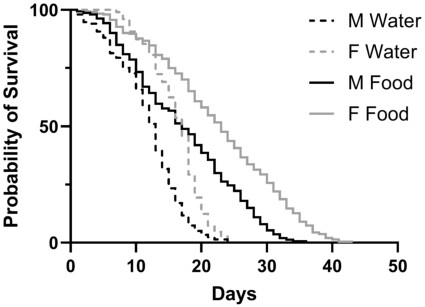
0-24-hour-old males and females lived longer when fed the 1:8 yeast: molasses diet at a 5% molasses concentration than when provided with only water (Figure 3; Mantel-Cox Log Rank Test; males:
Survival of male and female BSF given only water or a diet of 1:8 yeast: molasses (5% molasses concentration). Males and females were fed a 1:8 yeast: molasses diet lived longer than when they were only provided water. The median survival of unfed or fed females was greater than unfed or fed males, respectively (all
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
Reproduction on preferred diets
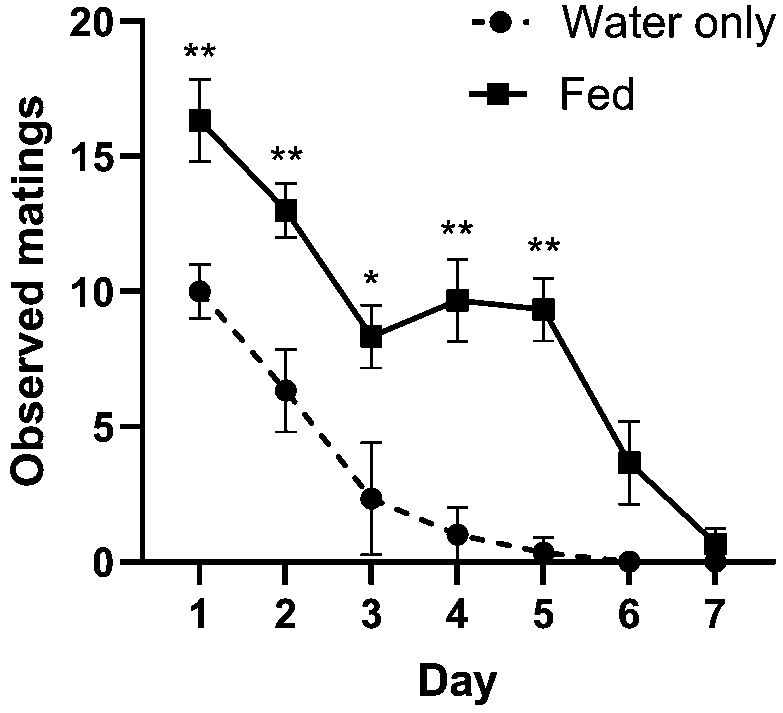
Diet, day, and the interaction of diet and day all had an effect on observed instances of actively mating couples. Day had the strongest effect on the number of observed mating couples (Figure 4; two-way RM ANOVA: 59.44% of total variation,
Number of observed mating pairs in cages provided with water only versus a diet of 1:8 yeast:5% molasses. More matings were observed on days 1-5 when flies were fed a diet of yeast and molasses compared to being provided only water (Tukey’s MCT; all
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
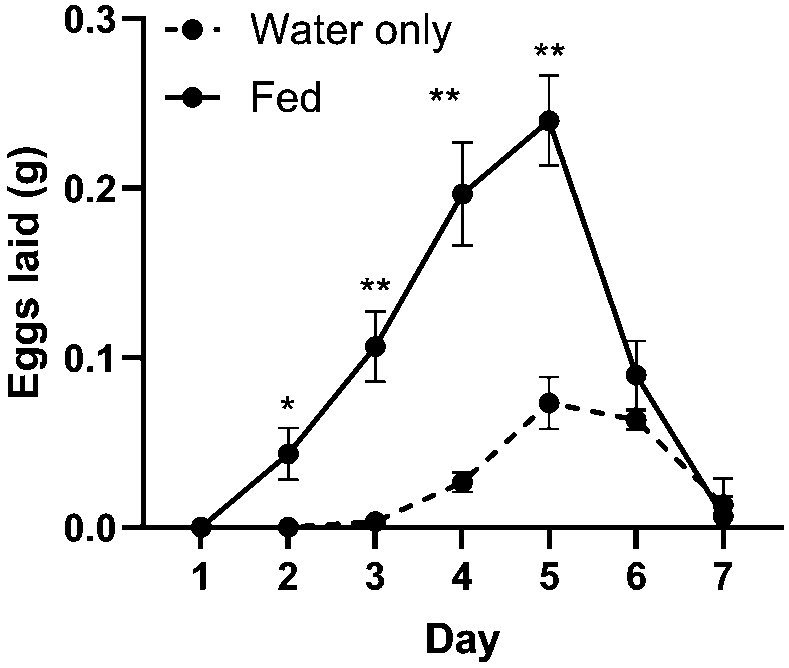
Mass of eggs laid by females in cages provided with water only versus a diet of 1:8 yeast:5% molasses. Females laid more eggs per day on days 2-5, and began egg lay sooner, when fed a diet of yeast and molasses compared to being provided only water (all
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
As with mating behaviour, diet, day, and the interaction of diet and day all had an effect on the mass of eggs laid in the cage. Day had the strongest effect on egg mass in the cage (Figure 5; two-way RM ANOVA: 51.61% of total variation,
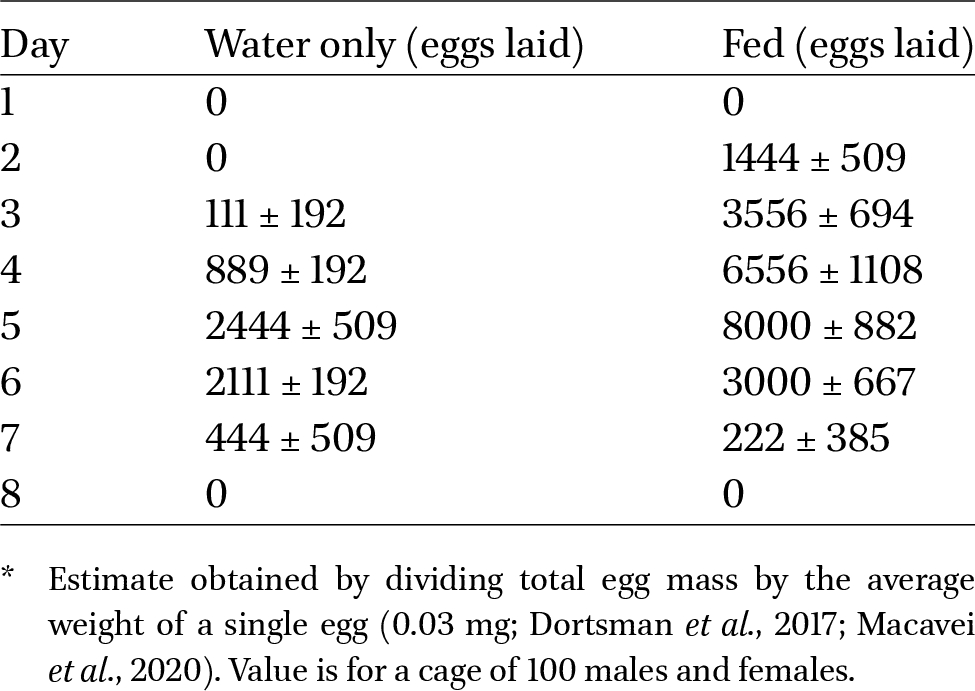
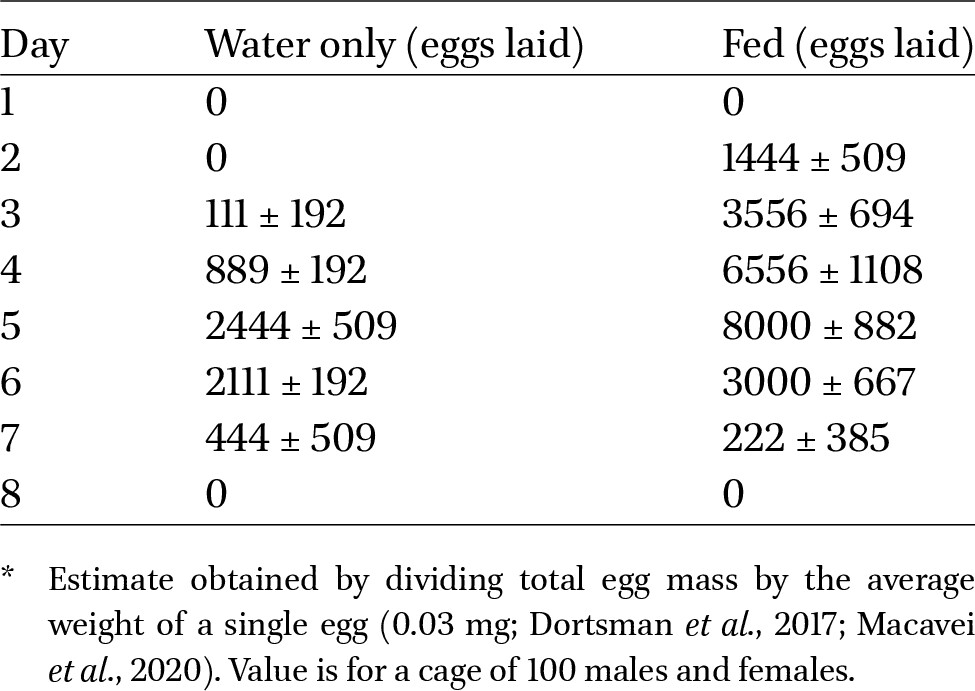
Egg lay began on day 2 for fed treatments and on day 3 for unfed treatments; it peaked on day 5 in both treatments and declined back to 0 g of laid eggs by day 8 in both treatments (Table 1).
Estimated number of eggs laid* in cages of fed vs unfed adult BSF
Citation: Journal of Insects as Food and Feed 12, 4 (2026) ; 10.1163/23524588-bja10295
4 Discussion
Despite mounting evidence that adult black soldier flies do have the ability to consume nutrients, including behavioural, anatomical, and physiological data, a commonly held misconception in the industry is that adult BSF cannot consume food (all data reviewed in Barrett et al. 2023, and see Klüber et al. 2023). Our results add to the growing body of work that demonstrates adult BSF can eat and are benefited by nutrition – but also strengthens the case by demonstrating adult preferences in protein, carbohydrates, and macronutrient ratios, and by showing that both lifespan and reproduction can be benefited through the inclusion of feed for adult BSF.
We found that mixed-sex cages of flies preferred molasses over white sugar and honey for carbohydrates. Molasses can contain a diverse array of micronutrients (e.g. iron, magnesium, etc.); future studies might consider if micronutrients play a role in mediating this carbohydrate preference in BSF. Further, we found that flies preferred yeast over casein or agar; yeast has never before been tested as a protein source for BSF, but is commonly included in standard fruit fly diets (as is molasses; e.g. Fiocca and Barrett et al. 2019) and is a widely available protein source for flies in the wild (Madden et al., 2018). In fruit flies, olfactory cues allow mated females to recognize yeast as an appropriate protein source (Corrales-Carvajal et al. 2016); similar cues could be mediating the response of 4-5 day old female BSF to the yeast solution.
The preferred yeast: molasses ratio was 1:4 or 1:8 (out of a range of 1:2-1:32). These data match studies in fruit flies, which regulate their intake to a P:C ratio of 1:4, leading to maximized fitness (as measured by egg production). Other diets could maximize longevity (e.g. P:C of 1:16) or egg-laying rate (P:C of 1:2), though at the expense of overall fitness (Lee et al., 2008). While we did not directly test the effects of other yeast: molasses ratios on adult BSF longevity, egg lay rate, or fitness, BSF fed only sugar water had survival in excess of 40-70 days, whether mated (Macavei et al., 2020) or unmated (Nakamura et al., 2016); these trends match results in many other animals demonstrating that high-carbohydrate, low-protein diets may increase longevity at the expense of fitness (e.g. Solon-Biet et al., 2014). By contrast, Bertinetti et al. (2019) found that egg lay could be benefited without such significant extensions to longevity when providing milk or agar as protein in the feed, similar to our results with yeast. However, where Bertinetti et al., (2019) found that lifetime egg production increased by 2.79-fold on the milk protein diet across the first 18 days of adult life, we demonstrate a 3.8-fold increase in egg production on the preferred diet in just the first seven days. In general, studies on BSF demonstrate that fitness and/or longevity are maximized when BSF are fed almost any combination of macronutrients, compared to the water-only conditions more typical in industry settings (Bertinetti et al., 2019; Bruno et al., 2019, Chia et al., 2018, Nakamura et al., 2016, Macavei et al., 2020,; though see some results in Klüber et al., 2023).
The split in preference between 1:4 and 1:8 yeast:molasses ratios may represent a sex-specific split where females might prefer a higher protein diet (important for producing eggs) and males might prefer more carbohydrates (important for the energetically-intensive processes of flying and mating). Studies in other flies and crickets have demonstrated these kinds of sex-specific dietary preferences (Camus et al., 2019, Maklakov et al., 2008, Sydney et al., 2024). While future studies could investigate if males and females differ in their preferred P:C ratio, in industry settings cages will always be mixed-sex and it will likely only be feasible for producers to provide a single dietary formulation. Finally, at a 1:8 ratio of heat-inactivated yeast to molasses, we found that a 5% molasses concentration was preferred to higher concentrations (15-20%).
On this preferred diet, female flies lived 6 days longer and male flies lived 4 days longer (approx. 25% increase in lifespan) compared to water-only conditions. Flies were observed mating more frequently on days 1-5 post-eclosion when fed; this preceded an increase in the number of eggs per cage laid on days 2-7. Flies began to lay eggs one day sooner in fed conditions, and egg production both peaked and stopped at the same time as the water-only condition.
Based on our data, we recommend producers consider molasses and heat-inactivated yeast (cooked to >60 °C, which prevents fermentation of the feed) for feeding their flies, in a 1:4-1:8 ratio and with a molasses concentration of 5% or less. We provided this feed in plastic, honeybee entrance feeders which prevented feed from becoming messy, made it easy to refill, and largely prevented drownings by allowing flies to land nearby and eat. Further research should investigate preferred P:C ratios based on fly age, mating status, and sex, test fly preference for molasses concentrations between 0 and 5%, address the impacts of micronutrients on fly behaviour, fitness, and welfare, and further optimize feeder design to make feeding as safe as possible for flies and feasible for producers. Importantly, some studies have shown that diet is more important than even lighting conditions for BSF adult reproductive output (Macavei et al., 2020). Given that the industry has developed special lights to maximize BSF mating (SPR AGRTCH), substantially more research and development attention to adult BSF nutrition seems like a critical next step for improving welfare and productivity on farms. This research will be essential to adult BSF diet optimization for the industry, which has the potential to both improve BSF adult welfare (e.g. freedom from hunger) and help meet production goals.
Corresponding author; e-mail: meghbarr@iu.edu
Acknowledgements
We thank Spencer Behmer for a helpful conversation with MRB about this work as it was in progress. We also want to acknowledge both Evo Conversion Systems and Fluker Farms (especially Vance) for timely shipments of BSF pupae on unusual schedules.
Conflict of interest
Meghan Barrett reports a prior relationship (ending in 2023) with Rethink Priorities that includes: consulting or advisory.
Ethical statement
Although no regulations currently exist to guide the ethical use of insects in research, we followed the 2023 Insect Welfare Research Society Guidelines for Protecting and Promoting Insect Welfare in Research (Fischer et al., 2024), except where experimental treatment necessitated the provisioning of inadequate nutrition in order to assess the effects of current industry practices in comparison to our experimental diets. Insects were monitored for disease, inadequate food/water supply, changes in abiotic conditions, etc., daily by a trained member of the research team, including on weekends. In order to transport insects humanely, we opted to have the pupal life stage shipped (where it is possible that ongoing nervous system reorganization may reduce welfare concerns). We minimized subject numbers by sourcing insects from producers specifically for each study instead of maintaining a permanent lab stock colony. We sacrificed all individuals not needed for research – or at the end of experiments – using cold air euthanasia at 4 °C, the best available method given cost and logistical constraints. Whenever possible, euthanasia occurred during the pupal life stage.
Funding
Rethink Priorities provided funding from EA Animal Welfare Funds to EAW at Holy Family University for this work.
References
Adamo, S.A., 2016. Do insects feel pain? A question at the intersection of animal behaviour, philosophy and robotics. Animal Behaviour 118: 75-79. https://doi.org/10.1016/j.anbehav.2016.05.005
Adamo, S.A., 2019. Is it pain if it does not hurt? On the unlikelihood of insect pain. The Canadian Entomologist 151: 685-695. https://doi.org/10.4039/tce.2019.49
Barrett, M. and Adcock, S.J.J., 2024. A robust field of insect welfare science: An integral piece of growing IAFF industry success. Journal of Insects as Food and Feed 10: 517-531. https://doi.org/10.1163/23524588-20230126
Barrett, M., Chia, S., Fischer, B. and Tomberlin, J.K., 2023. Welfare considerations for farming black soldier flies, Hermetia illucens (Diptera: Stratiomyidae): a model for the insects as food and feed industry. Journal of Insects as Food and Feed 9: 119-148. https://doi.org/10.3920/JIFF2022.0041
Barrett, M. and Fischer, B., 2023. Challenges in farmed insect welfare: Beyond the question of sentience. Animal Welfare 32: e4. https://doi.org/10.1017/awf.2022.5
Barrett, M. and Fischer, B., 2025. The era beyond Eisemann et al. (1984): Insect pain in the 21st century.. The Quarterly Review in Biology 100: 161-187. https://doi.org/10.1086/737365
Barron, A.B. and Klein, C., 2016. What insects can tell us about the origins of consciousness. Proceedings of the National Academy of Sciences of the United States of America 113: 4900-4908. https://doi.org/10.1073/pnas.1520084113
Bear, C., 2019. Approaching insect death: understandings and practices of the UK’s edible insect farmers. Society and Animals 27: 751-768. https://doi.org/10.1163/15685306-00001871
Bertinetti, C., Samayoa, A.C. and Hwang, S.-Y., 2019. Effects of feeding adults of Hermetia illucens (Diptera: Stratiomyidae) on longevity, oviposition, and egg hatchability: insights into optimizing egg production. Journal of Insect Science 19: 19. https://doi.org/10.1093/jisesa/iez001
Birch, J., 2017. Animal sentience and the precautionary principle. Animal Sentience 16: 1200. https://doi.org/10.51291/2377-7478.1200
Boppré, M. and Vane-Wright, R.I., 2019. Welfare dilemmas created by keeping insects in captivity. In: Carere, C. and Mather, J. (eds.) The welfare of invertebrate animals. Springer, Cham, pp. 23-67. https://doi.org/10.1007/978-3-030-13947-6_3
Bruno, D., Bonelli, M., Cadamuro, A.G., Reguzzoni, M., Grimaldi, A., Casartelli, M. and Tettamanti, G., 2019. The digestive system of the adult Hermetia illucens (Diptera: Stratiomyidae): morphological features and functional properties. Cell and Tissue Research 378: 221-238. https://doi.org/10.1007/s00441-019-03025-7
Camus, M.F., Huang, C.-C., Reuter, M. and Fowler, K., 2018. Dietary choices are influenced by genotype, mating status, and sex in Drosophila melanogaster. Ecology and Evolution 8: 5385-5393. https://doi.org/10.1002/ece3.4055
Camus, M.F., Piper, M.D.W. and Reuter, M., 2019. Sex-specific transcriptomic responses to changes in the nutritional environment. eLife 8: e47262. https://doi.org/10.7554/eLife.47262
Chakraborty, R., Vepuri, V., Mhatre, S.D., Paddock, B.E., Miller, S., Michelson, S.J., Delvadia, R., Desai, A., Vinokur, M., Melicharek, D.J., Utreja, S., Khandelwal, P., Ansaloni, S., Goldstein, L.E., Moir, R.D., Lee, J.C., Tabb, L.P., Saunders, A.J. and Marenda, D.R., 2011. Characterization of a Drosophila Alzheimer’s Disease model: pharmacological rescue of cognitive defects. PLoS ONE 6: e20799. https://doi.org/10.1371/journal.pone.0020799
Chia, S.Y., Tanga, C.M., Osuga, I.M., Mohamed, S.A., Khamis, F.M., Salifu, D., Sevgan, S., Fiaboe, K.K.M., Niassy, S., van Loon, J.J.A., Dicke, M. and Ekesi, S., 2018. Effects of waste stream combinations from brewing industry on performance of black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). PeerJ 6: e5885. https://doi.org/10.7717/peerj.5885
Chia, S.Y., Tanga, C.M., van Loon, J.J. and Dicke, M., 2019. Insects for sustainable animal feed: inclusive business models involving smallholder farmers. Current Opinion in Environmental Sustainability 41: 23-30. https://doi.org/10.1016/j.cosust.2019.09.003
Corrales-Carvajal, V.M., Faisal, A.A. and Ribiero, C., 2016. Internal states drive nutrient homeostasis by modulating exploration-exploitation trade-off. eLife 5: e19920. https://doi.org/10.7554/eLife.19920
Council on Animal Affairs, 2018. The emerging insect industry. Council on Animal Affairs. Available online at https://english.rda.nl/documents/2018/09/03/the-emerging-insect-industry
Crump, A., Browning, H., Schnell, A., Burn, C. and Birch, J., 2022. Sentience in decapod crustaceans: A general framework and review of the evidence. Animal Sentience 32: 1691. https://doi.org/10.51291/2377-7478.1691
Dawkins, M.S., 2017. Animal welfare with and without consciousness. Journal of Zoology 301: 1-10. https://doi.org/10.1111/jzo.12434
de Goede, D., Erens, J., Kapesomenou, E. and Peters, M., 2013. Large scale insect rearing and animal welfare. In: Röcklinsberg, H. and Sandin, P. (eds.) The ethics of consumption. Wageningen Academic Publishers, Wageningen, pp. 236-243. https://doi.org/10.3920/978-90-8686-784-4_38
de Jong, B. and Nikolik, G., 2021. No longer crawling: Insect protein to come of age in the 2020s. RaboBank, Utrecht.
Delvendahl, N., Rumpold, B.A. and Langen, N., 2022. Edible Insects as Food-Insect Welfare and Ethical Aspects from a Consumer Perspective. Insects 13: 121. https://doi.org/10.3390/insects13020121
Dortmans, B.M.A., Diener, S., Verstappen, B.M. and Zurbrügg, C., 2017. Black soldier fly biowaste processing – a step-by-step guide. Eawag-Swiss Federal Institute of Aquatic Science and Technology, Dübendorf. Available online at https://www.eawag.ch/fileadmin/Domain1/Abteilungen/sandec/publikationen/SWM/BSF/BSF_Biowaste_Processing_LR.pdf
Elwood, R., 2016. Might insects experience pain? Animal Sentience 9: 1156. https://doi.org/10.51291/2377-7478.1156
Elwood, R.W., 2023. Behavioural indicators of pain and suffering in arthropods and might pain bite back? Animals 13: 2602. https://doi.org/10.3390/ani13162602
EuroGroup for Animals, 2023. The future of insect farming: where’s the catch? EuroGroup for Animals, Brussels. Available online at https://www.eurogroupforanimals.org/files/eurogroupforanimals/2023-03/The_future_of_insect_farming__where%E2%80%99s_the_catch__final_ver.pdf
Fiocca, K., Barrett, M., Waddell, E.A., Viveiros, J., McNair, C., O’Donnell, S. and Marenda, D.M., 2019. Mannitol ingestion causes concentration-dependent, sex-biased mortality in adults of the fruit fly (Drosophila melanogaster). PLoS ONE 14: e0213760. https://doi.org/10.1371/journal.pone.0213760
Fukada, E.P., Carrasco, M.N., Perez, A.J., Fischer, B. and Drewery, M.L., 2023. U.S. consumer perceptions of insects as livestock feed: Ethical considerations for insects. OSF Preprints. https://doi.org/10.31219/osf.io/ck3ma
Furman, D.P., Young, R.D. and Catts, E.P., 1959. Hermetia illucens (Linnaeus) as a factor in the natural control of Musca domestica Linnaeus. Journal of Economic Entomology 52: 917-921. https://doi.org/10.1093/jee/52.5.917
Gibbons, M., Crump, A., Barrett, M., Sarlak, S., Birch, J. and Chittka, L., 2022. Can insects feel pain? A review of the neural and behavioural evidence. Advances in Insect Physiology 63: 155-229. https://doi.org/10.1016/bs.aiip.2022.10.001
Gibbons, M. and Sarlak, S., 2020. Inhibition of pain or response to injury in invertebrates and vertebrates. Animal Sentience 29: 1649. https://doi.org/10.51291/2377-7478.1649
Giunti, G., Campolo, O., Laudani, F. and Palmeri, V., 2018. Male courtship behaviour and potential for female mate choice in the black soldier fly Hermetia illucens L. (Diptera: Stratiomyidae). Entomologia Generalis 38: 29-46. https://doi.org/10.1127/entomologia/2018/0657
Innovafeed, 2020. Largest Insect Protein Facility in Illinois. New Protein. Available online at https://www.newprotein.net/news/the-worlds-largest-insect-protein-facility-is-coming-to-illinois (accessed 6 October 2023).
IPIFF, 2019. Ensuring high standards of animal welfare in insect production. IPIFF, Brussels. Available online at https://ipiff.org/ensuring-high-standards-of-animal-welfare-in-insect-production/
Jensen, K., McClure, C., Priest, N.K. and Hunt, J., 2015. Sex-specific effects of protein and carbohydrate intake on reproduction but not lifespan in Drosophila melanogaster. Aging Cell 14: 605-615. https://doi.org/10.1111/acel.12333
Julita, U., Fitri, L.L., Putra, R.E. and Permana, A.D., 2020. Mating success and reproductive behavior of black soldier fly Hermetia illucens L. (Diptera, Stratiomyidae) in tropics. Journal of Entomology 17: 117-127. https://doi.org/10.3923/je.2020.117.127
Kakrada, E. and Colombo, M., 2022. Extending the null hypothesis to invertebrate pain sentience. Animal Sentience 32: 1754. https://doi.org/10.51291/2377-7478.1754
Klein, C. and Barron, A., 2016. Insects have the capacity for subjective experience. Animal Sentience 9: 1113. https://doi.org/10.51291/2377-7478.1113
Klobučar, T. and Fisher, D.N., 2023. When do we start caring about insect welfare? Neotropical Entomology 52: 5-10. https://doi.org/10.1007/s13744-022-01023-z
Klüber, P., Arous, E., Zorn, H. and Rühl, M., 2023. Protein- and carbohydrate-rich supplements in feeding adult black soldier flies (Hermetia illucens) affect life history traits and egg productivity. Life 13: 355. https://doi.org/10.3390/life13020355
Knutsson, S. and Munthe, C., 2017. A virtue of precaution regarding the moral status of animals with uncertain sentience. Journal of Agricultural and Environmental Ethics 30: 213-224. https://doi.org/10.1007/s10806-017-9662-y
Kortsmit, Y., van der Bruggen, M., Wertheim, B., Dicke, M., Beukeboom, L.W. and van Loon, J.J.A., 2022. Behaviour of two fly species reared for livestock feed: optimising production and insect welfare. Journal of Insects as Food and Feed 9: 149-169. https://doi.org/10.3920/JIFF2021.0214
Lambert, H., Elwin, A. and D’Cruze, N., 2021. Wouldn’t hurt a fly? A review of insect cognition and sentience in relation to their use as food and feed. Applied Animal Behaviour Science 243: 105432. https://doi.org/10.1016/j.applanim.2021.105432
Lee, K.P., Simpson, S.J., Clissold, F.J., Brooks, R., Ballard, J.W.O., Taylor, P.W., Soran, N. and Raubenheimer, D., 2008. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America 105: 2498-2503. https://doi.org/10.1073/pnas.0710787105
Lemke, N.B., Dickerson, A.J. and Tomberlin, J.K., 2023. No neonates without adults: a review of adult black soldier fly biology, Hermetia illucens (Diptera: Stratiomyidae). Bioessays 45: e2200162. https://doi.org/10.1002/bies.202200162
Macavei, L.I., Benassi, G., Stoian, V. and Maistrello, L., 2020. Optimization of Hermetia illucens (L.) egg laying under different nutrition and light conditions. PLoS ONE 15: e0232144. https://doi.org/10.1371/journal.pone.0232144
Madden, A.A., Epps, M.J., Fukami, T., Irwin, R.E., Sheppard, J., Sorger, D.M. and Dunn, R.R., 2018. The ecology of insect-yeast relationships and its relevance to human industry. Proceedings of the Biological Society 285: 20172733. https://doi.org/10.1098/rspb.2017.2733
Maklakov, A.A., Simpson, S.J., Zajitschek, F., Hall, M.D., Dessmann, J., Clissold, F., Raubenheimer, D., Bonduriansky, R. and Brooks, R.C., 2008. Sex-specific fitness effects of nutrient intake on reproduction and lifespan. Current Biology 18: 1062-1066. https://doi.org/10.1016/j.cub.2008.06.059
Mikhalevich, I. and Powell, R., 2020. Minds without spines: Evolutionarily inclusive animal ethics. Animal Sentience 29: 1993. https://doi.org/10.51291/2377-7478.1993
Nakamura, S., Ichiki, R.T., Shimoda, M. and Morioka, S., 2016. Small-scale rearing of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), in the laboratory: low-cost and year-round rearing. Applied Entomology and Zoology 51: 161-166. https://doi.org/10.1007/s13355-015-0376-1
Oliveira, F.R., Doelle, K. and Smith, R.P., 2016. External morphology of Hermetia illucens Stratiomyidae: Diptera (L.1758) based on electron microscopy. Annual Research and Review in Biology 9: 1-10. https://doi.org/10.9734/ARRB/2016/22973
Oonincx, D.G.A.B., Volk, N., Diehl, J.J.E., van Loon, J.J.A. and Belušič, G., 2016. Photoreceptor spectral sensitivity of the compound eyes of black soldier fly (Hermetia illucens) informing the design of LED-based illumination to enhance indoor reproduction. Journal of Insect Physiology 95: 133-139. https://doi.org/10.1016/j.jinsphys.2016.10.006
Overgaard, M., 2021. Insect consciousness. Frontiers in Behavioural Neuroscience 15. https://doi.org/10.3389/fnbeh.2021.653041
Parodi, A., Ipema, A.F., Van Zanten, H.H.E., Bolhuis, J.E., Van Loon, J.J.A. and De Boer, I.J.M., 2022. Principles for the responsible use of farmed insects as livestock feed. Nature Food 3: 972-974. https://doi.org/10.1038/s43016-022-00641-5
Purkayastha, D. and Sarkar, S., 2022. Sustainable waste management using black soldier fly larva: a review. International Journal of Environmental Science and Technology 19: 12701-12726. https://doi.org/10.1007/s13762-021-03524-7
Reynolds, M., 2023. Insect farming is booming. but is it cruel? Wired. Available online at https://www.wired.com/story/insect-farming-sentience/ (accessed 13 April 2023).
Röcklinsberg, H., Gamborg, C. and Gjerris, M., 2017. Ethical issues in insect production. In: van Huis, A. and Tomberlin, J.K. (eds.) Insects as food and feed: from production to consumption. Wageningen Academic Publishers, Wageningen, pp. 364-379. https://doi.org/10.3920/978-90-8686-849-0
Romano, N. and Fischer, H., 2020. Colour and sugar preferences of adult black soldier fly (Hermetia illucens) (Dipetera: Stratiomyidae) for feeding and oviposition. Journal of Environmental Biology 41: 1132-1137. https://doi.org/10.22438/jeb/41/5(SI)/MS_03
Rowe, A., 2020. Insects raised for food and feed – global scale, practices, and policy. Rethink Priorities. Available online at https://rethinkpriorities.org/publications/insects-raised-for-food-and-feed (accessed 16 April 2023).
Rumbos, C.I. and Athanassiou, C.G., 2021. ‘Insects as food and feed: if you can’t beat them, eat them!’ – To the magnificent seven and beyond. Journal of Insect Science 21: 9. https://doi.org/10.1093/jisesa/ieab019
Sebo, J. and Schukraft, J., 2021. On the torment of insect minds and our moral duty not to farm them. Aeon. Available online at https://aeon.co/essays/on-the-torment-of-insect-minds-and-our-moral-duty-not-to-farm-them (accessed 13 April 2023).
Sheppard, D.C., Tomberlin, J.K., Joyce, J.A., Kiser, B.C. and Sumner, S.M., 2002. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). Journal of Medical Entomology 39: 695-698.
Solon-Biet, S.M., McMahon, A.C., Ballard, J.W.O., Ruohonen, K., Wu, L.E., Cogger, V.C., Warren, A., Huang, X., Pichaud, N., Melvin, R.G., Gokarn, R., Khalil, M., Turner, N., Cooney, G.J., Sinclair, D.A., Raubenheimer, D., Le Couteur, D.G. and Simpson, S.J., 2014. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism 19: 418-430. https://doi.org/10.1016/j.cmet.2014.02.009
Sydney, M.C., Chapman, T. and Perry, J.C., 2024. Diet choice is insensitive to mating in male fruit flies. Animal Behaviour 214: 73-86. https://doi.org/10.1016/j.anbehav.2024.05.010
Thinn, A.A. and Kainoh, Y., 2022. Effect of diet on the longevity and oviposition performance of black soldier flies, Hermetia illucens (Diptera: Stratiomyidae). Japan Agricultural Research Quarterly 56: 211-217. https://doi.org/10.6090/jarq.56.211
Tingle, F.C., Mitchell, E.R. and Copeland, W.W., 1975. The soldier fly, Hermetia illucens, in poultry houses in North Central Florida. Journal of the Georgia Entomological Society 10: 179-183.
Tomberlin, J.K. and Sheppard, D.C., 2001. Lekking behavior of the black soldier fly (Diptera: Stratiomyidae). Florida Entomologist 84: 729-730. https://doi.org/10.2307/3496413
van Huis, A., 2013. Potential of insects as food and feed in assuring food security. Annual Review of Entomology 58: 563-583. https://doi.org/10.1146/annurev-ento-120811-153704
van Huis, A., 2021. Welfare of farmed insects. Journal of Insects as Food and Feed 7: 573-584. https://doi.org/10.3920/JIFF2020.0061
van Huis, A. and Gasco, L., 2023. Insects as feed for livestock production. Science 379: 138-139. https://doi.org/10.1126/science.adc9165
van Huis, A. and Tomberlin, J.K. (eds.), 2017. Insects as food and feed: from production to consumption. Wageningen Academic Publishers, Wageningen. https://doi.org/10.3920/978-90-8686-849-0
Voulgari-Kokota, A., van Loon, M.S. and Bovenkerk, B., 2023. Insects as mini-livestock: Considering insect welfare in feed production. NJAS: Impact in Agricultural and Life Sciences 95: 2191797. https://doi.org/10.1080/27685241.2023.2191797
World Wildlife Fund and Tesco, 2021. The future of feed: A WWF roadmap to accelerating insect protein in UK feeds. World Wildlife Fund, London. Available online at https://www.wwf.org.uk/sites/default/files/2021-06/The_future_of_feed_July_2021.pdf







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}