Abstract
The sustainability of aquaculture production is crucial for the industryâs long-term viability. Diversification of feed ingredients is a key component in achieving this sustainability. Considered a promising alternative to traditional feed ingredients, terrestrial insect meals often lack sufficient n-3 polyunsaturated fatty acids (PUFA). This study explores the potential of marine insect species, specifically evaluating the influence of five different substrates on the growth and composition of the European native seaweed fly, Fucellia maritima. The substrates tested include a brown seaweed (Fucus sp.), a green seaweed (Ulva sp.), a red seaweed (the invasive Agarophyton vermiculophyllum), the invasive freshwater hyacinth (Eichhornia crassipes), and fish processing waste (codfish Gadus morhua frames). Results show no significant difference in the number of individuals per substrate, although Fucus sp. had a higher total number of individuals completing their life cycle. In contrast, feeding on codfish frames resulted in a lower number of completed life cycles, while E. crassipes led to no survival. Protein content in adult flies ranged from 55.2% to 56.7%, and in pupae, it ranged from 46.1% to 48.7% across different feeding substrates. Notably, pupae fed with Ulva sp., exhibited the highest protein content at 48.7%. Lipid content in adult flies ranged from 10.0% to 13.0%, while in pupae, it ranged from 8.8% to 11.4%. Codfish frames had the highest lipid content in both pupae (11.4%) and adults (13%). The most abundant fatty acid in Fucus sp. â fed pupae and adult flies was oleic acid (18:1 n-9), while palmitoleic acid (16:1 n-7) dominated in other treatments. It is worth highlighting that docosahexaenoic acid (22:6 n-3) was reported for the first time in a seaweed fly, namely when supplied with fish processing waste. These findings suggest that Fucellia maritima may be a promising complementary ingredient for formulating marine aquafeeds.
1 Introduction
As estimates point to a global population of 9.8 billion people by 2050, concerns about producing enough food will also increase (UN, 2022). Current estimates suggest that we need to increase food production by more than 50% to sustainably meet the demand of this expanding population for wholesome food (van Dijk et al., 2021). Aquaculture is an important food production system that can contribute to many countriesâ food security (FAO et al., 2022). In 2022, aquaculture surpassed fisheries for the first time in the annual production of aquatic animals, accounting for 51% of the total production worldwide (FAO, 2024).
Marine carnivorous fish and shrimp production requires a diet high in animal protein and certain lipids (FAO, 2022; Glencross, 2009). To meet these nutritional requirements, two ingredients are commonly used in aquafeed formulation: fishmeal and fish oil; the latter is rich in long-chain n-3 polyunsaturated fatty acids (LC-PUFA) (FAO, 2020). The need for these two ingredients has caused great pressure on fisheries stocks, encouraging the search for more sustainable alternative ingredients, such as plants (not so environmentally sustainable and may contain anti-nutritional factors), single-cell proteins (expensive and not easily obtainable), livestock sources (not environmentally friendly and a potential vector for diseases) and insect meals (terrestrial insects are poor on n-3 polyunsaturated fatty acids, PUFA) (Ameixa et al., 2020).
Despite insects being the most diverse group of animals on the planet, with nearly one million species described (BaÌnki et al., 2024), only eight species are authorised to be used as animal feed in the European Union. In 2017, the European Commission (2017) authorised the use of seven insect species as aquafeed ingredients, namely, two species of Diptera, the black soldier fly (Hermetia illucens, BSF) and the common housefly (Musca domestica, CHF), two species of Coleoptera, the yellow mealworm (Tenebrio molitor, YM) and the lesser mealworm (Alphitobius diaperinus), and three species of Orthoptera, the house cricket (Acheta domesticus), the banded cricket (Gryllodes sigillatus), and the field cricket (Gryllus assimilis). In 2021, another species was added to this list, the Lepidoptera silkworm (Bombyx mori) (EU Regulation 2021/1925, 2021). This authorisation was extended to other farmed animal species in 2021, including poultry, swine, pets (e.g. dogs, cats, birds, or reptiles), and fur animals (e.g. Mustela lutreola) with the Regulation (EU) 2021/1372 of 17 August 2021 (EU, 2021). In 2024, the European Commission authorised the use of live insects in animal feed in the European Union. This includes feed for farmed fish, poultry, pigs, and pets, excluding ruminant animals.
Despite the knowledge gaps that remain when it comes to up-scaling the use of these eight species, the list is comparatively small compared to the estimated insect diversity. In fact, due to their particular life histories, several species may present characteristics that enrich the diets of farmed animals. For instance, evidence shows that insects that live in coastal areas and feed on organic matter of marine origin present a nutritional composition richer in n-3 (Duarte et al., 2021). For this reason, they can potentially be good candidates as aquafeed ingredients destined for marine fish. One example is the seaweed flies, which are known to feed on seaweed and dead fish in coastal areas (Biancarosa et al., 2018b; Cheng, 1976).

Fucellia maritima adult fly on the sand at Barra beach, Aveiro, Portugal (40°38â31.1âN; 8°44â57.7âW).
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
The seaweed fly Fucellia maritima (Haliday, 1838) (Figure 1), native to western and northern Europe, is a member of the family Anthomyiidae and can be seen throughout the year overflying the beach wrack along the shores of Portugal (Lourenço et al., 2020). It dwells on the beach wrack, predominantly composed of brown seaweed, mostly from the genera Laminaria and Fucus. It is also the only seaweed fly species known to date that can feed on decaying animal matter (Egglishaw, 1960). This species has a relatively short life cycle of approximately 36 days at room temperature (15-18 °C), with three larval instar and pupae phases (Egglishaw, 1960). In captivity, this species is described as reproducing on moderately decomposed brown seaweed Laminaria sp. and Fucus sp. with adult flies freely mating and laying their eggs on the seaweed (Egglishaw, 1960). Like many others, this species is understudied, and the potential to be used as a feed ingredient is unknown. Hence, it is not authorised for feed production in the EU.
In the Centre region of mainland Portugal, namely in the coastal areas of the Aveiro region, beach wrack is predominantly composed of a mixture of Fucus sp. and Ulva sp., but at certain times of the year, large amounts of the invasive aquatic plant, the water hyacinth (Eichhornia crassipes), can also be observed (O. Ameixa, pers. obs.). To the authorsâ best knowledge, no study has ever investigated the biochemical profile of F. maritima reared in captivity. Hence, information on its fatty acid (FA) profile, namely in PUFA on different substrates, has yet to be determined in both pupae and adult flies.
In this work we assessed the potential of marine insect species, through the evaluation of the influence of five different substrates on the growth (biomass) and nutritional composition of the European native seaweed fly, Fucellia maritima. For this assessment we evaluated the nutritional composition of F. maritima through the use of feeding substrates commonly found in the composition of beach wrack on the coast of Aveiro region, namely: (a) the brown seaweed Fucus sp. which was used as the control group (CG), due to the fact that this is the preferential feeding substrate for this fly species; (b) the green seaweed Ulva sp.; (c) the invasive red seaweed Agarophyton vermiculophyllum; (d) the invasive freshwater plant, the water hyacinth (Eichhornia crassipes), which can be very abundant in the local ecological niche of the fly; and (e) frames of Atlantic codfish (Gadus morhua, a common co-product of local codfish salting industries). Whole seaweeds are listed as authorised feed ingredients in the European catalogue of feed materials (Reg. EU 68/2013) (European Commission, 2013). However, the use of whole codfish frames is currently not authorised as a substrate for insects in the EU (European Parliament and Council, 2009).
2 Material and methods
Substrates sampling and preparation
The brown seaweed Fucus sp. and the green seaweed Ulva sp. were sampled at Ria de Aveiro coastal lagoon in April 2021 (40°38â04.4âN; 8°39â40.2âW), while the invasive red seaweed A. vermiculophyllum was harvested in July 2020 in the same location. The invasive water hyacinth E. crassipes was sampled at the mouth of the Vouga River, also in Ria de Aveiro, in April 2021 (40°41â41.6âN; 8°36â06.8âW), with whole thalli being collected in plastic buckets along with some water to maintain the conditions during transport. In the laboratory at ECOMARE (University of Aveiro, Portugal), on the same day of sampling, all seaweed and water hyacinth were rinsed in cold fresh water to remove salt, debris, and other algal or animal species present and were subsequently stored in small plastic bags at â20 °C for further use in the experiment. Frames of Atlantic codfish (G. morhua) were supplied by an industrial codfish-salting unit (Bacalhau do Barents, Produtos Alimentares Lda, Gafanha da NazareÌ, Portugal) and stored at â20 °C until further use.
The feeding substrates being tested were later defrosted and minced using a 200Â W Selecline Hand Blender to increase the surface-to-area ratio and facilitate their consumption by the larvae. The exception was codfish frames, which were cut into small pieces using scissors. Two hundred grammes of each substrate were placed into 1.1Â L plastic food containers with a mesh net in the lid for air circulation.
Insect collection
Adult specimens of F. maritima overflying the beach wrack were captured using a sweeping net at Barra beach, Aveiro, Portugal, in April 2021 (40°37â44.3âN; 8°44â42.0âW). Collected specimens were stored in 50-mL polypropylene flasks for transportation to the insectarium facility at ECOMARE (University of Aveiro, Portugal) and kept under controlled conditions (a photoperiod of 16 h L:8 h D, at 40 ± 5% Relative Humidity and 25 ± 3 °C temperature).
Experimental design
In the insectarium, adult flies were transferred to plastic boxes of 1.1 L containing the feeding substrates being tested (as described above). Fifty adult flies were placed in each box, with the number of males and females unknown since it is impossible to distinguish sexes without dissecting this species. Five independent replicates were tested for each substrate, with 50 flies à 5 feeding substrates à 5 replicates = 1,250 adult flies being used. The adult flies were allowed to freely mate and lay their eggs on the substrate for 48 hours, and the eggs deposited were not counted. The containers were supplied with tap water daily to maintain humidity, especially when supplying seaweed, as desiccation induced larval mortality. Within 4 days, larvae started to moult into pupae, and half of the pupae were sampled and frozen at â20 °C. After one week, adult flies emerged from the remaining puparium and were frozen at â20 °C until further analysis. Therefore, the experiment stopped when the adult flies emerged from the remaining puparium.
Sample processing
All frozen material was transported to the Instituto de Investigaciones Marinas (IIM-CSIC, Vigo, Spain), where it was flash-frozen at â80 °C before being freeze-dried. After freeze drying, all samples, except for codfish frames (see below), were ground, with a porcelain mortar and pestle being used to homogenise samples of pupae and adults of F. maritima and a blender (La Moulinette 700 W from Moulinex) being used for seaweed and freshwater hyacinth. Skin and meat from codfish frames were separated from the bone and minced using a blender (Moulinette Compact mini hachoir 270 W from Moulinex).
Nutrient content analysis
Due to the low biomass of the initial five sample replicates per treatment, these were mixed and re-distributed into two replicates per treatment to analyse nutrient content. Moreover, it was impossible to repeat the experiment to increase the biomass necessary for the analysis due to the start of the cleaning period at the beaches in Portugal, which happens yearly during spring to autumn, which removes the beach wrack from the beaches, affecting the local populations of seaweed flies. Therefore, we decided to follow the analysis nonetheless, due to the novelty of the work. Analytical determinations of water content (moisture), crude protein, total lipid, and crude ash were performed. The moisture content was determined by the gravimetric difference between the final weight and the weight of the sample after being dried for 24 hours at 105 °C. Crude ash was determined using the AOAC method (AOAC, 2003). Protein content was determined via the Kjeldahl method (AOAC, 2003), and total nitrogen of the substrates was multiplied by a 6.25 conversion factor. For both pupae and adult flies, a conversion factor of 4.76 was used. This value is more suitable to avoid an overestimation of insectsâ protein content (see Janssen et al., 2017). Results are expressed as dry matter.
Chemicals
Methanol (CH3OH), Chloroform (CHCL3), Dichloromethane (CH2CL2), Toluene (C7H8), Hexane HPLC (C6H14), Sulfuric Acid (H2O4S) and Petroleum Ether were purchased from Fischer Scientific (Loughborough, UK) MILLIQ Water was obtained by a purification system (Synergy, Millipore Corporation, Burlington, MA, USA). Amino acid standards mixture containing acidic, neutral, and basic amino acids were purchased from Waters (Milford, MA, USA). The AccQâTag eluent concentrates, and AccQâTag Fluor Reagent Kit derivation for hydrolysate Amino Acid Analysis were purchased from Waters Corporation (Milford, MA, USA). Tryptophane, OH-Proline, taurine, and ornithine were purchased from Merck (St. Louis, MO, USA). Acetonitrile and Water HPLC grade were purchased from Fisher Scientific.
Lipid extraction and fatty acids analysis
The lipids from the pupae, the adult flies, and the codfish were extracted using a method based on the Bligh-Dyer protocol with a proportion of CH3OH: CH2CL2 1:1 (Bligh and Dyer, 1959). Approximately 0.065 g was weighed for the adult flies, and 4 mL of CH3OH + 2 mL CH2Cl2 were added and mixed with Turrax for 60 seconds. Then, more 2 mL CH2Cl2 + 2.5 MILLIQ Water were added before mixing in Turrax again for 60 seconds. They were centrifuged at 3500 r.p.m. for 10 minutes at 4 °C (Allegra X-12R from Beckman Coulter, Brea, CA, USA). Lipids were recovered from the organic phase.
For the pupae and the codfish samples, 6Â mL of CH3OH and 3Â mL of CH2Cl2 were added and mixed for 60 seconds with Turrax. This procedure in pupae was followed by adding more 3Â mL CH2Cl2 and 4Â mL Milli-Q water. As for the codfish, 3Â mL of Milli-Q water was added because the sample was not freeze-dried; hence, less water was needed to separate the phases. For the seaweed and the water hyacinth, a modification of the method described by Lopes et al. (2019), using CH2Cl2 instead of CHCL3, was employed.
Fatty acids (FA) were analysed as Fatty Acid Methyl Esters (FAME). Briefly, lipids were methylated with 1% sulfuric acid methanolic solution by incubation at 50 °C for 18 h; after that, an aqueous 5% NaCl solution was added, and the resulting methyl esters were extracted into toluene. FAMEs were determined in duplicates in a Gas chromatograph (Perkin Elmer Clarus 500 GC, Madrid, Spain) equipped with a flame ionisation detector and a fused silica capillary column (SP-2330, 0.25 mm à 30 m à 0.2 μm, Supelco, catalogue no.: 24019). Identification of FAME was achieved using commercially available certified standard material (Supelco 37 Component FAME Mix, CRM47885), and nonadecanoic fatty acid was used as an internal standard.
Separation and quantification of lipids by Thin Layer Chromatography (TLC)
TLC plates (Silica gel on TLC plates, 20 cm à 20 cm, Merck) were prewashed with CH3OH. At the time of use, the plates were activated in an oven at 120 °C for one hour. Lipid samples at a concentration of 25 ug/uL in CH2Cl2 were injected using Hamilton TLC syringes. TLC plates were eluted in a mobile phase of hexane, ether, and acetic acid (80:20:2 by volume). The spots were visualised with a 10% copper sulphate solution in 8% phosphoric acid and heated at 160 °C for 10 minutes. Lipids were identified and quantified using a standard mixture of triglycerides, free fatty acids, monoglycerides, diglycerides, cholesterol, phospholipids, and waxes.
Amino acids analysis of the substrates for fly rearing
Amino Acids (aa) analysis was performed using the Waters procedure (AccQâTag Kit, Milford USA). Briefly, aa were extracted from defatted samples by hydrolysis with HCL 6N, 1% Phenol at 110 °C for 24 hours, and Tryptophan was determined directly by HPLC-FL after basic hydrolysis with NaOH 4N at 110 °C for 24 hours. Amino Acids from acid hydrolysis derivatization with AccQâTag reagents were prepared according to the manufacturerâs protocol. The derivatised aa and tryptophan were then analysed by HPLC coupled with a Fluorescence detector (Waters Alliance 2695 HPLC system, Multi λ Fluorescence detector 2475). The separation column was a Waters AccQâTag for Hydrolysate Amino Acid Analysis working at 37 °C. The Fluorescence detector was set at
Statistical analysis
Statistical tests were carried out to compare the influence of the type of substrate on the number of pupae and adults. Larval weight was not measured individually; rather, it was presented as a ratio of the total weight for each treatment by the number of individuals. A two-way ANOVA was performed using linear models (independent variables: substrate used with four levels, namely: Fucus sp., A. vermiculophyllum, Ulva sp., and cod fish frames). Dependent variables: life cycle stage with two levels: number of adult flies and number of pupae). Homogeneity in variances and normal distribution was verified by graphical evaluation using the plot function in R (residuals vs fitted, normal Q-Q plot) as well as numerically by the Shapiro-Wilks test, and a post-hoc test was performed using the Tukey Honest Significant Differences (Tukey HSD). All statistical analyses were performed using the software R (R Development Core Team, 2022).
3 Results
Seaweed fly productivity under different feeding substrates
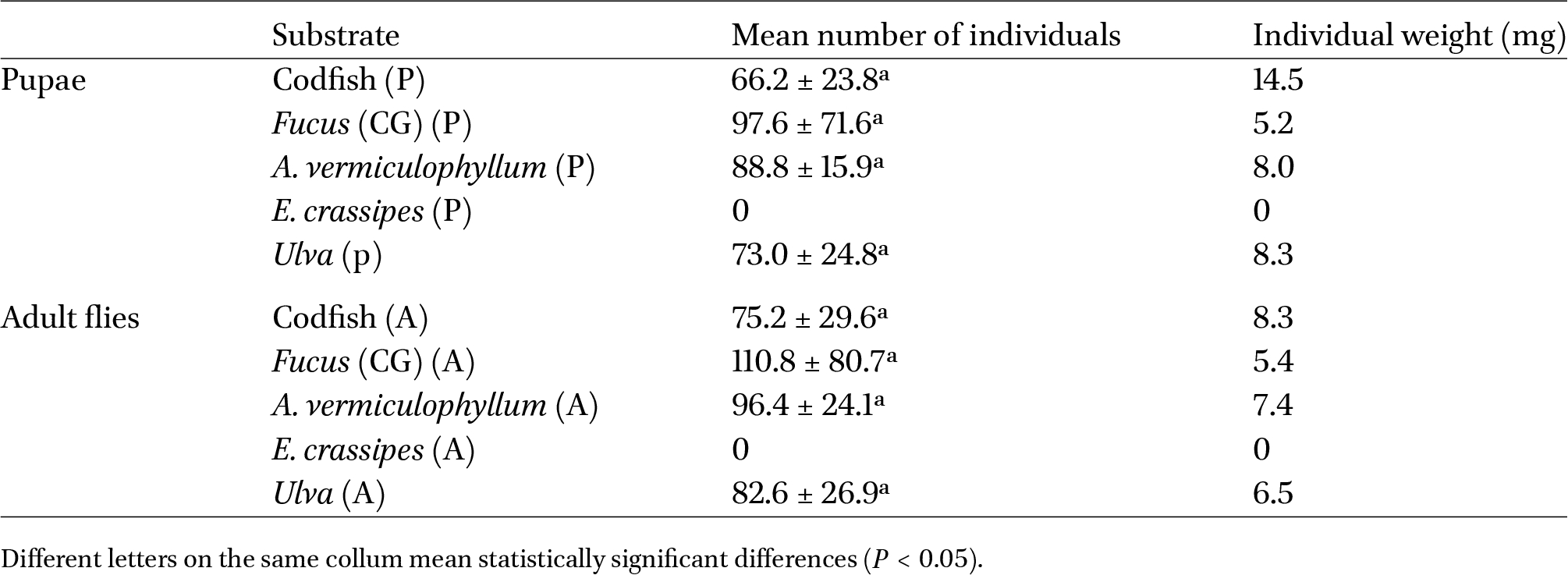
There were no significant differences in the total number of individuals between pupae and adults reared on the different substrates (not accounting for E. crassipes). It is worth mentioning that replicates displayed high variability, namely those supplied with Fucus sp. as feeding substrate, as seen in Table 1. No specimens could complete their life cycle using E. crassipes as a feeding substrate; however, female flies laid eggs on this substrate, and larvae emerged, so no specimens survived beyond the first larval instar. For this reason, this substrate was excluded from further analysis related to the composition of the pupae and adult flies. The substrate on which more larvae moulted into pupae was Fucus sp. (see Figure 2), followed by A. vermiculophyllum and Ulva sp. The feeding substrate on which fewer pupae developed was codfish frames, even though individual mean weight was higher in larvae fed with this substrate (14.5 mg) when compared to Fucus sp. (5.2 mg).
Productivity of pupae (P) and adult (A) flies of Fucellia maritima under different feeding substrates. Mean number of individuals (mean ± standard deviation, n = 5), and individual weight (obtained by dividing the total weight recorded by the total number of individuals, n = 5)
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
Fucellia maritima pupa on on the brown seaweed Fucus sp.
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
Nutritional composition of feeding substrates, pupae and adult flies
The nutritional composition of the feeding substrates employed in the present study is shown in Table 2. Codfish frames presented the highest values of lipid and protein content. The lowest lipid content was recorded in Ulva sp., and the lowest protein content was recorded in Fucus sp., which also presented the highest ash content.
Average composition of feeding substrates (S) tested in the present study, pupae (P), and adult (A) specimens of seaweed fly Fucellia maritima cultured using those feeding substrates, in terms of lipid, protein (substrates (Kp) = 6.25; adults and pupae (Kp) = 4.76), and ash content (dry weight basis, n = 2)
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
The composition of pupae and adult flies reared under the feeding substrates detailed above is also shown in Table 2. Pupae fed with Ulva sp. and A. vermiculophyllum had the highest protein content compared to pupae reared with other feeding substrates. Pupae fed with codfish and Ulva sp. had the highest lipid content and the lowest ash content. Pupae fed with A. vermiculophyllum had the lowest lipid content and the highest ash content. Regarding adult flies, those fed A. vermiculophyllum and Fucus sp. showed the lowest lipid content, while flies fed with codfish showed the lowest protein content.
Amino acids composition of feeding substrates
Pupae and adult flies fed with Ulva sp. showed higher protein content despite this substrateâs lower amount of protein. Therefore, it was important to consider the digestibility of the protein and the bioavailability of the different amino acids. For this reason, the full amino acid profile of the substrates was assessed to confirm the protein quality of the substrate composition used to rear F. maritima. The amino acid composition of the different seaweeds used as feeding substrates (A. vermiculophyllum, Fucus sp., and Ulva sp.), the aquatic plant substrate (E. crassipes), and the marine animal substrate (codfish frames) are all displayed in Table 3. All feeding substrates presented Essential Amino Acids (EAA). Compared to codfish, seaweed substrates had a greater concentration of five of the nine essential amino acids (EAAs): threonine (Thr), valine (Val), isoleucine (Ile), leucine (Leu), phenylalanine (Phe), and tryptophan (Trp). Despite its interesting composition of amino acids, E. crassipes was not used as a substrate by F. maritima.
Amino Acid profile of feeding substrates employed to culture the seaweed fly Fucellia maritima. Data are shown as the mean of two replicates (% of total amino acids)
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
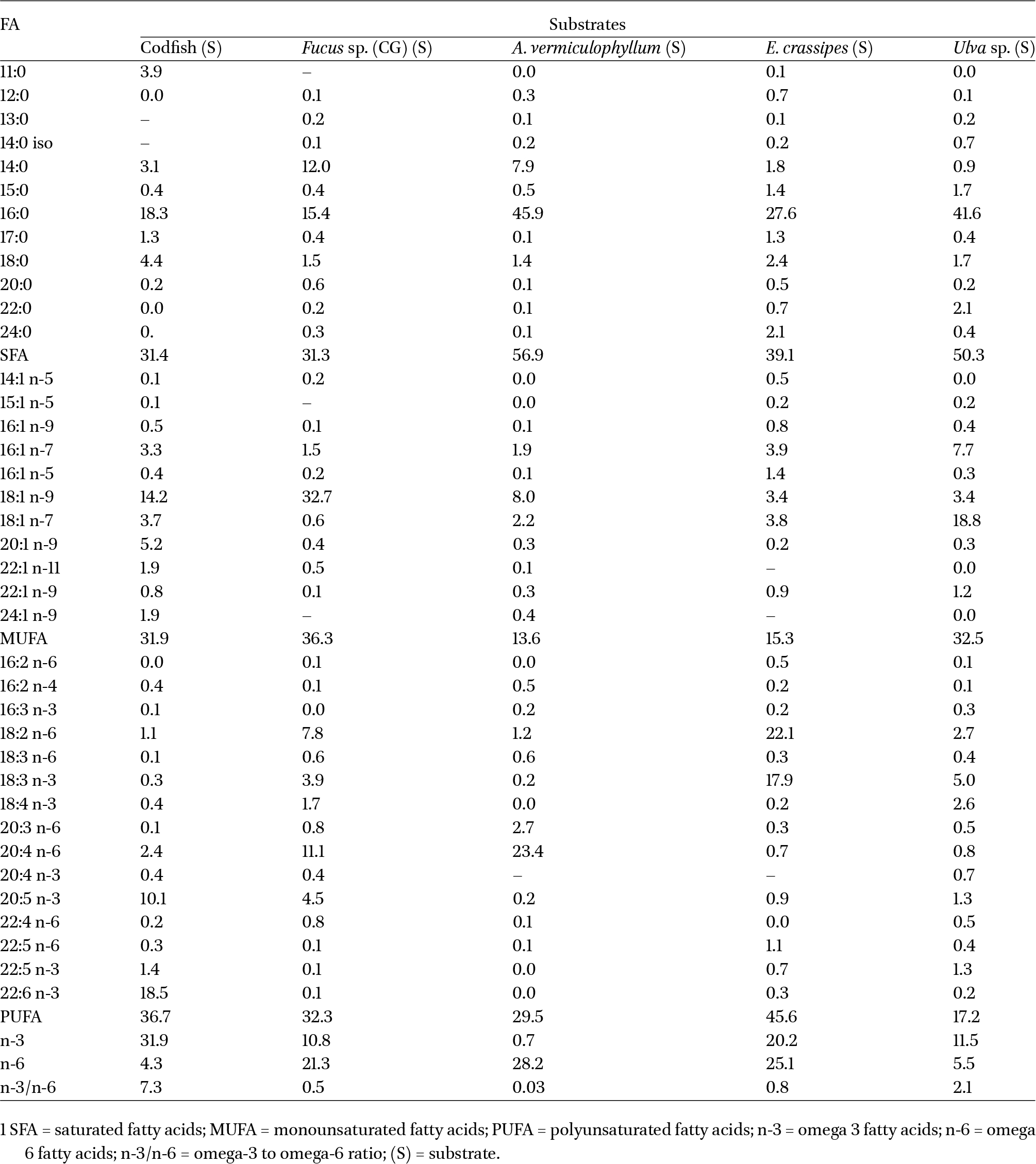
Fatty acid relative abundance (%) of the substrates tested. Data are expressed as mean values of two replicates for Fucus sp. A. vermiculophyllum, E. crassipes and Ulva sp., while four replicates were used for the codfish frames
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
Fatty acids composition of feeding substrates
The composition of fatty acids varied between feeding substrates, as shown in Table 4. The most abundant fatty acids present in the different feeding substrates tested were 22:6 n-3 DHA (18.5%) in codfish frames, 18:1 n-9 oleic acid (OA) (32.7%) in Fucus, and 16:0 palmitic acid (PA) in A. vermiculophyllum, E. crassipes, and Ulva (with 45.9%, 27.6%, and 41.6%, respectively). The brown seaweed Fucus sp. displayed the highest abundance of 20:5 n-3 EPA (4.5%) among the seaweed species tested, and the abundance of 18:3 n-3 alpha-linoleic acid (ALA) (3.9%) on this seaweed is also noteworthy. Codfish frames displayed the highest n-3/n-6 ratio, with E. crassipes featuring high percentages of PUFA due to its content in 18:2 n-6 linoleic acid (LA) and ALA (18:3 n-3); the lowest levels of PUFA abundance were recorded in Ulva sp., with the lowest n-3/n-6 ratio being recorded in A. vermiculophyllum, which also presented the highest levels of saturated fatty acids and the lowest levels of monounsaturated fatty acids (Table 4).
Lipid classes and fatty acids composition of pupae and adult flies
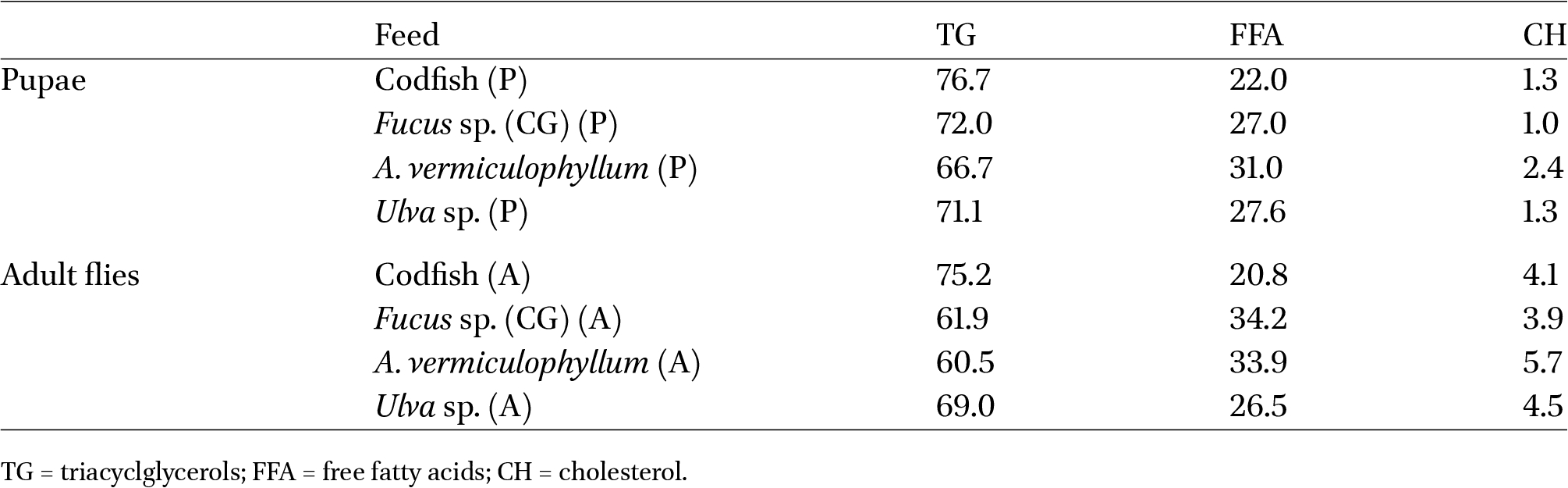
The lipid classes identified in pupae and adult flies were triacycloglycerols (TG), free fatty acids (FFA), and cholesterol (CH) and their abundance is shown in Table 5. The highest value of TG was from pupae fed with codfish; for FFA, the highest value was from pupae fed with A. vermiculophyllum; and for CH, the highest value was from adult flies fed with A. vermiculophyllum.
Lipid classes of Fucellia maritima pupae (P) and adult (A) flies fed on different feeding substrates. Data are shown as mean values (% of total lipid classes, n = 2)
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
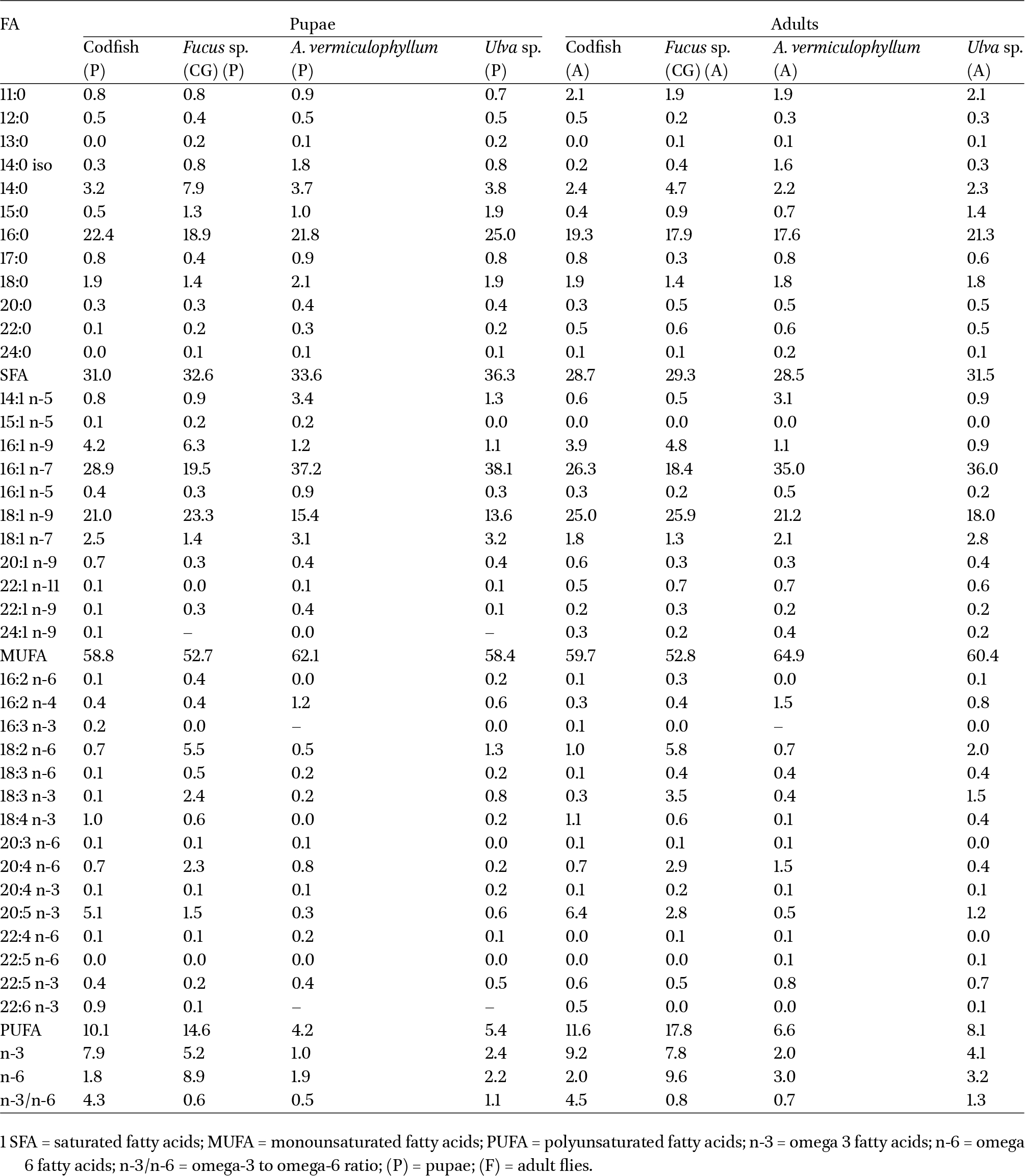
The most abundant fatty acid was OA, both in pupae and adult flies fed with Fucus sp., while for all the other treatments, the most abundant FA was palmitoleic acid (16:1 n-7), as shown in Table 6. Levels of EPA higher than 5% of the total pool of fatty acids were only found on pupae and adult flies fed with codfish frames.
Fatty acid composition (Relative abundance %) of pupae (P) and adult (A) flies of Fucellia maritima fed with different substrates (codfish frames, Fucus sp. A. vermiculophyllum, E. crassipes, Ulva sp.). Data are expressed as mean values of two replicates
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
Noteworthy, the abundance of PUFAs (Table 6) was higher in pupae fed with Fucus sp., followed by codfish frames, and was lower in larvae fed with A. vermiculophyllum. Among these PUFAs, the amount of PUFA n-3 in pupae fed with Fucus sp. was notable. The results were similar for adult flies. The n-3/n-6 ratio was higher in pupae fed with codfish frames, followed by those fed with Ulva sp. Lower values were obtained in individuals fed with A. vermiculophyllum. The same results were observed for adult flies. All samples had a higher percentage of MUFA when compared to PUFA. Still, pupae and adult flies fed with A. vermiculophyllum had the highest percentage of monounsaturated fatty acids.
4 Discussion
The present study investigated the nutritional profile of an understudied European native insect species that was only recently recorded from the coastal area of mainland Portugal (Lourenço et al., 2020). Its nutritional profile and ability to breed in captivity make the seaweed fly F. maritima a suitable candidate as a novel ingredient for marine aquafeed production, affording new potential insect species for this purpose (Lourenço et al., 2022).
Seaweed flies were able to complete their life cycle on most feeding substrates tested in this study. In our facilities they were maintained in captivity for several generations in insect cages (350 mm à 350 mm à 600 mm) under stable climatic conditions (unpubl. data). It is noteworthy to mention that there are no known life tables for this species or mass-rearing protocols described to date.
In this experimental setting, we doubled the populationâs initial size (50 adult flies per replicate) to more than 100 individuals per replicate (sum of pupae and adult flies in Table 1). However, there was a high deviation between the replicates, which can be caused by the unknown ratio of females in the initial population, where a high ratio of females is needed in a mass-rearing context (Fitz-Earle and Barclay, 1989). As mentioned earlier, it is impossible to distinguish sexes in F. maritima without dissecting. Since the experiment ended with the first generation, it was impossible to confirm the exponential population buildup. However, the observed duplication of the number of individuals in the first generation seems to indicate this possibility. To clarify this, more studies on F. maritima population dynamics to test the likelihood of its mass-rearing are necessary.
The invasive plant E. crassipes was not a suitable substrate, and F. maritima larvae could not complete their life cycle using this substrate. It is known that E. crassipes produces secondary metabolites that deter insect feeding, namely quercetin (a flavonoid) and tomatine (a glycoalkaloid), which have documented insecticidal and larvicidal activity against Diptera (Ben Bakrim et al., 2022). Annually, in Ria de Aveiro, these plants are washed from the Vouga River to the beach and comprise about 90% of the beach wrack, overlapping with the naturally most abundant seaweeds, Fucus sp. and Ulva sp. (O. Ameixa, pers. obs.) (Figure 3). This finding reinforces the negative impact that this invasive freshwater plant species can have on the ecosystems it invades and adjacent ones, especially for seaweed flies.
Overall, the type of feeding substrate seems to influence the weight of the pupae of F. maritima, especially when larvae are fed with codfish frames, although this was not confirmed by statistical analysis. As with other insect orders, weight is known to change in Diptera when different feeding substrates are used; examples are known for CHF (Ganda et al., 2022), BSF (Liland et al., 2017), Coelopa frigida, and Coelopa pilipes (Biancarosa et al., 2018b). However, we also observed that the weight of the larvae and adult flies was inversely proportional to the number of individuals, indicating a possible effect of density on the weight of these specimens. Although, as mentioned earlier, there is no statistical test on the weight of individuals, the density was not tested, and the number of individuals presents no significant difference. Nevertheless, the rearing density is known to have a negative effect on the weight of different insect species. For instance, a higher density of BSF can significantly impact the individual larval weight when compared to a treatment with a low density (Yakti et al., 2022). Similar results are noticed in the pupal weight of CHF, with the lower weight seen on the highest density (Kökdener and Kiper, 2021). With YM, there is a similar effect, with the highest larval weight seen in the lower-density treatment (Koutsogeorgiou et al., 2024).
The lipid content recorded for Fucus sp. was above the range of other studies available in the literature (1.4-3.7%) (da Costa et al., 2019; Lorenzo et al., 2017; Neto et al., 2018). Ash content, however, was in the range reported by the same authors (20.7-25.5%), the same being true for protein content (12.9-15.1%). In comparison to other studies, A. vermiculophyllum showed a lower lipid content (0.7-2.0%), while Ulva sp. had a lipid content within the range of other studies (0.9-2.53%) (Afonso et al., 2021; Neto et al., 2018). Both A. vermiculophyllum and Ulva sp. showed lower ash content (19.6-28.9% and 26.4-37.1%, respectively) and protein content inside the range for A. vermiculophyllum and higher for Ulva sp. (14.7-23.6% and 9.3-17.7%) (Afonso et al., 2021; Lopes et al., 2019; Neto et al., 2018). Seasonal changes in protein and lipid values are common in seaweeds, with the highest values being found in winter and the beginning of spring and lower values being found in summer and autumn (Afonso et al., 2021; da Costa et al., 2019). Eichhornia crassipes lipid content (0.9-14.9%) and ash content (12.4-39.8%) were in the range of other studies, but protein content was higher (6.6-15.2%) (Arayana et al., 1984; Hossain et al., 2015; Okoye et al., 2000). The high range found in other studies was probably due to the separation of plant parts by those authors, with roots having the lowest protein and lipid content and leaves having the highest; in our study, the whole plant tissue was ground and homogenised, likely promoting the contrasting results recorded. Codfish frames had a higher lipid (5.2%) and protein (58.9%) content and a lower ash content (30.4%) in comparison to what was previously found in another study that also used codfish frames from the same company but to culture black soldier fly (Rodrigues et al., 2021). The differences in our results may arise from our analyses only including the flesh and skin of codfish frames, while in the study by Rodrigues et al. (2021), the whole ground frames, which contained a considerable mass of bones, were also being used.
Eichhornia crassipes composing most of the biomass of beach wrack (40°38â31.1âN; 8°44â57.7âW).
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001259
All the tested substrates presented EAAs. The vegetable samples had a higher value on five of the nine EAAs when compared to the amino acids present on codfish frames, making them a source of good-quality protein with high digestibility for the seaweed fly F. maritima. The EAA/total AA ratio suggests that almost 40% of the total AA was EAA on all samples; these values are close to the ones reported from other studies addressing Fucus sp. (40.9%) (Lorenzo et al., 2017), and Ulva sp. (40.7%) (Machado et al., 2020), but lower than those reported for A. vermiculophyllum (51%) (Afonso et al., 2021), and E. crassipes (50.3%) (for leaf protein concentrate) (Hontiveros and Serrano, 2015).
The fat content was lower, but the protein content of F. maritima (as a total percentage of dry matter as shown in Table 2) was within the range observed in other terrestrial insect species produced for food and feed, namely BSF and YM (30.7-58.8% and 51.8-59.8%, respectively) (Nogales-MeÌrida et al., 2019). Moreover, F. maritima has a higher protein content than other seaweed flies, like C. frigida and C. pilipes (Biancarosa et al., 2018b). However, the higher protein content observed in adult flies (55.2-56.7%) could be overestimated, as adult insects have a higher content of chitin and non-digestible protein (Jonas-Levi and Martinez, 2017). Considering this, we recommend the study of the total amino acids profile of F. maritima to have a more accurate estimate of its protein content and to select the most adequate conversion factor, as suggested by Janssen et al. (2007).
Considering FAs, F. maritima showed a considerable amount of n-3 PUFAs in their composition, especially in pupae fed with codfish frames, which had a higher n-3/n-6 ratio and a higher level of EPA and DHA than those found in other seaweed flies, C. frigida and C. pilipes (Biancarosa et al., 2018b), but in a lower abundance than what was found in the coastal long-legged fly, Machaerium maritimae (Duarte et al., 2021). Despite these differences, the levels of PUFA were, in general, still higher than those found on terrestrial species, even when these were fed with substrates rich in these biomolecules (Nogales-MeÌrida et al., 2019; Oonincx et al., 2020). This was most evident in pupae whose larvae were fed codfish frames. Moreover, higher inclusions of n-3 PUFA-rich substrates in the diets presented a negative trade-off, such as reduced larval growth in terrestrial species, namely BSF (Ewald et al., 2020; Liland et al., 2017; Rodrigues et al., 2022). In the study done by Liland et al. (2017) it is hypothesised that this trade-off occurs mostly because of the low protein content of the seaweed used and due to its larger particle size when compared to the control diets, suggesting that a further reduction of the particle size of seaweed could improve the growth of the larvae (Liland et al., 2017).
The most abundant fatty acid found on both pupae and adult flies of F. maritima in all feeding treatments (except Fucus sp., for which the most abundant fatty acid was OA) was the monounsaturated palmitoleic acid (16:1 n-7), which, when supplemented in the diet of the Pacific white shrimp Litopenaeus vannamei, can help to protect the growth rate and enzymatic activity of this species against aflatoxins that are often present in aquafeeds (GarcıÌa-PeÌrez et al., 2020).
The saturated PA (the most abundant saturated fatty acid found in animals) and the monounsaturated OA were the other abundant fatty acids recorded. These FAs are important in maintaining the cellular membraneâs physical properties; however, an excess of PA in the diet may increase an inflammatory response in mammalsâ intestinal and hepatic tissues (Ghezzal et al., 2020). These damages to the gut tissue can be mitigated by including EPA in farmed fishâs diets (Shi et al., 2022). However, for zebrafish Danio rerio, a higher inclusion of PA increased their antiviral protection (LibraÌn-PeÌrez et al., 2019). Even though many animal and vegetable fats and oils naturally contain OA, fish raised in aquafeeds that include OA have higher immune responses to bacterial infections, which promotes fish growth and survival rates when infected with Vibrio vulnificus. (Natnan et al., 2022).
Palmitoleic acid and OA are the most abundant fatty acids in other seaweed flies, such as C. frigida and C. pilipes (Biancarosa et al., 2018b). However, for M. maritimae, also a coastal insect, the abundance of PA was much lower when compared to these three seaweed fly species (Duarte et al., 2021). The reason for this might be the difference in their diets, as M. maritimae is an amphibiotic insect, and as with most Dolichopodidae species, both larvae and adult flies may be predaceous, while palmitoleic acid is mostly a product of PA metabolism in the body or can be acquired trophically through the consumption of seaweed (McAlpine et al., 1981; Vannice and Rasmussen, 2014).
Regarding n-3 fatty acids, EPA was much higher in insects fed with codfish frames, suggesting that F. maritima larvae can assimilate the fatty acid content of their feeding substrate, which was much richer on this FA than all others tested in this study. The abundance of EPA in pupae fed with codfish frames was similar to the abundance found in another study using wild-caught F. maritima, with approximately 6% of relative abundance (Lourenço et al., 2024). The abundance of DHA was lower than 1% of the total pool of FA, even in pupae and adult flies whose larvae were fed with codfish frames (this FA was not detected on pupae from specimens fed with A. vermiculophyllum or Ulva sp.). The presence of DHA is reported for the first time in a seaweed fly in the present study, although it has already been found in another marine fly species, namely M. maritimae (Duarte et al., 2021).
Despite the lower abundance of EPA and DHA in seaweed flies than in fish or shrimp, both adult flies and pupae of F. maritima, namely those fed with codfish frames, still present an interesting composition in terms of these essential fatty acids, which can be used as insect meals for the formulation of marine aquafeeds.
5 Conclusions
Due to its short life cycle duration (from 20 to 28 days, egg to adult fly), F. maritima can be a potential novel ingredient for marine aquafeed formulations despite its small size compared with other insect species normally used in insect meals. Preliminary rearing trials by the authors revealed that exponential population growth can be achieved relatively quickly. Since the need for heating room temperature during production is also lower compared to other insect species currently reared, the costs of producing F. maritima as an insect meal may eventually be lower than for other âmainstreamâ insect species. Furthermore, one of the advantages of using native insect species in comparison with exotic ones is that it minimises the potential risks of invasion when commercial-scale farming is put forward in Europe (Lourenço et al., 2022). In addition, one must highlight that F. maritima can also be used to add value to the biomass of the invasive red seaweed A. vermiculophyllum, as it can be used as a feeding substrate for this insect. From this study, one can suggest that to maintain a successful culture of F. maritima, one should most likely use a mixture of Fucus sp. and cod fish frames to achieve higher survival and insect biomass production, which has a more suitable PUFA content for marine aquafeed formulation.
However, the authors know this study lacks strong statistical testing to confirm our findings. Still, overall, the data suggests that the seaweed fly F. maritima has the potential to be used as an aquafeed ingredient, considering their fatty acid profile and high protein content. However, there needs to be more research regarding their capacity to be mass-produced, the stability of their nutritional profile, and developing life tables to guarantee an optimum product for commercialisation, before this species can be considered a true candidate to be used as an aquafeed ingredient. Moreover, like other seaweed flies, F. maritima is dependent on marine organic matter as a substrate, and it is known that seaweeds can accumulate undesirable metals like arsenic, mercury, and cadmium (Biancarosa et al., 2018a). However, Ferrari et al. (2002) have demonstrated that bioaccumulation of mineral and trace elements in reared insects can also be positive, opening new scenarios for feed and food production (Ferrari et al., 2022). Therefore, studies on the bioaccumulation of metals in F. maritima are needed to truly consider this species as a candidate for aquafeed ingredients.
Overall, F. maritima shows the potential to contribute to a higher diversification of insect production, considering that this sector is still in its infancy. It may join the list of potential insect species to be explored as novel ingredients for the formulation of marine aquafeeds.
Corresponding authors; e-mail:Â mjgp@iim.csic.es; olga.ameixa@ua.pt
Acknowledgements
Thanks are due to Bacalhau do Barents, Produtos Alimentares Lda, Gafanha da NazareÌ, Portugal, for the supply of codfish frames.
Conflict of interest
The authors have no conflict of interest to declare.
Funding
Thanks are due to FCT/MCTES for the financial support to CESAM (UIDP/50017/2020+UIDB/50017/2020+LA/P/0094/2020), through national funds and to the project SUShI (CENTRO-01-0145-FEDER-030818) co-funded by Centro 2020 program, Portugal 2020, through the European Regional Development Fund, through FCT/MCTES, and to PORBIOTAÂ â Portuguese E-Infrastructure for Information and Research on Biodiversity (POCI-01-0145-FEDER-022127), financed by FCT through PIDAC national funds and co-funded by the FEDER. Thanks also due to the project IN607B 2023/05 funded by Xunta de Galicia, Galician Agency for Innovation, GAIN. F.S.L. is supported by a PhD grant (PD/BD/150577/2020) funded by FCT/MCTES (https://doi.org/10.54499/PD/BD/150577/2020). O.M.C.C.A. is funded by national funds through the Portuguese Foundation for Science and Technology (FCT) under the project 2022.08112.CEECIND.
References
Afonso, C., Correia, A.P., Freitas, M.V., Baptista, T., Neves, M. and Mouga, T., 2021. Seasonal changes in the nutritional composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the center of Portugal. Foods 10: 1145. https://doi.org/10.3390/foods10051145
Ameixa, O.M.C.C., Duarte, P.M. and Rodrigues, D.P., 2020. Insects, food security, and sustainable aquaculture. In: Filho, L.W., Azul, A.M., Brandli, L., Ãzuyar, P.G. and Wall, T. (eds.) Zero hunger. Springer International Publishing, Cham, Switzerland, pp. 425-435. https://doi.org/10.1007/978-3-319-69626-3_111-1
AOAC, 2003. Official methods of analysis of the Association of Official Analytical Chemists. AOAC International, Washington, DC, USA.
Arayana, G.L., Rao, K.S., Pantulu, A.J. and Thyagarajan, G., 1984. Composition of lipids in roots, stalks, leaves and flowers of Eichhornia crassipes (Mart.) Solms. Aquatic Botany 20: 219-227. https://doi.org/10.1016/0304-3770(84)90088-3
BaÌnki, O., Roskov, Y., Döring, M., Ower, G., HernaÌndez Robles, D.R., Plata Corredor, C.A., Stjernegaard Jeppesen, T., Ãrn, A., Vandepitte, L., Hobern, D., Schalk, P., DeWalt, R.E., Ma, K., Miller, J., Orrell, T., Aalbu, R., Abbott, J., Adlard, R., Aedo, C. and Al, E., 2024. Catalogue of life checklist (version 2024-05-20). Catalogue of Life, Amsterdam, The Netherlands. https://doi.org/https://doi.org/10.48580/dg6lk
Ben Bakrim, W., Ezzariai, A., Karouach, F., Sobeh, M., Kibret, M., Hafidi, M., Kouisni, L. and Yasri, A., 2022. Eichhornia crassipes (Mart.) Solms: a comprehensive review of its chemical composition, traditional use, and value-added products. Frontiers in Pharmacology 13. https://doi.org/10.3389/fphar.2022.842511
Biancarosa, I., Belghit, I., Bruckner, C.G., Liland, N.S., Waagbø, R., Amlund, H., Heesch, S. and Lock, E.J., 2018a. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: benefits of and limitations to their potential use in food and feed. Journal of the Science of Food and Agriculture 98: 2035-2042. https://doi.org/10.1002/jsfa.8798
Biancarosa, I., Liland, N.S., Day, N., Belghit, I., Amlund, H., Lock, E.-J. and Gilburn, A.S., 2018b. The chemical composition of two seaweed flies (Coelopa frigida and Coelopa pilipes) reared in the laboratory. Journal of Insects as Food and Feed 4: 135-142. https://doi.org/10.3920/JIFF2018.0008
Bligh, E.G. and Dyer, W.J., 1959. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37: 911-917. https://doi.org/10.1139/o59-099
Cheng, L., 1976. Marine insects. American Elsevier Publishing Company, New York, NY, USA. Available at: https://escholarship.org/uc/item/1pm1485b
da Costa, E., Domingues, P., Melo, T., Coelho, E., Pereira, R., Calado, R., Abreu, M.H. and Domingues, M.R., 2019. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Marine Drugs 17: 335. https://doi.org/10.3390/md17060335
Duarte, P.M., Maciel, E., Pinho, M., Domingues, M.R., Calado, R., Lillebø, A.I. and Ameixa, O.M.C.C., 2021. Omega-3 on the fly: long-legged fly Machaerium maritimae as a potential source of eicosapentaenoic acid for aquafeeds. Journal of Insects as Food and Feed 7: 1089-1100. https://doi.org/10.3920/JIFF2020.0112
Egglishaw, H., 1960. The life-history of Fucellia maritima (Haliday) (Diptera, Muscidae). Entomologist 93: 225-231.
EU, 2021. Commission Regulation (EU) 2021/1372 of 17 August 2021 amending Annex IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council as regards the prohibition to feed non-ruminant farmed animals, other than fur animals, with protein deri. Official Journal of the European Union 64: 1-21.
EU Regulation 2021/1925, 2021. Commission Regulation (EU) 2021/1925. Official Journal of the European Union 393: 4-8.
European Commission, 2013. Commission Regulation (EU) No 68/2013 of 16 January 2013 on the Catalogue of feed materials. Regulations 29: 1-64.
European Commission (EC), 2017. Council Regulation (EC) 2017/893/ EC of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as regards the provisions on pro. Official Journal of the European Union 138: 1-25.
European Parliament and Council, 2009. Regulation (EC) No 1069/2009. Official Journal of the European Union 300: 1-33.
Ewald, N., Vidakovic, A., Langeland, M., Kiessling, A., Sampels, S. and Lalander, C., 2020. Fatty acid composition of black soldier fly larvae (Hermetia illucens)Â â possibilities and limitations for modification through diet. Waste Mnagement 102: 40-47.
FAO, 2020. The state of world fisheries and aquaculture 2020. FAO, Rome, Italy. https://doi.org/10.4060/ca9229en
FAO, 2022. The state of world fisheries and aquaculture. Towards blue transformation. FAO, Rome, Italy. https://doi.org/https://doi.org/10.4060/cc0461en
FAO, 2024. The state of world fisheries and aquaculture. FAO, Rome, Italy. https://doi.org/10.4060/cd0683en
FAO, IFAD, UNICEF, WFP and WHO, 2022. The state of food security and nutrition in the world 2022. Repurposing food and agricultural policies to make healthy diets more affordable. FAO, Rome, Italy. https://doi.org/10.4060/cc0639en
Ferrari, L., Sele, V., Silva, M., Bonilauri, P., De Filippo, F., Selmin, F., Ãrnsrud, R., Pinotti, L. and Ottoboni, M., 2022. Biofortification of selenium in black soldier fly (Hermetia illucens) prepupae reared on seaweed or selenium enriched substrates. Journal of Insects as Food and Feed 8: 887-899. https://doi.org/10.3920/JIFF2021.0153
Fitz-Earle, M. and Barclay, H.J., 1989. Is there an optimal sex ratio for insect mass rearing? Ecological Modelling 45: 205-220. https://doi.org/10.1016/0304-3800(89)90082-3
Ganda, H., Zannou, E.T., Kenis, M., Abihona, H.A., Houndonougbo, F.M., Chrysostome, C.A.A.M., Chougourou, D.C. and Mensah, G.A., 2022. Effect of four rearing substrates on the yield and the chemical composition of housefly larvae, Musca domestica L. 1758 (Diptera: Muscidae). International Journal of Tropical Insect Science 42: 1331-1339. https://doi.org/10.1007/s42690-021-00651-z
GarcıÌa-PeÌrez, O.D., Tapia-Salazar, M., Nieto-LoÌpez, M.G., Cruz-Valdez, J.C., Maldonado-MunÌiz, M., Guerrero Guerrero, L.M., Cruz-SuaÌrez, L.E. and MarroquıÌn-Cardona, A.G., 2020. Effects of conjugated linoleic acid and curcumin on growth performance and oxidative stress enzymes in juvenile Pacific white shrimp (Litopenaeus vannamei) feed with aflatoxins. Aquaculture Research 51: 1051-1060. https://doi.org/10.1111/are.14451
Ghezzal, S., Postal, B.G., Quevrain, E., Brot, L., Seksik, P., Leturque, A., Thenet, S. and CarrieÌre, V., 2020. Palmitic acid damages gut epithelium integrity and initiates inflammatory cytokine production. Biochimica et Biophysica Acta (BBA)Â â Molecular and Cell Biology of Lipids 1865: 158530. https://doi.org/10.1016/j.bbalip.2019.158530
Glencross, B.D., 2009. Exploring the nutritional demand for essential fatty acids by aquaculture species. Reviews in Aquaculture 1: 71-124. https://doi.org/10.1111/j.1753-5131.2009.01006.x
Hontiveros, G.J.S. and Serrano, A.E., 2015. Nutritional value of water hyacinth (Eichhornia crassipes) leaf protein concentrate for aquafeeds. AACL Bioflux 8: 26-33. Available at: http://www.bioflux.com.ro/home/volume-8-1-2015/
Hossain, E., Sikder, H., Kabir, H. and Sarma, S.M., 2015. Nutritive value of water hyacinth (Eichhornia crassipes). Online Journal of Animal and Feed Research 5: 40-44.
Janssen, R.H., Vincken, J.-P., van den Broek, L.A.M., Fogliano, V. and Lakemond, C.M.M., 2017. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. Journal of Agricultural and Food Chemistry 65: 2275-2278. https://doi.org/10.1021/acs.jafc.7b00471
Jonas-Levi, A. and Martinez, J.-J.I., 2017. The high level of protein content reported in insects for food and feed is overestimated. Journal of Food Composition and Analysis 62: 184-188. https://doi.org/10.1016/j.jfca.2017.06.004
Kökdener, M. and Kiper, F., 2021. Effects of larval population density and food type on the life cycle of Musca domestica (Diptera: Muscidae). Environmental Entomology 50: 324-329. https://doi.org/10.1093/ee/nvaa165
Koutsogeorgiou, E.I., Gourgouta, M., Rumbos, C.I., Grigoriadou, K., Bonos, E., Giannenas, I., Athanassiou, C.G. and Andreadis, S.S., 2024. Density-dependent development of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Journal of Insects as Food and Feed 10: 1387-1395. https://doi.org/10.1163/23524588-00001048
LibraÌn-PeÌrez, M., Pereiro, P., Figueras, A. and Novoa, B., 2019. Antiviral activity of palmitic acid via autophagic flux inhibition in zebrafish (Danio rerio). Fish & Shellfish Immunology 95: 595-605. https://doi.org/10.1016/j.fsi.2019.10.055
Liland, N.S., Biancarosa, I., Araujo, P., Biemans, D., Bruckner, C.G., Waagbø, R., Torstensen, B.E. and Lock, E.-J., 2017. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLOS ONE 12: e0183188. https://doi.org/10.1371/journal.pone.0183188
Lopes, D., Moreira, A.S.P., Rey, F., da Costa, E., Melo, T., Maciel, E., Rego, A., Abreu, M.H., Domingues, P., Calado, R., Lillebø, A.I. and RosaÌrio Domingues, M., 2019. Lipidomic signature of the green macroalgae Ulva rigida farmed in a sustainable integrated multi-trophic aquaculture. Journal of Applied Phycology 31: 1369-1381. https://doi.org/10.1007/s10811-018-1644-6
Lorenzo, J., AgregaÌn, R., Munekata, P., Franco, D., Carballo, J., Şahin, S., Lacomba, R. and Barba, F., 2017. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Marine Drugs 15: 360. https://doi.org/10.3390/md15110360
Lourenço, F., Calado, R., Medina, I. and Ameixa, O.M.C.C., 2022. The potential impacts by the invasion of insects reared to feed livestock and pet animals in Europe and other regions: a critical review. Sustainability 14: 6361. https://doi.org/10.3390/su14106361
Lourenço, F., Calado, R., Pinho, M., Domingues, M.R., Medina, I. and Ameixa, O.M.C.C., 2024. Fatty acid profile and Escherichia coli and Salmonella sp. load of wild-caught seaweed fly Fucellia maritima (Haliday, 1838) (Diptera: Anthomyiidae). Insects 15: 163. https://doi.org/10.3390/insects15030163
Lourenço, F., Prado e Castro, C. and Ameixa, O.M.C.C., 2020. First record of Fucellia maritima (Haliday, 1838) (Diptera, Anthomyiidae) populations in Portugal. Norwegian Journal of Entomology 67: 246-248.
Machado, M., Machado, S., Pimentel, F.B., Freitas, V., Alves, R.C. and Oliveira, M.B.P.P., 2020. Amino acid profile and protein quality assessment of macroalgae produced in an integrated multi-trophic aquaculture system. Foods 9: 1382. https://doi.org/10.3390/foods9101382
McAlpine, D.K., Peterson, B., Shewwell, G., Teskey, H., Vockerhoth, J. and Wood, D., 1981. Manual of Nearctic Diptera 1. Research Branch. Agriculture Canada, Ottawa, Canada. Available at: https://publications.gc.ca/collections/collection_2016/aac-aafc/A54-3-27-eng.pdf
Natnan, M.E., Low, C.F., Chong, C.M., Daud, N.I.N.A.A., Om, A.D. and Baharum, S.N., 2022. Comparison of different dietary fatty acids supplement on the immune response of hybrid grouper (Epinephelus fuscoguttatus à Epinephelus lanceolatus) challenged with Vibrio vulnificus. Biology 11: 1288. https://doi.org/10.3390/biology11091288
Neto, R., Marçal, C., QueiroÌs, A., Abreu, H., Silva, A. and Cardoso, S., 2018. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as functional ingredients. International Journal of Molecular Sciences 19: 2987. https://doi.org/10.3390/ijms19102987
Nogales-MeÌrida, S., Gobbi, P., JoÌzefiak, D., Mazurkiewicz, J., Dudek, K., Rawski, M., KieronÌczyk, B. and JoÌzefiak, A., 2019. Insect meals in fish nutrition. Reviews in Aquaculture 11: 1080-1103. https://doi.org/10.1111/raq.12281
Okoye, F.C., Ilesanmi, B.D. and Daddy, F., 2000. The nutritive value of water hyacinth (Eichhornia crassipes) and its utilisation in fish feed. In: Proceedings of the International Conference on Water Hyacinth, New Bussa, Nigeria, pp. 65-70.
Oonincx, D.G.A.B., Laurent, S., Veenenbos, M.E. and van Loon, J.J.A., 2020. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Science 27: 500-509. https://doi.org/10.1111/1744-7917.12669
Rodrigues, D.P., Calado, R., Ameixa, O.M.C.C., Valcarcel, J. and VaÌzquez, J.A., 2021. Valorisation of Atlantic codfish (Gadus morhua) frames from the cure-salting industry as fish protein hydrolysates with in vitro bioactive properties. LWT 149: 111840. https://doi.org/10.1016/j.lwt.2021.111840
Rodrigues, D.P., Calado, R., Pinho, M., RosaÌrio Domingues, M., Antonio VaÌzquez, J. and Ameixa, O.M.C.C., 2022. Bioconversion and performance of black soldier fly (Hermetia illucens) in the recovery of nutrients from expired fish feeds. Waste Management 141: 183-193. https://doi.org/10.1016/j.wasman.2022.01.035
Shi, P., Liao, K., Xu, J., Wang, Y., Xu, S. and Yan, X., 2022. Eicosapentaenoic acid mitigates palmitic acid-induced heat shock response, inflammation and repair processes in fish intestine. Fish & Shellfish Immunology 124: 362-371. https://doi.org/10.1016/j.fsi.2022.04.011
Team, Rs., 2022. RStudio: integrated development environment for R. PBC, Boston, MA, USA. Available at: http://www.rstudio.com/
United Nations Department of Economic and Social Affairs, 2016. World population prospects. The 2015 revision â volume II: demographic profiles. https://doi.org/10.18356/cd7acf62-en
van Dijk, M., Morley, T., Rau, M.L. and Saghai, Y., 2021. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010-2050. Nature Food 2: 494-501. https://doi.org/10.1038/s43016-021-00322-9
Vannice, G. and Rasmussen, H., 2014. Position of the Academy of Nutrition and Dietetics: dietary fatty acids for healthy adults. Journal of the Academy of Nutrition and Dietetics 114: 136-153. https://doi.org/10.1016/j.jand.2013.11.001
Yakti, W., Schulz, S., Marten, V., Mewis, I., Padmanabha, M., Hempel, A.J., Kobelski, A., Streif, S. and Ulrichs, C., 2022. The effect of rearing scale and density on the growth and nutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae. Sustainability 14: 1772. https://doi.org/10.3390/su14031772






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}