Abstract
In order to achieve optimal growth and health of farm animals, a different amount/ratio of essential amino acids in the feed is necessary in each phase of life. Knowledge about which amino acids are essential and in which quantities/proportions they are best administered has already greatly advanced the pig and poultry sector and is still being generated today. For the still young, rapidly developing insect sector, this knowledge could mean a major step forward towards breeding optimization. Therefore, in this research the needs of the 2 most commercially reared insect species, namely mealworms (Tenebrio molitor) (MW) and black soldier fly larvae (Hermetia illucens) (BSF), were investigated. The focus was on requirements for lysine, methionine, threonine, phenylalanine and tryptophan based on evidence from the literature and those of pigs. Semi-artificial diets based on at least 25% wheat bran (MW) and chicken feed/water (30/70) (BSF) (basic feed important for structure and certain micronutrients) supplemented with sugar and synthetic amino acids were tested. The diets were isoenergetic and isoproteinous. Of the amino acids studied, one was administered in different doses per experiment. The non-essential glutamic acid was used as a substitute at lower doses. Growth, as measured by weight gain, and survival were observed. The following results were for MW and BSF respectively. For lysine from a content of 0.35 g/100 g diet and 0.53 g/100 g diet no extra growth was noticeable. For methionine this was from 0.13 g/100 g and 0.16 g/100 g. Each time more threonine was given, a significant difference could be noticed and the minimum doses were around 0.42 g/100 g and 0.49 g/100 g. With tryptophan, for MW, there were no differences between the different concentrations. At the lowest dose of 0.06 g/100 g the maximum growth was already reached, whereas for BSF this was at 0.13 g/100 g.
1 Introduction
Essential amino acids play a crucial role in the nutrition of farm animals, which have specific dietary requirements to support optimal growth, development, and overall health. Monogastric animals like poultry and pigs are unable to synthesize certain amino acids endogenously, making it necessary that these essential amino acids are provided through their diets. Methionine, lysine, threonine, tryptophan, and valine are among the key essential amino acids that must be included in feed formulations. These amino acids are vital for protein synthesis, enzyme function, immune response, and various metabolic processes. Striking the right balance and ensuring sufficient levels of essential amino acids in their diets is essential to achieve efficient feed utilization, maximize growth rates and enhance the quality of meat production in both poultry and pig industries (Wu et al., 2014).
For pigs, lysine, methionine (+ cystine), threonine, tryptophan, isoleucine, phenylalanine, valine, leucine, histidine, and arginine (the last two in young animals) are essential. High-quality proteins contain all essential amino acids at acceptable levels, whereas poor-quality proteins lack one or more. Upon entering the digestive tract, proteins are broken down into individual amino acids, absorbed into the bloodstream, and transported throughout the body. These amino acids are then incorporated into various types of proteins to fulfil the body’s diverse needs. Thus, a deficiency in one or more essential amino acid can compromise the pig’s metabolic functions, leading to biological inefficiency (e.g. impaired growth) and potential disease. Experience from practice consistently underscores the importance of high-quality proteins and amino acid availability, particularly during periods of stress, management changes, and immune challenges (van Milgen and Dourmad, 2015).
Insects are not mammals, nor vertebrates, yet research shows that many functions performed by amino acids are also applicable to these invertebrate creatures e.g. growth and the buildup of protein-rich biomass, digestion, antibodies against pathogens and hormone metabolism (Davis, 1975; Chang, 2004). Mediterranean fruit fly, Ceratitis capitata (Wiedemann) larvae died when fed with diets free of 10 exogenous essential amino acids (arginine, isoleucine, leucine, lysine, histidine, methionine, phenylalanine, threonine, tryptophan, and valine) or containing 9 exogenous amino acids with removal of any one of the 10 essential amino acids. Contrarily, when larvae were reared on diets lacking the nonessential amino acids together (alanine, aspartic acid, cystine, glutamic acid, glycine, proline, serine, and tyrosine), or either glycine or serine, they survived but exhibited significantly delayed larval development (Chang, 2004).
Given that mealworms (Tenebrio molitor) (MW) and black soldier fly (Hermetia illucens) (BSF) larvae are best cultivated using waste streams for sustainability, these diets are likely deficient in essential amino acids. Moreover, the more qualitative feeds currently marketed for mealworm rearing might have room for improvement, as they rely on grain sources that often lack lysine and methionine, as is the case for pigs and poultry. Studies from the 1970s indicate that MW larvae require a food source containing the same 10 amino acids that are essential for growth in rats and other vertebrates. Research by Davis (1975) revealed that serine, tyrosine, glutamic acid, and possibly glycine were unnecessary for growth in these insects. Alanine, cystine, proline, and aspartic acid were found to be partially indispensable. According to John et al. (1978), larvae achieved maximum weight gain when receiving reference levels (concentrations equivalent to those found in larval tissues) of all essential amino acids, except for phenylalanine, threonine, and tryptophan. Larvae that were fed 50% of the phenylalanine level found in larval tissues attained maximum growth. Threonine and tryptophan were the limiting amino acids, likely requiring dietary concentrations of over twice those found in larval tissues.
While some rough data regarding protein content is available for mealworms, little is known about the ideal amino acid composition of their diet (Davis, 1975; John et al., 1978; John et al., 1979). Research on BSF larvae is even more limited and has just been initiated in this regard (Spranghers et al., 2019). Knowledge of essential amino acids and their optimal amounts/ratios has significantly benefited the swine and poultry sectors and continues to be accumulated. The emerging insect sector has considerable room for improvement and welcomes new knowledge regarding cultivation optimization. This research contributes to this endeavour.
In order to achieve optimal growth and health of farmed insects, in this study the amount/ratio of essential amino acids was evaluated in 2 most commercially cultivated insect species, namely mealworms and black soldier fly larvae. The focus was on dietary requirements for 5 AAs namely lysine, methionine, threonine, phenylalanine and tryptophan. Threonine, phenylalanine, and tryptophan were selected based on the aforementioned literature. Additionally, lysine and methionine were investigated, as these are the primary limiting amino acids for pigs and poultry and also appear to be essential for insects.
2 Materials and methods
The mealworms and BSF larvae in this study were reared at the Inagro Insect Research Centre in Roeselare, Belgium. They were both kept in 60 × 40 cm plastic crates at a temperature of 27 ± 1 °C, 60 ± 3% relative humidity and in complete darkness (except during maintenance). The mealworms and BSF larvae were fed ad libitum with INSECTUS Mealworm Grow (Mijten NV, Bekkevoort, Belgium) with fermented chopped chicory roots as moisture source and Farm1crumble chicken feed (HobbyFirst, Kortrijk, Belgium) mixed with water (30/70), respectively.

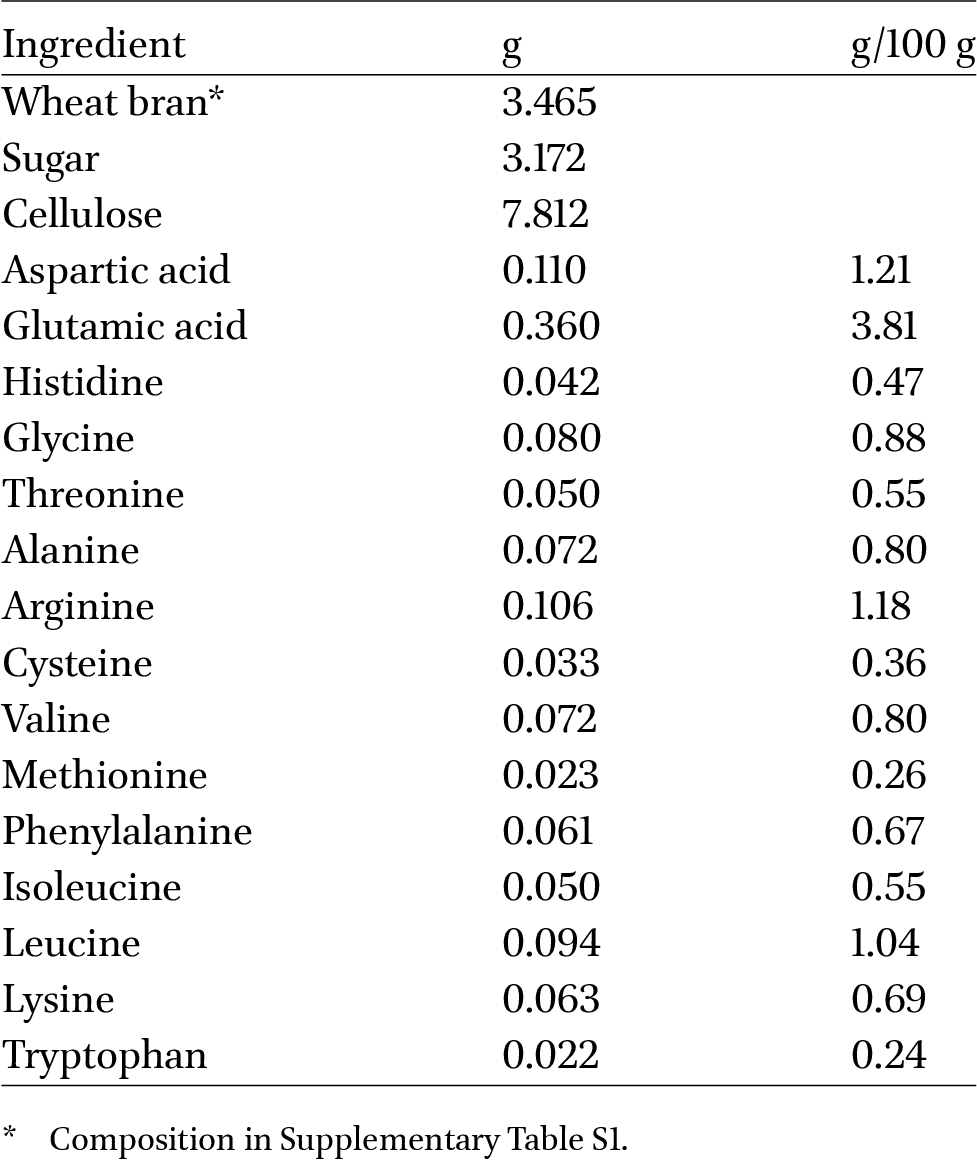
semi-artificial diet, ingredient list and amino acid content in g/100 g diet, simulating energy level and amino acid composition of a 100% wheat bran diet, for 100 mealworm larvae
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Semi-artificial diet, ingredient list and amino acid content in g/100 g diet, simulating energy level and amino acid composition of a 100% chicken feed diet, for 100 BSF larvae
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
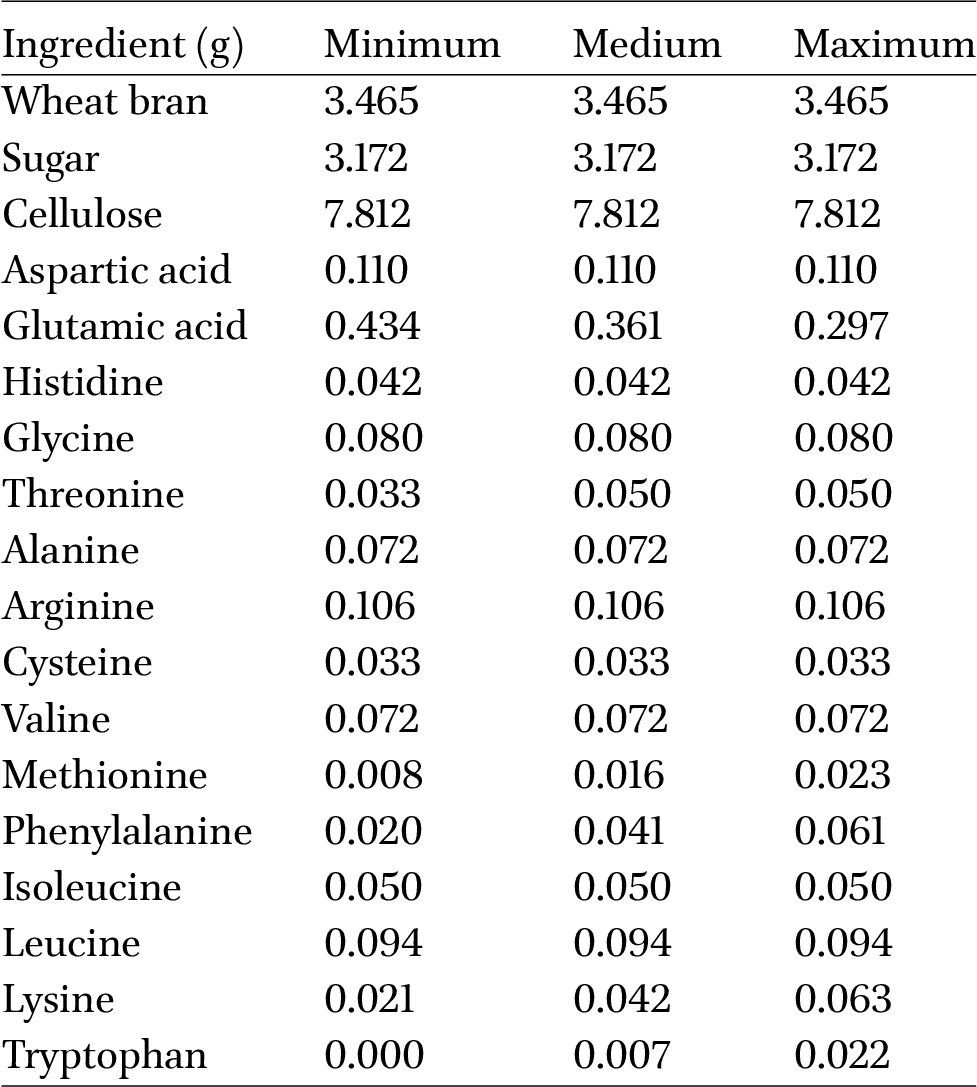
Verification diets with added levels of amino acids for 100 mealworms
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Verification diets with added levels of amino acids for 100 BSF larvae
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Semi-artificial diets based on wheat bran for mealworms (Table 1) and a mixture of chicken feed/water (30/70) for BSF (Table 2), supplemented with sugar (i.e. table sugar = sucrose) and synthetic amino acids (Sigma-Aldrich, Merck, Hoeilaart, Belgium) were tested. An analytical balance (VWR PBA224l-1S, precise up to 0.0001 g) and laboratory micro spatula were used in order to be able of accurate weighing and distributing of the small AA doses. The diets were isoenergetic and isoproteinous. Everything was provided at the start of the experiment and doses were based on downsizing practical circumstances (Coudron et al., 2019). Among the 5 studied amino acids, one was administered in varying doses for each experiment. These were 25, 50, 75 and 100% of what was present in the basic substrates of wheat bran and chicken feed. Non-essential glutamic acid was used as a substitute at lower doses, in order to be isoenergetic and isoproteinous. Growth, as measured by weight gain, and survival were observed at 27 ± 1 °C and 60 ± 5% relative humidity. For mealworms, 4-5 week-old larvae were reared on the experimental diets until reaching maximal biomass. Preliminary tests showed that following up until pupation was not optimal, given the limited space in the experimental units. For BSF, the study began with 6-7-day-old larvae and continued until the first pre-pupae developed. Three repetitions per diet and 100 larvae per repetition, kept in polypropylene cups with bottom surface of 38.5 cm2 and 10 cm high, were used for both species. All feed was provided at the outset, with moisture provided when necessary, and samples of 10 larvae were taken (and put back) every 3 days to monitor growth. In the case of mealworms, cellulose (indigestible according to Barragan-Fonseca et al., 2021) was added to ensure a sufficient volume for larvae to burrow beneath the surface. At harvest, larvae were separated from the substrate, counted and weighed. Hence, survival and yielded weight were recorded.
Since only one amino acid was adjusted at a time, the interactions between these amino acids are not yet fully understood. It is possible that setting all these amino acids simultaneously to their minimum levels could reveal certain limiting deficiencies. To investigate this, a verification test was conducted where all amino acids were simultaneously set to their minimum levels necessary for optimal growth as determined in the first experiments. This treatment was then compared with a 100% diet and an intermediate value, aiming to identify any potential new optima (Tables 3 and 4).
Concerning mealworms, there are alternatives to wheat bran available on the market that contribute to an additional 10% growth. The essential amino acid contents in these feeds generally surpass those of wheat bran. Therefore, for some amino acids, based on the outcome of the first experiments, doses of 125% and 150% were also tested.
It’s crucial to note that nutrient levels hold significance only when the feeding amounts are known. Our examinations of pure wheat bran yielded a feed conversion ratio (FCR) of 1.9. For commercial feeds, this ratio can even decrease to 1.6 (Coudron and Deruytter, 2019). For BSF reared on chicken feed the FCR under optimal conditions is 1.2 (Spranghers et al., 2019). From these findings, the minimum amounts of amino acids required to achieve a growth of 100 g can be calculated by multiplying the FCR with the requirements.
Yield expressed as harvest weight of all larvae (g) and survival (%) data were analysed using SPSS 28 (IBM Corp.). Given the nature of the data (limited amount of replicates) normality could not be tested and in order to assure sufficient power, non-parametric Kruskall-Wallis tests were used to compare the means. When differences were detected, pairwise Mann-Whiney U tests were applied to identify. P-values below 0.05 were considered statistically significant.
3 Results
Tenebrio molitor
For mealworms, survival consistently ranged between 90-95% across all treatments, with no distinction between them. Hence, figures for mortality are not being presented. Furthermore, all experimental (i.e. semi-artificial) treatments showed a similar trend in terms of development time (Figure 1). They required an additional week to develop compared to the control (i.e. pure wheat bran) group (42 and 35 days resp.). Additionally, the harvest weight of the control group was approximately 40% higher than that of the 100% treatment, which should have been equivalent. However, this equivalence is solely with respect to amino acids and does not account for potential deficiencies in micronutrients such as vitamins and minerals. Moreover, an unavoidable difference existed in the structures of the substrates, measured with a Bostwick consistometer (Diantom et al., 2017), which significantly influenced these larvae living within their feed environment.
Growth of Tenebrio molitor larvae reared on experimental (i.e. semi-artificial) diets compared to the control of pure wheat bran (25% contains 25% lysine of what is present in the control).
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Harvest weights of T. molitor larvae reared on diets with different lysine (A), methionine (B), threonine (C), phenylalanine (D) and tryptophan (E) content and with all 5 amino acids (F) at the determined minimal (minimum) and intermediate (medium) percentages based on the previous experiments and 100% (maximum); percentages of what is present in wheat bran; means ± stdev; different letters indicate statistical significant differences.
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Regarding lysine (Figure 2A), a significant reduction in harvest weight was observed at 25% of the control concentration (H = 8.231, df = 3,
Based on the experiments with methionine and threonine (Figure 2B,C), it was hypothesized that increasing their levels above those of the control might further improve yield. However, this hypothesis was not supported by the follow-up experiments (Figure 3A,B).
Extension of methionine (A) and threonine (B) levels to higher doses than the control for T. molitor larvae, percentages of what is present in wheat bran; means ± stdev.
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Hermetia illucens
For BSF, in contrast to mealworms, differences in survival were observed (Figure 4). The development time of the larvae in all experimental treatments was 23 days, which was about 14 days longer than on a 100% chicken feed diet.
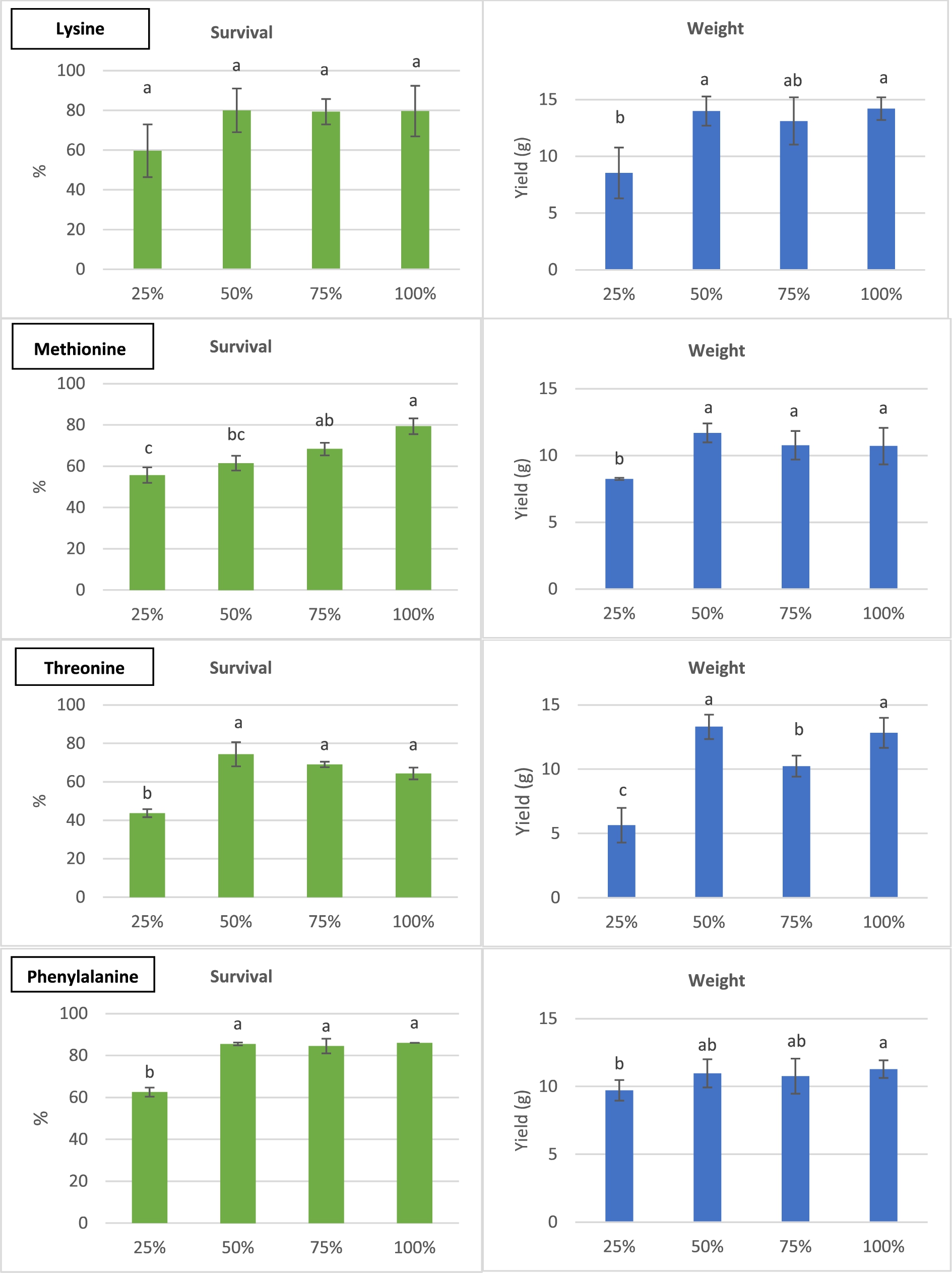
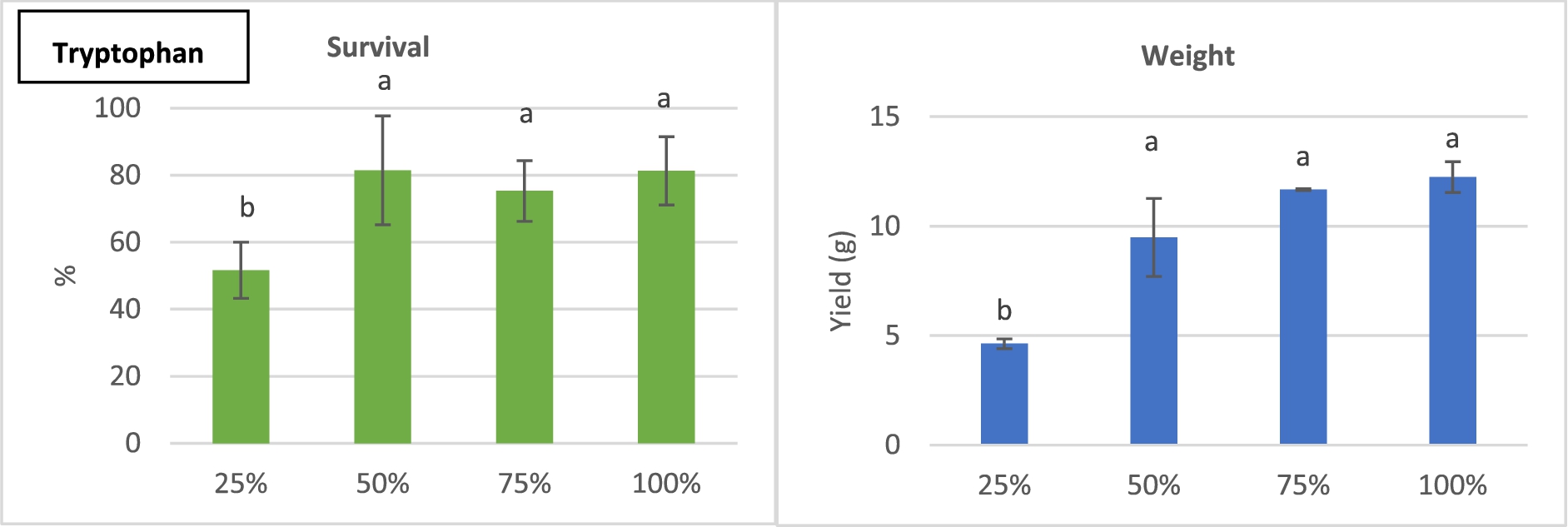
Survival (l.) and harvest weights (r.) of H. illucens larvae reared on diets with different lysine (A), methione (B), threonine (C), phenylalanine (D) and tryptophan (E) contents, percentages of what is present in chicken feed; means ± stdev; different letters indicate statistical significant differences.
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
(Continued.)
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
Despite the considerable standard deviations at 25% and 75% of the control, it could be deduced that 50% lysine (0.53 g/100 g) was already optimal (H = 4.727, df = 3,
There were no observed differences between diets with all 5 amino acids at their minimum required and maximum levels (Figure 5). Analogue to what is projected in Figure 1 for mealworms, a control diet made out of 100% chicken feed resulted in heavier larvae than an isoenergetic and isoproteinous semi-artificial diet containing sugar and synthetic amino acids (H = 9.419, df = 3,
Harvest weights of H. illucens larvae reared on diets with all 5 amino acids at the determined minimal (minimum) and intermediate (medium) percentages based on the previous experiments and 100% (maximum) of what is present in a chicken feed diet compared to a pure chicken feed diet (CTR).
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
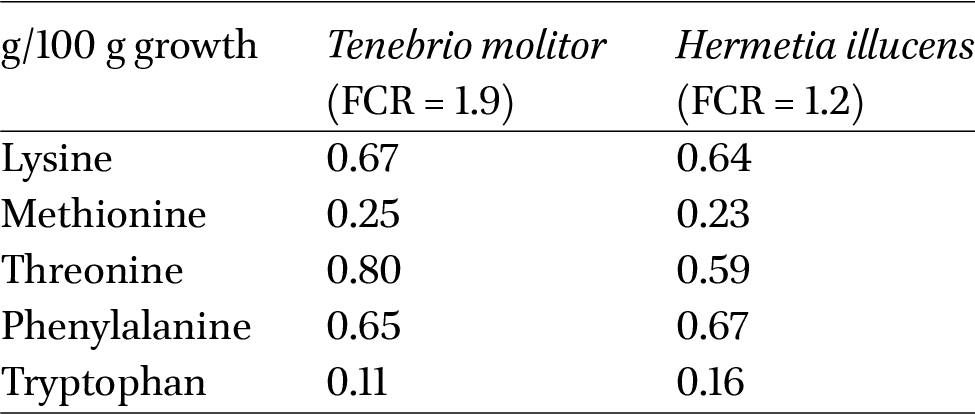
Amino acid uptake (in g) necessary per 100 g of weight gain (as is) of Tenebrio molitor and BSF larvae
Citation: Journal of Insects as Food and Feed 11, 6 (2025) ; 10.1163/23524588-00001271
From Table 5 we can see that the amounts of amino acids necessary for gaining 100 g mealworm and BSF larvae are similar for lysine, methionine and phenylalanine. However, for threonine mealworms require more whereas tryptophan requirements are lower compared to BSF larvae.
4 Discussion
From these experiments, it can be concluded that the commonly used substrates (wheat bran for mealworms and chicken feed for BSF) contain ample essential amino acids. Regarding mealworms, the added value of market-available feeds (which are largely based on wheat bran) is situated more in other micronutrients such as vitamins and minerals. For mealworms, an excess of amino acids, and hence proteins, is not of major concern since dry feeds are utilized. However, in the case of BSF, harmful ammonia can be produced, which could be the case when chicken feed is provided with water (Coudron et al., 2021).
Few studies on insects have been conducted and therefore a comparison with other arthropod species can be made (which is better than comparison with vertebrates like the current farm animals) (Ottoboni et al., 2017). Shrimp require also the usual 10 essential amino acids (EAA): arginine, methionine, valine, threonine, isoleucine, leucine, lysine, histidine, phenylalanine and tryptophan. The first limiting amino acids when applying least-cost formulation of commercial shrimp feed are generally lysine, methionine and arginine. The nonessential cysteine can have a sparing effect for methionine whereas tyrosine could spare for phenylalanine (Fox et al., 2019). As protein nutritive value is based on its amino acid composition, it is also advisable that shrimp feeds contain a similar amino acid profile to that of the body composition of the shrimp species (Deshimaru and Shigeno, 1972; Wilson and Poe, 1985; Kanazawa, 1989). This is analogue to the findings of John et al. (1978, 1979) for mealworms. Studies about the fruit fly Drosophila melanogaster show that dietary restriction through dilution of casein may extend adult longevity, at the cost of lower fecundity, because of the limited intake of amino acids (Min and Tatar, 2006). However, it may be possible to obtain the benefits to longevity of dietary restriction without causing a fecundity reduction, through a suitable balance of amino acids in the diet (Grandison et al., 2009). Taylor and Medici (1966) also developed a pure diet with respect to amino acids to determine amino acid requirements of several grain beetles. The confused flour beetle, Tribolium confusum, required (in % of diet) 0.56% lysine; 0.3% methionine; 0.7% phenylalanine; 0.4% threonine and 0.1% tryptophan. These levels are comparable to our findings for mealworms, with the exception of threonine (Table 5).
According to Koethe et al. (2022), 0.30 g lysine/100 g diet should already be sufficient for optimal growth of BSF larvae. However, this is not coming out of the present study as 0.27 g clearly resulted in less yield. The importance of phenylalanine aligns with the needs observed in other insects, such as honeybees (Apis mellifera). It is also the most phagostimulant amino acid (i.e. enhances the food intake) and is often the amino acid with the highest concentration in nectar of plants visited by the bees (Wright et al., 2018). Tryptophan regulates Drosophila melanogaster zinc body stores (Garay et al., 2022). Given that BSF larvae tend to accumulate this heavy metal (Lievens et al., 2021), it is advisable to not overprovide tryptophan. As shown by the present results, BSF larvae only need to take up 0.16 g in order to gain 100 g of body weight. In contrary, it is shown that methionine is a major limiting factor for fecundity of the fruit flies (Hoedjes et al., 2017). This might also be interesting for adult BSF, however, this was out of the scope of this research.
Considering the protein requirements it is mandatory to take in consideration the protein biological value. Typically, protein biological value is defined as the nitrogen used for tissue formation divided by the nitrogen absorbed from a giving ingredient/diet. This product is multiplied by 100 and expressed as a percentage of nitrogen utilized. The biological value provides a measurement of how efficient a giving animal utilizes protein consumed in the diet. It means that the protein quality of any food is a measure of its ability to provide all the amino acids required for growth and maintenance of body functions and the digestibility of the protein. A further aspect to take in consideration when selecting insect substrate on protein content is related to NPN an analytical method for protein quantification. Indeed, the protein content in vegetable matrix analyses is based on nitrogen content using the nitrogen-to-protein conversion factor (Kp) of 6.25 generally used for proteins (European Commission, 2009). The presence of nonprotein nitrogen (NPN), for example chitin, nucleic acids, phospholipids, and ammonia could lead to an overestimation of the protein content. Thus, the evaluation of potential bias in protein quantification (NPN correction) together with the estimation of biological value are key aspect to guarantee an optimal utilization of protein nitrogen contained in the substrate to meet, in turn, insect protein dietary requirements. In addition, for BSF larvae a correlation between the sum of proteins with non-fibre carbohydrates (NFCs) in the substrate and the harvested larval biomass (yield) was revealed. If we assume that, as found by Coudron et al., 2019, 10-15 g protein/100 g dry matter is sufficient, then this substrate should contain a minimum of 35-40 g NFCs/100 g dry matter to achieve an adequate yield. This connection highlights the interplay between substrate composition and larval growth and can guide the formulation of optimal diets for insect production (Coudron et al., 2019).
To achieve optimal growth in livestock, a different ratio of essential amino acids in the feed is necessary at each stage of their lives. Since amino acids are very important for growth, young animals often require different and more amino acids in their feed than older animals. Arginine is generally considered essential for newborn piglets and chicks because the metabolic routes that synthesize this amino acid are not yet fully developed in newborns (Lin, 2007). Arginine is the most abundant nitrogen carrier in tissue proteins and is therefore important for maximum growth of young mammals. Formation of nitric oxide from arginine can regulate inflammation and exogenous sources of arginine can help increase the proliferation of monocytes and lymphocytes and the formation of T-helper cells (Suchner et al., 2002). In addition to arginine, histidine is also an essential amino acid for piglets. It is a precursor of various hormones and critical metabolites that affect kidney function, neurotransmission, gastric secretion, and the immune system (Lin, 2007).
Besides absolute content of essential amino acids, protein quality of substrates is also determined by digestive physiology (i.e. digestibility), interaction with microbiota and feeding behaviour. The latter being especially important for insects living in their food substrate like mealworms and BSF larvae. Future research needs to investigate aspects influencing feeding behaviour, digestive physiology, morphology and function and establish parameters for monitoring larval health and development during mass rearing on specific substrates, and describe in vitro digestion models capable of predicting outcomes from in vivo feeding experiments (Guillaume et al., 2023; Moutinho et al., 2023).
The observed differences between larvae reared on pure diets and those reared on semi-artificial diets, despite being isoenergetic and isoproteinous, highlights the need for other micronutrients such as certain minerals and vitamins. A deficiency in micronutrients can significantly impact the body’s ability to respond to changes in dietary amino acid intake. This is because of their role in various functions like protein synthesis, amino acid metabolism, immune function, absorption and transport (Berger et al., 2022). In essence, a deficiency in micronutrients can create a cascade of effects that can mask or exacerbate the symptoms of amino acid deficiency. Therefore, when studying the effects of removing amino acids from a diet in the future, it is essential to consider the micronutrient status. In addition, also the physical structure of the diet plays an important role when comparing pure and semi-artificial diets, given that these larvae live in their food.
5 Conclusion
In conclusion, the experiments provide valuable insights into the nutritional requirements of mealworms and BSF larvae in relation to essential amino acids. The study emphasizes that commonly used substrates for these insects, such as wheat bran for mealworms and chicken feed for BSF, contain ample essential amino acids. However, it distinguishes the added value of market-available feeds for mealworms, where micronutrients like vitamins and minerals play a more significant role than excess amino acids. In addition, improvement of growth performance through amino acid provision may be possible for mealworms before the age of 5 weeks or BSF larvae during the first week nursery phase. This was not addressed by this study nor the effects of other amino acids besides lysine, methionine, threonine, phenylalanine and tryptophan. In contrast, the potential harm of ammonia production in BSF, provided slurry diets, emphasizes the importance of amino acid balance. Research indicates that lower protein content in BSF substrates is feasible, with optimal results achieved with 10-15% crude protein, depending on the quality.
To our knowledge, this study is the first to investigate the requirements of 5 essential amino acids for 2 species of production insects. The results provide a basis for insect rearing companies to formulate their diets with regards to optimal nutrition. Verification on actual side streams is necessary to further develop nutritional guidelines and construct feed tables like those that already exist for other farm animals (INRA CIRAD AFZ, 2021). In addition, there is the need for future research to explore aspects like feeding behaviour, digestive physiology, and larval health to optimize insect rearing on specific substrates.
Corresponding author; e-mail: thomas.spranghers@vives.be
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.27248121
Acknowledgements
This research was funded by the PWO project: Insecten van A tot Z (iAZ) of VIVES University of Applied Sciences. This project was in close cooperation with the industry and the authors would like to thank Carl Coudron (Inagro), Alexander Maroy (Nusect), Geert Poels (VDL Agrotech), Bas Artz (Duynie Feed), Dirk Coremans (Duynie Ingredients), Jeroen Geuens and Annelore Wens (KdG University of Applied Sciences and Arts), Frank Hartmann (Roxell), Meggie Van Peer (Thomas More University of Applied Sciences), Tom Douven and Tim van Heertum (VDL Insect Systems) and Kristof Das (Agropolis) for their valuable input as steering committee members.
Conflict of interest
The authors have no conflict of interest to declare.
References
Barragan-Fonseca, K.B., Gort, G., Dicke, M. and Van Loon, J.J.A., 2021. Nutritional plasticity of the black soldier fly (Hermetia illucens) in response to artificial diets varying in protein and carbohydrate concentrations. Journal of Insects as Food and Feed 7: 51-61. https://doi.org/10.1093/jfaf/iyab001
Berger, M.M., Shenkin, A., Schweinlin, A., Amrein, K., Augsburger, M., Biesalski, H.K., Bischoff, S.C., Casaer, M.P., Gundogan, K., Lepp, H.L., de Man, A.M.E., Muscogiuri, G., Pietka, M., Pironi, L., Rezzi, S. and Cuerda, C., 2022. ESPEN micronutrient guideline. Clinical Nutrition 41: 1357-1424. https://doi.org/10.1016/j.clnu.2022.02.015
Chang, C.L., 2004. Effect of amino acids on larvae and adults of Ceratitis capitata, Diptera: Tephritidae. Annals of the Entomological Society of America 97: 529-535. https://doi.org/10.1603/0013-8011(2004)097[0529:EOAOLA]2.0.CO;2
Coudron, C., Spranghers, T., Elliot, D. and Halstead, J., 2019. Insect breeding and requirements. BioBoost. Available at: https://www.bioboost-platform.com/documents/lab-scale-and-pilot-scale-experiments-with-mealworm-and-black-soldier-fly/
Coudron, C., Deruytter, D., Van Peer, M., Spranghers, T., Van Miert, S. and Claeys, J., 2021. Chapter 4: Production practices. In: Veldkamp, T., Claeys, J., Haenen, O.L.M., Van Loon, J.J.A. and Spranghers, T. (eds.) The basics of edible insect rearing: handbook for the production chain. Wageningen Academic Publishers, Wageningen, the Netherlands. https://doi.org/10.3920/978-90-8686-902-2
Davis, G.R.F., 1975. Essential dietary amino acids for growth of larvae of the yellow mealworm, Tenebrio molitor L. The Journal of Nutrition 105: 1071-1075. https://doi.org/10.1093/jn/105.8.1071
Deshimaru, O. and Shigeno, K., 1972. Introduction to the artificial diet for prawn Penaeus japonicus. Aquaculture 1: 115-133. https://doi.org/10.1016/0044-8486(72)90002-2
Diantom, A., Curti, E., Carini, E. and Vittadini, E., 2017. Effect of added ingredients on water status and physico-chemical properties of tomato sauce. Food chemistry 236: 101-108. https://doi.org/10.1016/j.foodchem.2017.05.038
European Commission, 2009. Regulation, EC No 152/2009 of the European Parliament and of the Council of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Official Journal of the European Union L 54/1-130.
Fox, J.M., Davis, D.A., Wilson, M. and Lawrence, A.L., 2019. Current status of amino acid requirement research with marine penaeid shrimp. Avances en Nutrición Acuı́cola. Available at: https://nutricionacuicola.uanl.mx/index.php/acu/article/view/168
Garay, E., Schuth, N., Barbanente, A., Tejeda-Guzmán, C., Vitone, D., Osorio, B., Clark, A.H., Nachtegaal, M., Haumann, M., Dau, H., Vela, A., Arnesano, F., Quintanar, L. and Missirlis, F., 2022. Tryptophan regulates Drosophila zinc stores. Proceedings of the National Academy of Sciences 119: e2117807119. https://doi.org/10.1073/pnas.2117807119
Grandison, R.C., Piper, M.D. and Partridge, L., 2009. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 462: 1061-1064. https://doi.org/10.1038/nature08619
Guillaume, J.B., Mezdour, S., Marion-Poll, F., Terrol, C. and Schmidely, P., 2023. Asymptotic estimated digestibility, a new indicator of black soldier fly (Hermetia illucens) conversion efficiency in relation to larval density. Journal of Insects as Food and Feed 9: 893-906. https://doi.org/10.1080/21658005.2023.1973965
Hoedjes, K.M., Rodrigues, M.A. and Flatt, T., 2017. Amino acid modulation of lifespan and reproduction in Drosophila. Current Opinion in Insect Science 23: 118-122. https://doi.org/10.1016/j.cois.2017.08.005
INRAE CIRAD AFZ., 2021. Composition and nutritive values of feeds for cattle, sheep, goats, pigs, poultry, rabbits, horses, and salmonids. Feedtables. Available at: https://www.feedtables.com/
John, A.M., Davis, G.R.F. and Sosulski, F.W., 1978. Protein nutrition of Tenebrio molitor L. XIX. Growth response to levels of dietary protein and of an amino acid mixture, 1. Archives Internationales de Physiologie et de Biochimie 86: 761-770. https://doi.org/10.3109/13813457809069528
John, A.M., Davis, G.R.F. and Sosulski, F.W., 1979. Protein nutrition of Tenebrio molitor L. XX. Growth response of larvae to graded levels of amino acids. Archives Internationales de Physiologie et de Biochimie 87: 997-1004. https://doi.org/10.3109/13813457909070548
Kanazawa, A., 1989. Protein requirements of Penaeid shrimp. In: Advances in Tropical Aquaculture, Workshop at Tahiti, French Polynesia, 20 Feb - 4 Mar 1989. Available at: https://archimer.ifremer.fr/doc/00000/1485/
Koethe, M., Taubert, J. and Vervuert, I., 2022. Impact of lysine supplementation on growth and development of Hermetia illucens larvae. Journal of Insects as Food and Feed 8: 35-44. https://doi.org/10.1080/21658005.2021.1973965
Lievens, S., Poma, G., De Smet, J., Van Campenhout, L., Covaci, A. and Van Der Borght, M., 2021. Chemical safety of black soldier fly (Hermetia illucens): knowledge gaps and recommendations for future research: a critical review. Journal of Insects as Food and Feed 7: 383-396. https://doi.org/10.1080/21658005.2021.1973965
Lin, C., 2007. Amino acid nutrition for growth and health of neonatal pigs, PhD thesis, Texas Tech University. Available at: https://ttu-ir.tdl.org/items/0d19790a-b595-4482-816e-40f130980d45
Min, K.J. and Tatar, M., 2006. Restriction of amino acids extends lifespan in Drosophila melanogaster. Mechanisms of Ageing and Sevelopment 127: 643-646. https://doi.org/10.1016/j.mad.2006.02.006
Moutinho, S., Peres, H., Martins, N., Serra, C., Santos, R.A., Monroig, Ó. and Oliva-Teles, A., 2023. Use of black soldier fly (Hermetia illucens) larvae meal in diets for gilthead seabream juveniles: effects on growth-related gene expression, intermediary metabolism, digestive enzymes, and gut microbiota modulation. Aquaculture 580: 740357. https://doi.org/10.1016/j.aquaculture.2023.740357
Ottoboni, M., Tretola, M., Cheli, F., Marchis, D., Veys, P., Baeten, V. and Pinotti, L., 2017. Light microscopy with differential staining techniques for the characterization and discrimination of insects versus marine arthropods processed animal proteins. Food Additives and Contaminants: Part A 34: 1377-1383. https://doi.org/10.1080/19440049.2017.1345075
Spranghers, T., Schillewaert, S. and Wouters, F., 2019. Optimizing nutrition of black soldier fly larvae. 70th Annual Meeting of the European Federation of Animal Science (EAAP), Ghent, 26-30 August. Available at: https://docs.eaap.org/2019/theater/16/1400_Spranghers.pdf
Suchner, U., Heyland, D.K. and Peter, K., 2002. Immune-modulatory actions of arginine in the critically ill. British Journal of Nutrition 87: S121-S132. https://doi.org/10.1079/BJN2001457
Taylor, M.W. and Medici, J.C., 1966. Amino acid requirements of grain beetles. The Journal of Nutrition 88: 176-180. https://doi.org/10.1093/jn/88.2.176
van Milgen, J. and Dourmad, J.Y., 2015. Concept and application of ideal protein for pigs. Journal of Animal Science and Biotechnology 6: 1-11. https://doi.org/10.1186/s40104-015-0004-1
Wilson, R.P. and Poe, W.E., 1985. Apparent digestible protein and energy coefficients of common feed ingredients for channel catfish. The Progressive Fish-Culturist 47: 154-158. https://doi.org/10.1577/1548-8640(1985)47<154:ADPAEC>2.0.CO;2
Wright, G.A., Nicolson, S.W. and Shafir, S., 2018. Nutritional physiology and ecology of honey bees. Annual Review of Entomology 63: 327-344. https://doi.org/10.1146/annurev-ento-020117-043348
Wu, G., Bazer, F.W., Dai, Z., Li, D., Wang, J. and Wu, Z., 2014. Amino acid nutrition in animals: protein synthesis and beyond. Annual Review of Animal Biosciences 2: 387-417. https://doi.org/10.1146/annurev-animal-022513-114113







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}