Abstract
The impact of blanching and storage temperature (−20 °C and −80 °C) on the formation of volatile organic compounds and changes in quality characteristics of lesser mealworm (Alphitobius diaperinus) larvae was examined over seven weeks. Acid and peroxide values, pH, and colour of lesser mealworm larvae were significantly affected by the studied variables. The changes in these characteristics as well as in the volatile profile indicated occurring lipid oxidation and lipid hydrolysis processes during the storage. The intensity of these reactions was mostly influenced by blanching treatment and storage time, and a lesser extent by storage temperature. It was proposed that changes in composition and intensity of volatiles can be used for rapid quality assessment of lesser mealworm larvae during frozen storage. Thus, the selection of optimum storage conditions for lesser mealworm larvae is important for the prevention of undesirable chemical reactions and preservation of the quality of insect biomass.
1 Introduction
Interest in edible insects as a sustainable food alternative has emerged in recent years. One of the reasons is that insect production has a significantly lower environmental footprint with less land and water use and lower greenhouse gases emission in comparison to traditional livestock (van Huis and Oonincx, 2017). From a nutritional point of view, insects are a good source of protein, essential amino acids and unsaturated fatty acids. Additionally, insects contain considerable amounts of vitamins such as vitamin B12 and minerals such as iron (Rumpold and Schlüter, 2013).
Production of insects for human consumption is linked to their processing (i.e. blanching, grinding) and storage. It was shown that the storage behaviour of whole unprocessed insects by freezing is affected by the presence of antifreeze proteins, which can inhibit ice crystals growth and thus maintain water unfrozen (Graham et al., 1997). Unfrozen water could favour biochemical reactions such as lipid oxidation and lipid hydrolysis resulting in undesirable colour and flavour formation (Leygonie et al., 2012). In particular, lipid oxidation products can react with amino acids producing Strecker aldehydes and ketoacids (Hidalgo and Zamora, 2016), which are important for flavour of foods. Dynamic of lipid oxidation can be assessed by analysis of volatile compounds and detection markers of lipid oxidation such as hexanal and propanal (Grebenteuch et al., 2021). Moreover, unfrozen water in insects at −20 °C favours protein hydrolysis resulting in changes in chemical properties and protein functionalities already during the first week of storage (Wessels et al., 2020). It indicates that storage temperature and time can be important factors for the preservation of insect quality during storage.
The microbial activity of edible insects, largely due to insect gut microbiota, should be also taken into consideration to ensure the safety of insects for human consumption. Moreover, microbial activity could have an adverse effect on the quality profile of stored insects (Klunder et al., 2012; Wynants et al., 2018). In this regard, blanching is widely used as a microbial decontamination technique to reduce microbial loads and prevent browning of edible insects (Mancini et al., 2019; Wynants et al., 2018). It was shown that blanching of yellow mealworm larvae in boiling water for 10-40 s leads to considerable log reductions of total viable count, Enterobacteriaceae, lactic acid bacteria, yeasts and molds, and can be used as a pretreatment for refrigerated storage (Vandeweyer et al., 2017). Additionally, blanching before storage can inactivate the enzymes that cause lipid oxidation and browning (Cacchiarelli et al., 2022; Larouche et al., 2019; Reyes De Corcuera et al., 2004). Another study demonstrated that beside assuring microbial safety, blanching of yellow mealworm larvae can also improve colour characteristics and the composition of macro-nutrients (Ribeiro et al., 2024). While heat treatment is an important step from a safety point of view, it leads to the denaturation of insect proteins resulting in the alteration of techno-functional properties of proteins and their potential application as food ingredients (Mishyna, Keppler and Chen, 2021).
Previous studies demonstrated that both processing and storage conditions affect quality of insect oils on an example of Rhynchophorus phoenicis. In particular, prolonged freezing and electrical drying at 50 °C affects oil stability, while refrigeration for less than 3 days or freezing for less than a month could preserve lipid quality (Tiencheu et al., 2013). Therefore, the selection of optimum storage conditions is important for preservation of nutritional, functional, and sensory characteristics of edible insects which can be affected by numerous undesirable reactions during storage. The quality of insect biomass also affects the quality of further produced insect-derived food ingredients and foods with edible insects. So far, to the best of our knowledge, the influence of low-temperature storage on volatile profiles and quality-associated indicators of edible insects was not previously studied. Therefore, this study aimed to estimate the effect of blanching, storage temperature (−20 °C and −80 °C), and storage time (7 weeks) on the volatile profiles and quality indicators of lesser mealworm larvae.
2 Materials and methods
Materials
Larvae of Alphitobius diaperinus (lesser mealworm) at 28 days old were provided by Protifarm Holding NV (Ermelo, the Netherlands). Not blanched larvae were prepared by snap freezing of alive larvae with liquid nitrogen. The blanching step was performed at 85 °C for 1 min, and insects were provided by Protifarm Holding NV. Afterwards, not blanched and blanched larvae were stored in sealed plastic buckets at −20 °C and −80 °C (EvoSafe™-series, Snijders Labs, Tilburg, the Netherlands) for 7 weeks.
pH and colour analysis
Larvae slurries were prepared by blending frozen lesser mealworm larvae with demineralised water in 1:2 (w/w) larvae/water ratio for 1 min to obtain a homogenous mixture. Then, larvae slurries were used for pH and colour measurements. The values of pH were measured using a pH meter. Colour evaluation was performed using a colour meter (Elscolab BV, Ede, the Netherlands) to monitor the browning intensity over time. Colour was analysed for L, a, and b parameters, and the browning index was calculated based on these parameters according to Pathare et al. (2013).
Lipid analyses
Lipid extraction
The lipid extraction method was based on Bligh and Dyer method (Bligh and Dyer, 1959). The volumes of chloroform (RS, CARLO ERBA Reagents GmbH, Emmendingen, Germany), methanol (HPLC gradient, Actu-All Chemicals BV, Oss, the Netherlands) and water were kept in proportions of 2:2:1.8 (v/v/v) by taking into account the water content of the not blanched (48%) and blanched (42%) larvae samples. After mixing the larvae samples with the abovementioned solvents, the mixture was vortexed for 1 min and centrifuged for 10 min at 2,000 rpm at 20 °C (Heraeus Multifuge X3R, Thermo Scientific, Waltham, MA, USA). After centrifugation, the upper layer was taken. Then 1:1 (v/v) chloroform:methanol mixture was added to this upper layer and after centrifugation (10 min, 2,000 rpm, 20 °C) the lower layer was collected. The solvent from this layer was evaporated at 40 °C using a vacuum rotary evaporator (R-215, Buchi, Flawil, Switzerland) to obtain the lipid fraction.
Peroxide value
Peroxide value was determined by using a modified method according to Jeon et al. (2016). One gram of lipid was dissolved in a 25 mL solution of glacial acetic acid (ACS, ISO, Reag. Ph. Eur., VWR Chemicals)-isooctane (ACS, Reag. Ph. Eur., VWR Chemicals, Amsterdam, the Netherlands) in 3:2 (v/v) ratio, then 1 mL of saturated potassium iodide (ISO, Reag. Ph Eur, Merck KGaA, Darmstadt, Germany) solution was added to the mixture and mixed in the dark for 1 min. Further, 30 mL of distilled water (Veolia Water Technologies Netherlands B.V., Ede, the Netherlands) and 1 mL of 1% starch (ISO, Merck KGaA, Darmstadt, Germany) solution (w/v) were added, and the mixture was titrated with 0.002 N sodium thiosulfate (Merck KGaA, Darmstadt, Germany) until the blue colour disappeared. Peroxide value was expressed as meq O2/kg insect oil.
Acid value
The acid value was determined by using the modified method based on Jeon et al. (2016). The neutralised solvent was 96% ethanol (GPR Rectapur, VWR Chemicals, Amsterdam, the Netherlands) and phenolphthalein-methanol solution (ACS, Reag. Ph. Eur, Merck KGaA, Darmstadt, Germany) was used as a colour indicator. Firstly, the colour indicator solution was added to the required amount of solvent in a ratio of 2:125 (v/v) and was neutralised with alkali to a faint, but permanent pink colour. Subsequently, the insect oil sample (0.5 g) was added to the neutralised solution and the mixture was shaken until the sample was completely dissolved in the solution. Then the mixture was titrated with 0.1 N standard potassium hydroxide (Merck KGaA, Darmstadt, Germany) to the first permanent pink colour (30 s) of the same intensity as that of the neutralised solvent. The acid value was expressed as mg KOH/g insect oil.
Volatile compounds analysis with GC-MS
Volatile compounds of lesser mealworm larvae were analyzed with headspace (HS) solid-phase microextraction (SPME) and GC-MS. Extraction of volatile compounds was done by using a 50/30 μm divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS) (2 cm) fiber (Sigma-Aldrich, St. Louis, MO, USA). First, 2 g of frozen larvae were weighed into 10 mL headspace vials sealed with 20 mm PTFE/silicone magnetic caps (10 ml × 23 × 46 mm; Phenomenex, Torrance, CA, USA) and incubated at 40 °C for 10 minutes. Subsequently, the extraction of volatile compounds to the SPME fiber was done for 10 minutes at 40 °C followed by 10 minutes of desorption at the same temperature. The volatile compounds were injected into a GC-MS (Thermo TraceGc, Breda, the Netherlands) system with a Stabilwax-DA column (30 m × 0.25 mm, 1 μm) (Restek, Bellefonte, PA, USA) via an autosampler (TriPlus, Thermo Scientific, TX, USA). The following temperature program was used: initial temperature 40 °C for 3 minutes, then the temperature was raised to 220 °C with a rate of 10 °C/min. The final temperature of 220 °C was held for 5 minutes. Carrier gas helium was fed with a constant flow rate at 1 mL/min and the MS ion source was maintained at 225 °C in full scan. Electron impact mode (EI/MS) was set at 70 eV with the mass range of 33-250 m/z. All samples were measured in triplicate. Identification of the volatile compounds was done by comparing fragmentation mass spectra of the samples to data available from the NIST (National Institute of Standards and Technology, Gaithersburg, MD, USA) library.
Statistical analysis
All experiments were performed in triplicate. The results were reported as mean values ± standard deviation. IBM SPSS Statistics 23 was used to perform one-way ANOVA on all data. A post-hoc Tukey HSD was used to identify the significant differences. Differences were considered significant at

Changes in pH values of not blanched and blanched mealworm larvae during 7 weeks of storage at −20 °C and −80 °C. Different letters indicate difference (
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001051
Browning index of not blanched and blanched lesser mealworm during 7 weeks of storage at −20 °C and −80 °C. Different letters indicate difference (
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001051
3 Results and discussion
Changes in pH, colour, peroxide and acid values
pH
Blanching treatment led to an increase in pH of larvae from 7.1 to 7.3 (Figure 1), that is similar to the previously reported pH increase of lesser mealworms after blanching (Wynants et al., 2018). A subsequent gradual decrease in pH values of both blanched and not blanched larvae was observed at both temperatures during 7 weeks of storage. In particular, a significant decrease from the initial pH of 7.1 to 6.4 (−20 °C) and 6.6 (−80 °C) of not blanched larvae can be caused by the release of hydrogen ions as a result of protein hydrolysis during storage. It can be due to the presence of unfrozen water as it has been previously demonstrated for lesser mealworms stored at −20 °C (Wessels et al., 2020). Moreover, native lipases of insects can cause lipid hydrolysis leading to the release of free fatty acids as has been shown for larvae of flesh fly stored at −20 °C (Cabot and Lumb, 1981). Free fatty acids in turn cause a decrease in pH values. In fact, the pH decrease was less pronounced in blanched larvae with pH changes from around 7.3 to 7.0 (−20 °C) and 7.1 (−80 °C) that could be related to the inactivation of native enzymes during the blanching process and thus preventing protein and lipid hydrolysis and change in pH (Wessels et al., 2020).
Colour
At day 0, browning index of blanched larvae (16.9-19.0) was significantly lower than that of not blanched larvae (42.8-43.9) that can be attributed to the inactivation of enzymes causing browning. This trend continued during 7 weeks of storage at both temperatures, i.e. browning index of blanched larvae (15.9-23.4) was significantly lower (
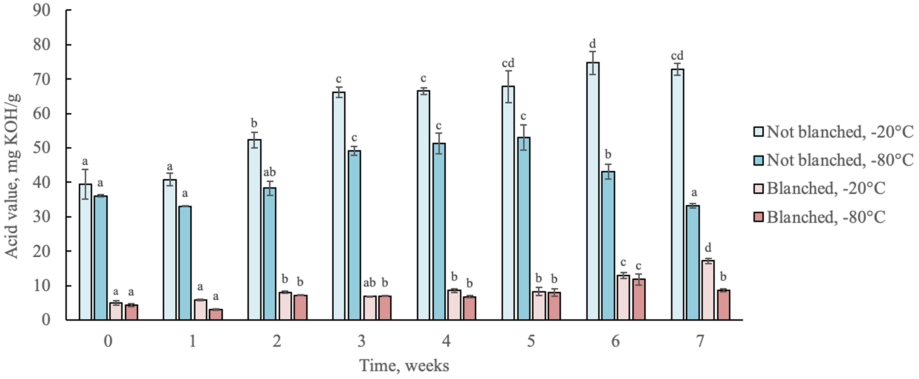
Changes in acid values of lipids from not blanched and blanched lesser mealworm during 7 weeks of storage at −20 °C and −80 °C. Different letters indicate difference (
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001051
Peroxide value
Peroxide value indicates primary lipid oxidation processes and is one of the oil quality indicators. Peroxide values of blanched and not blanched larvae stored for 7 weeks at −20 °C and −80 °C are shown in Supplementary Table S1. Peroxide values varied between 0.41 and 1.34 meq O2/kg insect oil except for one value of 3.46 meq O2/kg insect oil (blanched, −20 °C, 1 week of storage). These values are below the maximum recommended limit of peroxide value of ≤5 meq O2/kg fat recommended for yellow mealworm oil (Turck et al., 2021) indicating a good quality of larvae oil in terms of lipid oxidation. However, peroxide values of all samples fluctuated without a clear trend during storage. In this case, an increase in peroxide values may indicate accumulation of primary oxidation products (Jeon et al., 2016), while a decrease in these values can be referred to hydroperoxides decomposition to various oxidation products (Guo et al., 2019). It should be taken into account that the decomposition of peroxides to secondary products may lead to an underestimation of the degree of oxidation when measuring peroxide concentration (Ross and Smith, 2006). In terms of volatiles, it has been previously found that secondary degradation products from lipid oxidation appear after time lag to peroxide formation (Grebenteuch et al., 2021).
Acid value
As a measure of the amount of free fatty acids present, the acid values of lipids from blanched and not blanched larvae stored at −20 °C and −80 °C for 7 weeks are shown in Figure 3. In general, acid values of blanched larvae were significantly lower (2.96-17.15) than those of not blanched (33.02-74.73) indicating prevention of any reactions that might cause acid production by heat treatment. Although both types of larvae showed a trend for the increase of acid value during the storage, the increase was more pronounced for not blanched larvae. In particular, acid values of not blanched larvae stored at −20 °C increased during the storage from 39.37 to 72.77 mg KOH/g, indicating a faster hydrolysis rate than blanched larvae and being in line with the trend of a decrease in pH value. Correspondingly, a less noticeable increase in acid values was observed for not blanched larvae stored at −80 °C with an increase from 36.07 mg KOH/g to 53.03 mg KOH/g in 5 weeks and then decrease to the initial level (33.13 mg KOH/g) by week 7. Lower acid values of the samples that were stored at −80 °C can indicate faster enzymatic reactions at higher storage temperatures, being in this study at −20 °C. Moreover, the acidity of blanched larvae was a few times lower than those of not blanched larvae that can be related to enzyme inactivation caused by heat treatment.
Changes in volatile profile
Study of volatile compounds is important for the assessment of the quality of foods (Costello et al., 2023). In this study, a total of 39 volatile organic compounds (VOCs) were identified in the headspace of blanched and not blanched lesser mealworm larvae at day 0 (Table 1). Overall changes in the volatile profile of the larvae stored at −20 °C and −80 °C for 7 weeks are shown in Supplementary Figure S1. The identified VOCs were from different compound classes namely aldehydes, acids, ketones, alcohols, alkanes, aromatic compounds, esters, nitriles, pyrroles and sulphur compounds. Similarly, a broad spectrum of volatiles was previously identified in other edible insect species (Kröncke et al., 2019; Mishyna et al., 2020; Tzompa-Sosa et al., 2019; Yeo et al., 2013).
For an overall evaluation of VOCs emission during storage of lesser mealworm larvae, a PCA was performed and the resulting PCA biplot with the first two principal components (PCs) is shown in Figure 4. The first two PC accounted for 53.8% of the total variance. PC2 discriminated the samples based on the blanching process with high negative loadings of volatile acids (e.g. 2-methyl propanoic acid, octanoic acid, 3-methyl butanoic acid) and (Z)-2-penten-ol in not blanched larvae whereas high positive loadings for 2-butanone, 3-methyl-butanal, dimethyl disulfide and nonanal in blanched larvae. High negative loadings of volatile acids in blanched larvae indicates that these volatile compounds were more dominant in not blanched larvae. This observation in volatile acids also correlates well with the higher pH decrease and acid values observed in not blanched larvae stored for 7 weeks at −20 °C. PC1, on the other hand, discriminated the samples based on storage time. There was no clear separation based on storage temperature neither on PC1, nor PC 2. Thus, blanching and storage time were more influential on VOC profiles compared to the storage temperature (−20 °C and −80 °C).
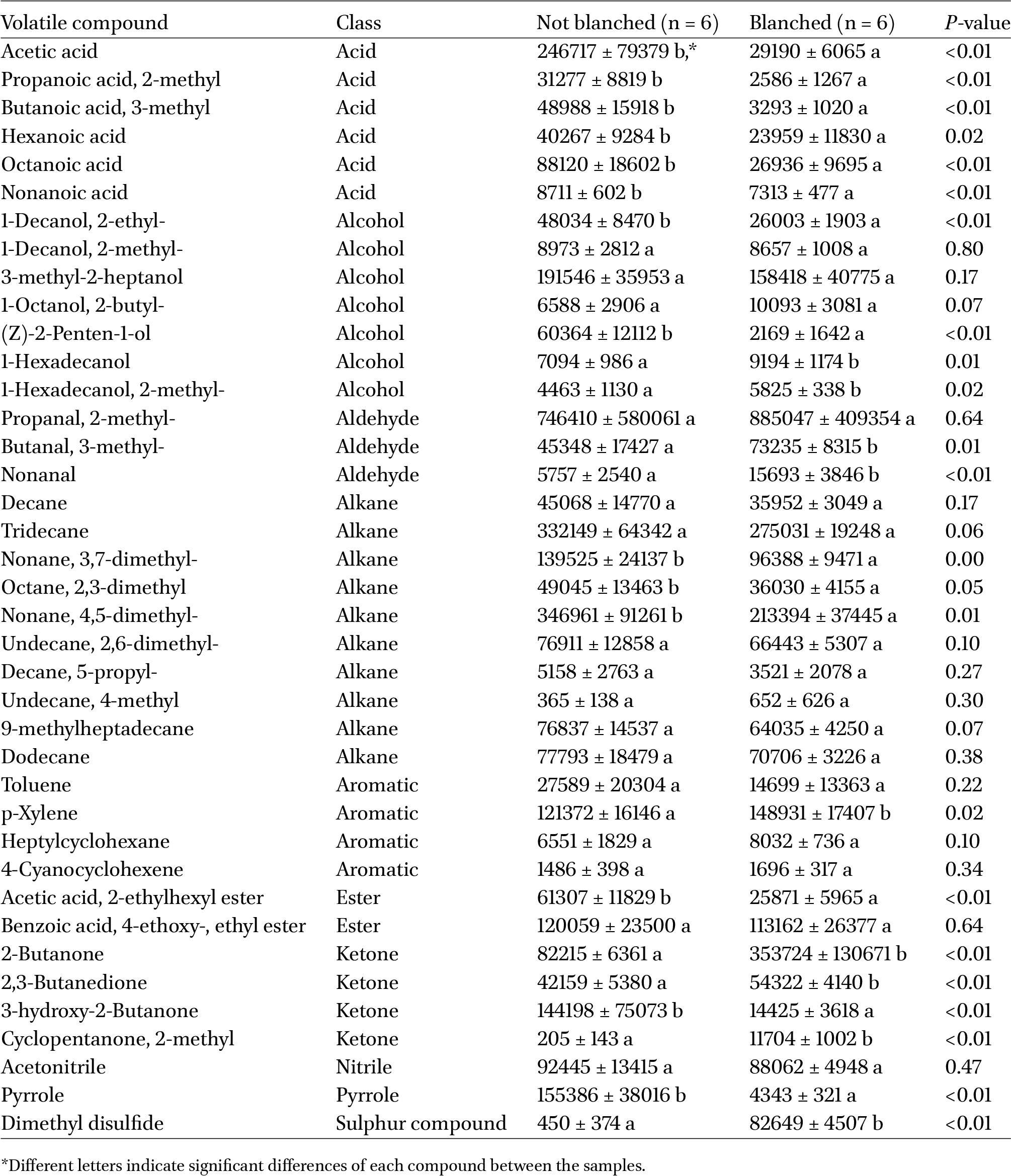
Headspace volatile compounds (peak area ± standard deviation) of not blanched and blanched lesser mealworm larvae at day 0
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001051
Principal Component Analysis (PCA) biplot of volatile profile of not blanched (NB, ▲) and blanched (B, ”) lesser mealworm during 7 weeks of storage at −20 °C (grey colour) and −80 °C (black colour). The biggest triangle (▲) and dot (”) indicate the centre of the not blanched and blanched larvae clusters, respectively. Numbers on the dots and triangles indicate storage weeks.
Citation: Journal of Insects as Food and Feed 11, 3 (2025) ; 10.1163/23524588-00001051
The variations with storage time observed in VOC profile of not blanched larvae were much higher than in blanched larvae (Table 1, Supplementary Figure S1), as it can also be seen on the PCA biplot. This can be explained by the heat treatment applied during blanching process that leads to the denaturation of the native proteases and lipases of lesser mealworm larvae and lowering of microbial decontamination in the lesser mealworm larvae (Vandeweyer et al., 2017). As a result, due to the lower enzymatic activity, the concentration of VOCs emitted from blanched and not blanched larvae differed already on day 0 (Table 1). Among these compounds, blanched and not blanched larvae differed in 24 volatile compounds (
Acids
The content of volatile acids in all samples differed and fluctuated depending on storage temperature and storage time (Supplementary Figure S1). Overall, the content of major volatile acids such as acetic acid and 2-methyl propanoic acid remained significantly higher in not blanched larvae for most time points at both temperatures compared to blanched larvae. For acetic acid, it can be related to the reduction of total aerobic count and lactic acid bacteria by blanching treatment (Vandeweyer et al., 2017). Insect gut microbiota in its turn can be involved in fermentation processes with the emission of acetic acid (Pinu and Villas-Boas, 2017). Similar to our study, acetic acid was previously detected in oil extracted from lesser mealworms, yellow mealworms and crickets (Tzompa-Sosa et al., 2019) and as a volatile from unprocessed locusts and silkworms (Mishyna et al., 2020). It has been also suggested that 2-methyl propanoic acid from Dubia cockroach oil is produced by gut microflora as well (Tzompa-Sosa et al., 2019). Interestingly, a steady decline of 2-methyl propanoic acid was observed for not blanched larvae stored at both temperatures with the minimum values at weeks 6 and 7. It can be assumed that the decomposition of 2-methyl propanoic acid during the storage and the absence of processes that could lead to a detectible increase in the emission of this compound.
Hexanoic acid was previously reported in vacuum-dried yellow mealworm (Tenebrio molitor) (Kröncke et al., 2019), freeze-dried house crickets (Acheta domesticus), Jamaican field crickets (Gryllus assimilis) (Khatun et al., 2021), and frozen fresh and salted boiled male giant water bugs (Kiatbenjakul et al., 2015). In this study, hexanoic acid content fluctuated without a clear trend as a function of blanching and storage conditions. Its presence can be related to numerous occurring reactions, but the exact mechanism still should be studied. For instance, hexanoic acid is known as a product of oleic acid oxidation. Oleic acid is prone to oxidation due to its unsaturation and is one of the dominant fatty acids in lesser mealworm, followed by palmitic acid and linoleic acid (Adámková et al., 2016). Oxidation of oleic acid might also be a cause for an increase in the emission of octanoic acid from blanched larvae after 5 weeks of storage at both temperatures in comparison to day 0. However, the octanoic acid content of not blanched larvae lowered after 2-3 and 6-7 weeks of storage at both temperatures.
Similar to our study, nonanoic acid was previously reported in lesser mealworm oil (Tzompa-Sosa et al., 2019). The amount of produced nonanoic acid did not change significantly in all samples during the first week of storage, but a significant increase was observed between second and fifth weeks. Further, between weeks 4 and 6, not blanched larvae emitted more nonanoic acid at both temperatures than blanched larvae, which might indicate more intensive linolenic acid oxidation.
Aldehydes
Aldehydes are important breakdown products of secondary lipid oxidation that contribute to rancid off-flavours and possess low threshold values (Ross and Smith, 2006). Volatiles 2-methyl-propanal, nonanal, and 3-methyl-butanal were identified in the headspace of blanched and not blanched lesser mealworms. In particular, nonanal represents a special interest to assess the quality of lesser mealworm during the storage as it was proposed as a marker for lipid oxidation, for instance, for silver carp (Hypophthalmichthys molitrix) (Kunyaboon et al., 2021). In this study, nonanal was determined in blanched larvae on day 0 with a three times higher amount for not blanched larvae. This can be due to oleic acid oxidation (Khatun et al., 2021) caused by the thermal degradation of oleic acid during blanching of mealworms as has been shown for olive oil (Nunes et al., 2013). Storage did not have a strong effect on the formation of nonanal; and nonanal content remained higher for blanched larvae over 7 weeks in comparison to not blanched larvae (Supplementary Figure S1). As a naturally occurring volatile, nonanal was previously identified as volatile secretions from the European stink bug Graphosoma lineatum (Šanda et al., 2012), volatile defensive secretions of three species of Pyrrhocoridae (Krajicek et al., 2016).
Other aldehydes such as 2-methyl-propanal and 3-methyl-butanal were identified in lesser mealworms on day 0. While 2-methyl-propanal content did not differ between blanched and not blanched larvae, 3-methyl-butanal was emitted at a significantly higher amount from blanched mealworms than from not blanched. It can be caused by the blanching-induced formation of the Strecker aldehyde such as 3-methyl-butanal (Rainer Cremer and Eichner, 2000).
Alcohols
Volatile alcohols from lesser mealworm larvae are represented by 2-ethyl-1-decanol, 2-methyl-1-decanol, 3-methyl-2-heptanol, 2-butyl-1-octanol, 2-ethyl-1-decanol, (Z)-2-penten-1-ol, 1-hexadecanol, and 2-methyl-1-hexadecanol. On day 0, blanched larvae emitted a significantly lower amount of 2-penten-1-ol and 2-ethyl-1-decanol than not blanched larvae that is in accordance with the study that blanching with water resulted in a decrease of volatile alcohols and ketones of Cordyceps militaris (Wu et al., 2019). No significant effect of storage was observed on 2-penten-1-ol of blanched larvae at both temperatures for 7 weeks. In not blanched larvae, 2-penten-1-ol content fluctuated during the storage with a gradual decrease to the lowest values after 6 weeks of storage. However, these values were not higher than on day 0 that might indicate the decomposition of 2-penten-1-ol during the storage.
Ketones
Ketones 2-butanone, 2,3-butanedione, 3-hydroxy-2-butanone, and 2-methyl cyclopentanone were identified as volatiles from blanched and not blanched lesser mealworm larvae. 2-butanone is known as a pheromone of Strategus aloeus (Coleoptera) along with 3-pentanone and sec.-butyl acetate (Rochat et al., 2000), and a study on semiochemicals of confused flour beetle Tribolium confusum identified 2-butanone in adults and larvae (Abuelnnor et al., 2010). Volatile 2-butanone was determined in the headspace from rack oven-dried and vacuum-dried Tenebrio molitor larvae (Kröncke et al., 2019) and in freeze-dried locusts and microwave-dried silkworms (Mishyna et al., 2020). On day 0, blanched larvae emitted a significantly higher amount of 2-butanone than not blanched that can be related to the exposure to heat, and this trend was observed during the storage in weeks 3-5 and 7 (Supplementary Figure S1). The lowest emission was observed for not blanched larvae stored at −80 °C.
In this study, blanched larvae had higher 2,3-butanedione content than not blanched larvae on day 0. It was previously reported, that 2,3-butanedione can be generated as a result of Maillard reaction via a single pathway involving only glucose carbon atoms as has been demonstrated in D-glucose/L-alanine Maillard model systems (Yaylayan and Keyhani, 1999). 2,3-Butanedione was also identified in mussel extract and suggested to contribute to the desired flavor of cooked mussels (Le Guen et al., 2000) and as a predominant odorant in the headspace of fish sauce (Lapsongphon et al., 2015). Storage of not blanched larvae led to lowering of 2,3-butanedione emission despite the temperature and a gradual increase from the 5th week. A similar increase in 2,3-butanedione content at the end of the studied period was determined for blanched larvae, that can indicate its formation during prolonged larvae storage.
Thus, this study revealed the variations in volatile profiles of lesser mealworms as an effect of storage conditions and blanching. However, it should be taken into account, that further processing insects into food ingredients will also have an effect on flavour formation and sensory characteristics of insect food ingredients (Perez-Santaescolastica et al., 2022). In addition, further development of food products comprises a combination of various food ingredients and processing steps that influence the resulting flavour of a product (Paravisini and Guichard, 2016).
4 Conclusions
Quality characteristics (pH, acid and peroxide values, color) of lesser mealworm larvae were found to be affected by pre-storage heat treatment (i.e. blanching) and storage conditions. The associated changes in volatile profile indicated the course of such reactions as lipid oxidation and lipid hydrolysis throughout seven weeks of storage. The extent of these events was primarily affected by blanching treatment and storage time, and least by storage temperature. A combination of blanching treatment and storage at −80 °C generally retards and inhibits the formation of oxidation-related volatiles during the storage and prevents lowering pH, increasing acid and peroxide values, as well as preserves the colour. Thus, blanching treatment and low-temperature storage can be recommended for the preservation of lesser mealworm larvae quality, while the storage time is important to take into consideration especially when storing lesser mealworm larvae at −20 °C. Moreover, analysis of insect volatiles can provide an indication of undergoing chemical reactions in lesser mealworms larvae and can be an instrument for fast screening of the quality of insect biomass.
Corresponding author; e-mail: maryia.mishyna@wur.nl
These authors contributed equally to this work.
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.26554471
Conflict of interest
The authors have no conflict of interest to declare.
References
Abuelnnor, N., Jones, P., Ratcliffe, N.M., de Lacy Costello, B. and Spencer-Phillips, P.T., 2010. Investigation of the semiochemicals of confused flour beetle Tribolium confusum Jaquelin du Val and grain weevil Sitophilus granarius (L.) in stored wheat grain and flour. In: Proceedings of the 10th International Working Conference on Stored Product Protection, 27 June to 2 July 2010 Estoril, Portugal. https://doi.org/10.5073/jka.2010.425.089
Adámková, A., Kourimská, L., Borkovcová, M., Kulma, M. and Mlček, J., 2016. Nutritional values of edible Coleoptera (Tenebrio molitor, Zophobas morio and Alphitobius diaperinus) reared in the Czech Republic. Potravinarstvo 10: 663-671. https://doi.org/10.5219/609
Bligh, E.G. and Dyer, W.J., 1959. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37: 911-917.
Cabot, M.C. and Lumb, R.H., 1981. The activity of a low temperature lipase in the larvae of Sarcophaga bullata (Diptera: Sarcophagidae). Comparative Biochemistry and Physiology Part B: Comparative Biochemistry 68: 325-328. https://doi.org/10.1016/0305-0491(81)90106-1
Cacchiarelli, C., Fratini, F., Puccini, M., Vitolo, S., Paci, G. and Mancini, S., 2022. Effects of different blanching treatments on colour and microbiological profile of Tenebrio molitor and Zophobas morio larvae. LWT 157: 113112. https://doi.org/10.1016/j.lwt.2022.113112
Costello, B. de L., Wieczorek, M.N. and Drabinska, N., 2023. Editorial: The use of volatile compounds analysis for the assessment of food and beverage quality. Frontiers in Nutrition 10. https://doi.org/10.3389/fnut.2023.1250634
Graham, L.A., Liou, Y.C., Walker, V.K. and Davies, P.L., 1997. Hyperactive antifreeze protein from beetles. Nature 388: 727-728. https://doi.org/10.1038/41908
Grebenteuch, S., Kroh, L.W., Drusch, S. and Rohn, S., 2021. Formation of secondary and tertiary volatile compounds resulting from the lipid oxidation of rapeseed oil. Foods 10: 2417. https://doi.org/10.3390/foods10102417
Guo, X., Lu, S., Wang, Y., Dong, J., Ji, H. and Wang, Q., 2019. Correlations among flavor compounds, lipid oxidation indices, and endogenous enzyme activity during the processing of Xinjiang dry-cured mutton ham. Journal of Food Processing and Preservation 43: 1-15. https://doi.org/10.1111/jfpp.14199
Hidalgo, F.J. and Zamora, R., 2016. Amino acid degradations produced by lipid oxidation products. Critical Reviews in Food Science and Nutrition 56: 1242-1252. https://doi.org/10.1080/10408398.2012.761173
Janssen, R.H., Canelli, G., Sanders, M.G., Bakx, E.J., Lakemond, C.M.M., Fogliano, V. and Vincken, J.-P., 2019. Iron-polyphenol complexes cause blackening upon grinding Hermetia illucens (black soldier fly) larvae. Scientific Reports 9: 2967. https://doi.org/10.1038/s41598-019-38923-x
Janssen, R.H., Lakemond, C.M.M., Fogliano, V., Renzone, G., Scaloni, A. and Vincken, J.P., 2017. Involvement of phenoloxidase in browning during grinding of Tenebrio molitor larvae. PLOS ONE 12: 1-14. https://doi.org/10.1371/journal.pone.0189685
Jeon, Y.-H., Son, Y.-J., Kim, S.-H., Yun, E.-Y., Kang, H.-J. and Hwang, I.-K., 2016. Physicochemical properties and oxidative stabilities of mealworm (Tenebrio molitor) oils under different roasting conditions. Food Science and Biotechnology 25: 105-110. https://doi.org/10.1007/s10068-016-0015-9
Khatun, H., Claes, J., Smets, R., De Winne, A., Akhtaruzzaman, M. and Van Der Borght, M., 2021. Characterization of freeze-dried, oven-dried and blanched house crickets (Acheta domesticus) and Jamaican field crickets (Gryllus assimilis) by means of their physicochemical properties and volatile compounds. European Food Research and Technology 247: 1291-1305. https://doi.org/10.1007/s00217-021-03709-x
Kiatbenjakul, P., Intarapichet, K. and Cadwallader, K.R., 2015. Characterization of potent odorants in male giant water bug (Lethocerus indicus Lep. and Serv.), an important edible insect of Southeast Asia. Food Chemistry 168: 639-647. https://doi.org/10.1016/j.foodchem.2014.07.108
Klunder, H.C., Wolkers-Rooijackers, J., Korpela, J.M. and Nout, M.J.R., 2012. Microbiological aspects of processing and storage of edible insects. Food Control 26: 628-631. https://doi.org/10.1016/J.FOODCONT.2012.02.013
Krajicek, J., Havlikova, M., Bursova, M., Ston, M., Cabala, R., Exnerova, A., Stys, P. and Bosakova, Z., 2016. Comparative analysis of volatile defensive secretions of three species of Pyrrhocoridae (Insecta: Heteroptera) by gas chromatography-mass spectrometric method. PLOS ONE 11: e0168827. https://doi.org/10.1371/journal.pone.0168827
Kröncke, N., Grebenteuch, S., Keil, C., Demtröder, S., Kroh, L., Thünemann, A.F., Benning, R. and Haase, H., 2019. Effect of different drying methods on nutrient quality of the yellow mealworm (Tenebrio molitor L.). Insects 10: 1-13. https://doi.org/10.3390/insects10040084
Kunyaboon, S., Thumanu, K., Park, J.W., Khongla, C. and Yongsawatdigul, J., 2021. Evaluation of lipid oxidation, volatile compounds and vibrational spectroscopy of silver carp (Hypophthalmichthys molitrix) during ice storage as related to the quality of its washed mince. Foods 10. https://doi.org/10.3390/foods10030495
Lapsongphon, N., Yongsawatdigul, J. and Cadwallader, K.R., 2015. Identification and characterization of the aroma-impact components of Thai fish sauce. Journal of Agricultural and Food Chemistry 63: 2628-2638. https://doi.org/10.1021/jf5061248
Larouche, J., Deschamps, M.H., Saucier, L., Lebeuf, Y., Doyen, A. and Vandenberg, G.W., 2019. Effects of killing methods on lipid oxidation, colour and microbial load of black soldier fly (Hermetia illucens) larvae. Animals 9. https://doi.org/10.3390/ani9040182
Le Guen, S., Prost, C. and Demaimay, M., 2000. Characterization of odorant compounds of mussels (Mytilus edulis) according to their origin using gas chromatography-olfactometry and gas chromatography-mass spectrometry. Journal of Chromatography A 896: 361-371. https://doi.org/10.1016/S0021-9673(00)00729-9
Leygonie, C., Britz, T.J. and Hoffman, L.C., 2012. Impact of freezing and thawing on the quality of meat: review. Meat Science 91: 93-98. https://doi.org/10.1016/j.meatsci.2012.01.013
Mancini, S., Fratini, F., Tuccinardi, T., Turchi, B., Nuvoloni, R. and Paci, G., 2019. Effects of different blanching treatments on microbiological profile and quality of the mealworm (Tenebrio molitor). Journal of Insects as Food and Feed 5: 225-234. https://doi.org/10.3920/JIFF2018.0034
Mishyna, M., Haber, M., Benjamin, O., Martinez, J.J.I. and Chen, J., 2020. Drying methods differentially alter volatile profiles of edible locusts and silkworms. Journal of Insects as Food and Feed 6: 405-415. https://doi.org/10.3920/jiff2019.0046
Mishyna, M., Keppler, J.K. and Chen, J., 2021. Techno-functional properties of edible insect proteins and effects of processing. Current Opinion in Colloid & Interface Science 56: 101508. https://doi.org/10.1016/j.cocis.2021.101508
Nunes, C.A., de Souza, V.R., Corrêa, S.C., Silva, M. de C. da C. e, Bastos, S.C. and Pinheiro, A.C.M., 2013. Heating on the volatile composition and sensory aspects of extra-virgin olive oil. Ciência e Agrotecnologia 37: 566-572. https://doi.org/10.1590/s1413-70542013000600010
Paravisini, L. and Guichard, E., 2016. Interactions between aroma compounds and food matrix. In: Guichard, E., Salles, C., Morzel, M. and Le Bon, A.-M. (eds.) Flavour: from food to perception. John Wiley & Sons, Inc., Hoboken, NJ, USA, pp. 208-234. https://doi.org/10.1002/9781118929384.ch9
Pathare, P.B., Opara, U.L. and Al-Said, F.A.J., 2013. Colour measurement and analysis in fresh and processed foods: a review. Food and Bioprocess Technology 6: 36-60. https://doi.org/10.1007/s11947-012-0867-9
Perez-Santaescolastica, C., De Winne, A., Devaere, J. and Fraeye, I., 2022. The flavour of edible insects: a comprehensive review on volatile compounds and their analytical assessment. Trends in Food Science & Technology 127: 352-367. https://doi.org/10.1016/j.tifs.2022.07.011
Pinu, F.R. and Villas-Boas, S.G., 2017. Rapid quantification of major volatile metabolites in fermented food and beverages using gas chromatography-mass spectrometry. Metabolites 7: 1-13. https://doi.org/10.3390/metabo7030037
Rainer Cremer, D. and Eichner, K., 2000. The reaction kinetics for the formation of Strecker aldehydes in low moisture model systems and in plant powders. Food Chemistry 71: 37-43. https://doi.org/10.1016/S0308-8146(00)00122-9
Reyes De Corcuera, J., Cavalieri, R. and Powers, J., 2004. Blanching of foods. In: Heldman, D.R. (ed.) Encyclopedia of agricultural, food and biological engineering. Marcel Dekker Inc., New York, USA. https://doi.org/10.1081/E-EAFE-120030417
Ribeiro, J.C., Marques, J.P., Fernandes, T.R., Pintado, M.E., Carvalho, S.M.P. and Cunha, L.M., 2024. Effect of blanching, storage and drying conditions on the macro-composition, color and safety of mealworm Tenebrio molitor larvae. LWT 191: 115646. https://doi.org/10.1016/j.lwt.2023.115646
Rochat, D., Ramirez-Lucas, P., Malosse, C., Aldana, R., Kakul, T. and Morin, J.P., 2000. Role of solid-phase microextraction in the identification of highly volatile pheromones of two Rhinoceros beetles Scapanes australis and Strategus aloeus (Coleoptera, Scarabaeidae, Dynastinae). Journal of Chromatography A 885: 433-444. https://doi.org/10.1016/S0021-9673(99)01049-3
Ross, C.F. and Smith, D.M., 2006. Use of volatiles as indicators of lipid oxidation in muscle foods. Comprehensive Reviews in Food Science and Food Safety 5: 18-25. https://doi.org/10.1111/j.1541-4337.2006.tb00077.x
Rumpold, B.A. and Schlüter, O.K., 2013. Nutritional composition and safety aspects of edible insects. Molecular Nutrition & Food Research 57: 802-823. https://doi.org/10.1002/mnfr.201200735
Šanda, M., Žáček, P., Streinz, L., Dračı́nský, M. and Koutek, B., 2012. Profiling and characterization of volatile secretions from the European stink bug Graphosoma lineatum (Heteroptera: Pentatomidae) by two-dimensional gas chromatography/time-of-flight mass spectrometry. Journal of Chromatography B 881-882: 69-75. https://doi.org/10.1016/j.jchromb.2011.11.043
Tiencheu, B., Womeni, H.M., Linder, M., Mbiapo, F.T., Villeneuve, P., Fanni, J. and Parmentier, M., 2013. Changes of lipids in insect (Rhynchophorus phoenicis) during cooking and storage. European Journal of Lipid Science and Technology 115: 186-195. https://doi.org/10.1002/ejlt.201200284
Turck, D., Castenmiller, J., De Henauw, S., Hirsch-Ernst, K.I., Kearney, J., Maciuk, A., Mangelsdorf, I., McArdle, H.J., Naska, A., Pelaez, C., Pentieva, K., Siani, A., Thies, F., Tsabouri, S., Vinceti, M., Cubadda, F., Frenzel, T., Heinonen, M., Marchelli, R., Neuhäuser-Berthold, M., Poulsen, M., Prieto Maradona, M., Schlatter, J.R., van Loveren, H., Ververis, E. and Knutsen, H.K., 2021. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA Journal 19: 1-29. https://doi.org/10.2903/j.efsa.2021.6343
Tzompa-Sosa, D.A., Yi, L., van Valenberg, H.J.F. and Lakemond, C.M.M., 2019. Four insect oils as food ingredient: physical and chemical characterisation of insect oils obtained by an aqueous oil extraction. Journal of Insects as Food and Feed 1: 1-14. https://doi.org/10.3920/jiff2018.0020
van Huis, A. and Oonincx, D.G.A.B., 2017. The environmental sustainability of insects as food and feed. A review. Agronomy for Sustainable Development 37: 43. https://doi.org/10.1007/s13593-017-0452-8
Vandeweyer, D., Lenaerts, S., Callens, A. and Van Campenhout, L., 2017. Effect of blanching followed by refrigerated storage or industrial microwave drying on the microbial load of yellow mealworm larvae (Tenebrio molitor). Food Control 71: 311-314. https://doi.org/10.1016/j.foodcont.2016.07.011
Wessels, M.L.J., Azzollini, D. and Fogliano, V., 2020. Frozen storage of lesser mealworm larvae (Alphitobius diaperinus) changes chemical properties and functionalities of the derived ingredients. Food Chemistry 320: 126649. https://doi.org/10.1016/j.foodchem.2020.126649
Wu, X., Zhang, M., Bhandari, B. and Li, Z., 2019. Effect of blanching on volatile compounds and structural aspects of Cordyceps militaris dried by microwave-assisted pulse-spouted bed freeze-drying (MPSFD). Drying Technology 37: 13-25. https://doi.org/10.1080/07373937.2018.1433685
Wynants, E., Crauwels, S., Verreth, C., Gianotten, N., Lievens, B., Claes, J. and Van Campenhout, L., 2018. Microbial dynamics during production of lesser mealworms (Alphitobius diaperinus) for human consumption at industrial scale. Food Microbiology 70: 181-191. https://doi.org/10.1016/J.FM.2017.09.012
Yaylayan, V. and Keyhani, A., 1999. Origin of 2,3-pentanedione and 2,3-butanedione in D-glucose/L-alanine Maillard model systems. Journal of Agricultural and Food Chemistry 47: 3280-3284. https://doi.org/10.1021/jf9902292
Yeo, H., Youn, K., Kim, M., Yun, E., Hwang, J., Jeong, W. and Jun, M., 2013. Fatty acid composition and volatile constituents of Protaetia brevitarsis larvae. Preventive Nutrition and Food Science 18: 150-156. https://doi.org/10.3746/pnf.2013.18.2.150




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}