
Abstract
Lab-grown insect meat is a promising alternative to traditional livestock for sustainable food production. This review paper aims to provide a comprehensive overview of the current state of knowledge regarding lab-grown insect meat, emphasizing key aspects such as life cycle assessment, insect cell culture history, technological advancements, and bio-robotics in insect cell culture. Comparisons and challenges between insect and mammalian/avian cell culture methodologies are presented. The nutritional content of edible insects (proximate, amino acid, mineral, and vitamin content) and the potential health benefits of consuming insect meat are discussed. The paper also explores embryonic and adult myogenesis processes in insect cells, as well as the significance of insect body fat and muscle cells in culture. Applications of insect cell culture in various fields, such as food production and pharmaceutical development, are presented. Moreover, the potential occurrence of mutations in lab-grown insect cells is examined. Lastly, the review addresses the drawbacks and limitations of insect labriculture, discussing factors such as scalability, cost-efficiency, and public acceptance. Overall, this comprehensive review provides essential insights into the chemical and biological aspects of lab-grown insect meat, paving the way for further research and development in this emerging field. This article is the first review article reporting the chemical and biological insights of lab-grown insect meat.

Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
1 Introduction
As the global human population reaching more than 8.0 billion in 2022 (United Nation, 2023), the need for food worldwide is increasing, including the demand for meat. It is estimated that by 2050, the global consumption of meat will reach an astounding 460 to 570 million tons (Mendoza, 2023). However, traditional meat production may struggle to meet this demand sustainably. Nevertheless, traditional methods of meat production, especially those involving livestock, present substantial environmental, ethical, and public health issues (Bryant and Barnett, 2020). Livestock rearing for meat production is claimed as a significant contributor to greenhouse gas emissions, deforestation, water pollution, and habitat destruction (Adedibu, 2023). In response to these challenges, sustainable alternatives for meat have emerged, and one compelling innovation in this regard is lab-grown meat. Sustainable alternatives for meat are options to get meat that is produced in an environmentally friendly manner, including lower carbon emissions, water consumption, and other factors compared to meat produced from conventional livestock. Lab-grown meat offers a potential solution by reducing the environmental footprint associated with traditional animal agriculture (Smetana et al., 2023).
There are more than 2,000 known species of edible insects worldwide, which makes them an excellent alternative protein source for humans (van Huis, 2020). Among those, insects such as beetles, crickets, and mealworms have cells that are more easily genetically modified compared to animal cells, making them a suitable option for lab-grown insect meat (Kumar et al., 2022). This allows scientists to manipulate the genetic code of insects more easily, enabling the production of desired traits or enhancing nutritional profiles. Insects typically have a simpler genetic structure compared to mammals. They have fewer chromosomes and a more straightforward genome, making it easier to manipulate and introduce genetic modifications (Reid and OâBrochta, 2016). Insect cells are relatively easy to culture in the laboratory, which is crucial for genetic modification experiments. They can be grown in large quantities and maintained under controlled conditions, making it convenient for researchers to work with them. Insect cells often exhibit a higher propensity for homologous recombination, a process that allows for precise genetic modifications (Simoni et al., 2014). Homologous recombination involves the exchange of genetic material between similar DNA sequences, facilitating targeted gene editing. In addition to that, insect cells can be more efficiently transfected with foreign DNA compared to some animal cells (Dong et al., 2016).
Working with insect cells may involve fewer ethical and regulatory hurdles compared to animal experimentation, particularly for researchers conducting genetic modification studies. In some cases, the biology of insects may provide specific advantages for certain genetic modification applications. For instance, armyworm Spodoptera frugiperda are particularly well-suited for producing recombinant proteins, making them valuable for biotechnology and pharmaceutical applications (Park et al., 2022). In addition to that, insects are generally have a shorter lifespan and higher reproductive rates compared to the traditional livestock animals. This means that insect cells can multiply and generate biomass more rapidly, potentially increasing the efficiency of lab-grown meat production, which then allows for faster analysis of the modified traits. Furthermore, the entire life cycle of insects is highly suitable for lab-grown meat production. Insects, such as crickets, mealworms, and grasshoppers, go through complete metamorphosis, which involves distinct developmental stages. These distinct stages make it feasible to manipulate and control the breeding and growth processes in a lab setting (Jensen et al., 2017).
In recent years, there has been a growing interest in using insect cells to produce proteins for therapeutic purposes. These proteins are used to treat diseases like cancer, genetic disorders, and autoimmune diseases (Dambuza et al., 2017). The challenge lies in producing these proteins in large quantities and at reasonable costs. Traditionally, mammalian cells have been used for this purpose due to their ability to correctly fold and modify complex human proteins. However, using mammalian cells is expensive and requires strict environmental control. Insect cells, specifically those harvested from the fall armyworm and cabbage looper, have been studied as a more efficient and cost-effective alternative (Skora et al., 2015). Insect cells offer advantages such as high protein yields, correct folding and modifications, simplified culture process, reduced costs, and scalability (Gomes et al., 2016). In this review, the emphasis is on lab-grown cultures of insect cells. Cell culture is the technique of cultivating insect cells in a favorable artificial environment. The cells might be obtained from multicellular eukaryotes, established cell lines, or established cell strains. Insect cells can be utilized to produce cell lines. Cell lines that can develop in suspension are recommended because of their quicker growth rate (Verma et al., 2020). Insect cell-based models have been shown to be beneficial for activity evaluation not only because of their cheap cost-benefit ratio, ease of maintenance, repeatability, and potential for application in high-throughput screening, but also because they do not pose bioethical concerns (He et al., 2023).
While the consumption of lab-grown meat is still debated as it involves the recreation of animal tissue in a laboratory setting, the existence of edible insects challenges this perception. Lab-grown meat raises ethical questions, as it blurs the line between what is natural and what is artificially created (Liu et al., 2022). Additionally, some individuals may find the concept of eating meat grown in a lab unfamiliar and perceive it as unnatural or even unnerving. On the other hand, the consumption of edible insects has been part of the human diet for centuries in certain cultures and regions (Anankware et al., 2015). Insects are hailed as a highly sustainable protein source, rich in nutrients and can provide a valuable source of vitamins and minerals (Payne et al., 2019). They also require considerably less water, land, and feed compared to traditional livestock, making them ecologically friendly (Van Huis and Oonincx, 2017). Lab-grown insect meat involves the production of edible insect tissue in a controlled laboratory environment, free from the intensive resource requirements and ethical concerns associated with traditional animal farming. This technology harnesses the remarkable nutritional profile and efficiency of insects as a protein source while minimizing the ecological footprint and welfare concerns typically associated with livestock production. This comprehensive review aims to provide a detailed information of lab-grown insect meat, focusing on the chemical and biological aspects. By exploring the underlying principles of insect cell culture, nutrient composition, and technology advancements, this research paper aims to shed light on the potential of this technology to revolutionize the meat industry. The findings of this review will contribute to a better understanding of lab-grown insect meat, offering insights into its potential as a sustainable alternative to conventional meat production. As lab-grown insect meat continues to attract attention from researchers, environmentalists, and policymakers, this comprehensive review is a timely and critical resource to inform future developments and discussions in the field.
2 Insect cell culture
The high-quality insect production underpins the viability and sustainability of insects as a valuable source of protein and other essential nutrients. Thus, optimizing the nutritional composition of the insectâs diet is crucial to enhance its protein content and biomass production. The composition and nutritional value of the feed directly impact the growth, development, and overall health of the insects being reared (Hawkey et al., 2021), which, in turn, determines the quality of the final insect product. The choice of feed ingredients, their nutrient content, and the feeding regimen are critical factors in ensuring optimal insect growth and nutritional profile. High-quality feed sources rich in essential nutrients such as proteins, fats, vitamins, and minerals are essential for achieving desirable insect biomass and nutritional content (Pinotti and Ottoboni, 2021). Furthermore, the feed can influence not only the quantity but also the nutritional composition of the harvested insects. By manipulating the feed composition, it is possible to modify the macronutrient and micronutrient profiles of the insects (Scala et al., 2020), making them more suitable for specific applications, such as animal feed, human consumption, or pharmaceutical purposes.
Insect cell culture can be used to develop alternative meat production methods. Laboratory-grown meat can help address food security issues by providing a scalable and efficient meat production method (Cowan et al., 2022). Their innovative products typically fall into the âcell-basedâ or âculturedâ meat category. Unlike traditional meat production, which involves raising and slaughtering animals, Upside Foodsâ approach utilizes cellular farming techniques to produce meat directly from animal cells in a controlled environment. GOOD Meat, like other companies in the cultured meat industry, is dedicated to developing and producing ethical and sustainable meat alternatives. The first idea of lab-grown meat dates back to the early 20th century, when Winston Churchill speculated about the possibility of growing animal tissue in a lab (Treich, 2021). However, it was only in the 2000s that significant progress was made towards creating a practical technique for growing meat in vitro (Bhat et al., 2017). This laid the foundation for future research and development in the field. There are now over 600 documented cell lines from insects comprising the orders Diptera, Hemiptera, and Lepidoptera for insect cell culture. Frequently, these cultures are used in molecular biology studies to make recombinant proteins that can be used in biopharmaceuticals, vaccines, and diagnostics (Ghosh et al., 2020). For the production of meat derived from insects, the process of insect cell culture involves extracting cells from insects and culturing them in a nutrient-rich medium that provides the necessary nutrients and growth factors for cell proliferation. The cells are developed on a scaffold that resembles the texture and structure of actual meat, enabling them to develop into muscle fibers to produce a meat-like product (Zhang et al., 2022b). Insect cell culture for meat has tremendous potential advantages, such as higher food security, as it has been reported to demonstrate nutritional profiles, flavors, and tastes comparable to those of meat obtained from conventionally raised animals (Lynn, 2002; Murphy and Piwnica-Worms, 1994). Figure 1 shows a schematic representation of laboratory culturing of insect cells. This recent interest arises as insect cells present unique advantages, including rapid growth rates, easier genetic modifications, resource efficiency, high reproduction potential, and their ability to survive in suboptimal conditions (Rubio et al., 2019a), making them a promising alternative to conventional meat production.
Schematic representation of insect cell culture. (1) Isolation of embryonic cells, (2) Separation of fat and muscle cells, (3) Incubation of cells at 20-30 °C, (4) Supply of nutrient broth (with/without serum), (5) Adherent/adaptation to suspension culture, (6) Scaling up strategies in larger vessels, (7) Harvesting of multiple cell types, (8) Seeding/scaffolding of cells, and (9) Differentiation of cells.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
The development of automated systems for insect cell culture can make significant use of technology in bio-robotics. These systems utilize robotics, advanced imaging, and machine learning algorithms to ensure precise and consistent handling of cells. Automated systems can significantly reduce human error, increase productivity, and enhance reproducibility. They are capable of performing complex tasks, such as cell sorting, cell counting, media exchange, and subculture, which are essential for large-scale insect cell culture (Ker et al., 2011). As an example, Nguyen et al. (2011) proved that that the usage of an automated robotic clone picking system manage to produced 10-30% higher occlusion body yields than the original cell population. Furthermore, advancements in microfluidics have facilitated the development of miniaturized systems for insect cell culture (Alam et al., 2018). Microfluidics enables precise control and manipulation of fluids at the microliter or even nanolitre scale, allowing researchers to create a highly controlled and optimized environment for insect cells. Microfluidic devices can mimic the physiological microenvironment of cells, including nutrient gradients, oxygen supply, and waste removal, leading to improved cell viability and production efficiency (Torino et al., 2018).
Insect culture media composition
Mediums for growing insect cells typically include a plant-based additive (yeastolate, primatone RL, peptides, hydrolysates, and pluronic lipids), insect cell fluid (haemolymph), and either 5 or 10% foetal bovine serum in addition to a basal medium (Graceâs insect medium, Schneiderâs Drosophila medium, or Eagleâs medium) (The Buzzing Future of Food: An Introduction to Entomoculture | Protein Report, n.d.). Foetal bovine serum offers proteins with antioxidant and detoxifying effects, protease inhibitors, and growth factors to the culture medium. However, serum in cell culture has various downsides, including high cost, lot-to-lot fluctuation, presence of contaminants, and limited supply. As a result of its high protein concentration, it can also impede the purification of recombinant proteins. Such concerns have sparked the quest for serum- and protein-free, and ideally chemically defined media.
Glucose and glutamine are essential nutrients for energy metabolism in the growth of insect cells, with glucose being the most important carbon source (Butler, 2015). Insect cells require amino acids, lipids, cholesterol, and vitamins for proper growth and protein expression, and must be supplemented in the culture medium as the insect cells cannot synthesize them. Lipid supplementation is particularly important as insect cells cannot produce or modify fatty acids on their own, which can lead to cell degeneration. Since vitamins and cholesterol both play crucial roles in membrane production and the control of essential metabolic processes, supplementation of both is necessary. The culture medium also contains other ingredients such as trace elements, yeastolate, and the non-ionic detergent Kolliphor® P188 to prevent cell injury. However, many insect cell cultures currently in the market, including TC100, IPL-41, and Hinkâs TNM-FH, have been adapted from Graceâs medium, developed by Grace in 1962 (Grace, 1962). Commercial suppliers of insect cell media have been working on developing animal-free and chemically defined insect cell media to minimize variability caused by undefined ingredients like yeastolates and ensure more consistent and reliable cell culture results. A study compared different commercial media for making adeno-associated virus (AAV) vectors using the ExpiSf Baculovirus Expression System. The results showed that using a chemically defined culture medium with ExpiSf system gave significantly more AAV yields for three different types of AAV, thus supporting scalable AAV manufacture suitable for preclinical and clinical studies (Kurasawa et al., 2020). Thermo Fisher Scientific has developed several serum-free media namely, ExpiSf CD Medium, Sf-900 III SFM, Sf-900 II SFM, Express Five SFM, for adherent and suspension culture of insect cells in baculovirus expression vector systems (BEVS) (Insect Media â Gibco Insect Cell Culture Media | Thermo Fisher Scientific, IN, n.d.). Expression Systems, another firm, has produced a serum- and protein-free insect cell culture medium called ESF 921, that can support the development and expression of a broad variety of insect cells, such as Sf9, Sf21, and Drosophila S2 (ESF 921 Insect Cell Culture Medium, Protein Free â Expression Systems, n.d.). Supplementary Table S2 enlists different insect cell culture media manufactured by different companies.
Scaffolding/cell substrate
Scaffolds are extensively employed in insect cell culture as a support system to generate a 3D framework in which cells can grow and develop. Scaffolds can improve cell adhesion, proliferation, and functioning by creating a microenvironment that is similar to the native extracellular matrix (ECM) (Bomkamp et al., 2022; Campuzano and Pelling, 2019). Thus, collagen and other polymers derived from animals are frequently used as a benchmark. However, concerns regarding ethics, the environment, and variability make their reliance unattractive. When designing a scaffold intended for meat consumption, it must meet certain criteria. Firstly, it should not contain any animal components, as the goal is to produce meat/meat-like product without relying on traditional animal agriculture. Additionally, it should be inexpensive and readily available to ensure scalability and affordability. Another important consideration is that the scaffold should be edible, as it will be consumed along with the cultured meat. Furthermore, similar to scaffolds used in biomedical applications, the structure of the scaffold should provide a large surface area for the cells to adhere to. This promotes cell growth and tissue formation. Additionally, the scaffold should mimic the mechanical properties of the target tissues, ensuring that the resulting cultured meat has the desired texture and consistency (Campuzano and Pelling, 2019).
Scaffolds that are consistent, chemically defined, and inexpensive have been in demand, and studies of animal-free scaffolds such as cellulose, chitin, chitosan extracted from fungi such as mushrooms, alginate, recombinant silk, polylactic acid (PLA), and polyglycolic acid (PGA) have emerged as a promising source (Kraham, 2017). For instance, flexible polydimethylsiloxane (PDMS) moulds used to support cultures of Manduca sexta muscle cell and Ctenoplusia agnate vessel tissue, provides a flexible surface that can move with the muscle contractions and functions as a guide to help the cells arrange themselves (Akiyama et al., 2013; Baryshyan et al., 2014). Using chitin- or chitosan-based scaffolds for cell-cultured meats could address the challenge of scalability and enhance their nutritional and health advantages. Research indicates that these scaffolds exhibit antioxidant properties, hinder the growth of harmful microorganisms, lower cholesterol levels, expedite wound healing, and provide neuroprotection (Chitin: Providing Structure and Biofunction to Cell-Cultured Meat, n.d.). Chitosan extracted from mushrooms has potential as a biomaterial for scaffold production in biomedical engineering, as it is âgenerally recognized as safe (GRAS)â by the US FDA (Chinova Bioworks Self-Affirms GRAS for Natural Chitosan Preservative, n.d.). It is particularly relevant to insect cell culture since it is derived from chitin, a major component of their exoskeletons (Jana et al., 2013). In general, the materials suitable for scaffolding in insect cell culture should possess certain characteristics such as biocompatibility, porous structure to facilitate nutrient and oxygen exchange as well as waste removal, adequate mechanical strength to provide structural support, and must be biodegradable and promote cell adhesion. Supplementary Table S3 lists a number of other scaffolds that find frequent application in the field of tissue engineering. Limited research has been conducted to specifically evaluate materials for insect-based tissue engineering. Hence, it is crucial to explore and enhance scaffold properties to determine their ability to support adhesion, proliferation, survival, and differentiation of insect muscle and fat cells.
Growth promoters
Incorporation of a variety of potential growth inducing compounds as feed additives in insect cell culture is of particular importance for the optimum maintenance of cell growth. Polyamines, namely, spermine, putrescine, and spermidine, as well as their precursor ornithine, have all been shown to be required for chemically defined cell proliferation in a variety of cell lines (Mitsuhashi, 1998). Adequate levels of polyamines, particularly spermidine, have been shown to promote DNA synthesis in Drosophila cell lines Cl.8 and S2 (Burnette et al., 2014). Additionally, maintaining proper polyamine levels has been found to regulate cell cycle genes and improve cell cycle progression in the BmN cell line from the silkworm (Chang et al., 2021). These findings highlight the importance of polyamines for cellular growth and the regulation of key cellular processes in insect cell lines. Betaine has been studied for its role as an osmoprotectant, which helps cells maintain their viability and integrity under osmotic stress (Arumugam et al., 2021). In a patent granted in 2010, it was claimed that betaine can boost insect cell viability and polypeptide synthesis under normal osmotic circumstances (US7829309B1 â Cell Culture Performance with Betaine â Google Patents, n.d.). Additionally, when the gene responsible for producing the AGXTLTM protein was inserted into insect cells at 27 °C, the addition of betaine facilitated the production of a significant amount of the AGXTLTM protein in a soluble form (Santana et al., 2003). Further, hydrolysate extract added to culture media contains significant levels of nucleosides. Among these nucleic compounds, adenosine, uridine, and inosine were selected as primary growth-inducing compounds. The inclusion of nucleosides in the culture media provides the cells with reservoirs of these compounds, potentially minimizing growth delays that occur after cell passaging (Boegel, n.d.; Quattrociocchi, 2017). Sodium ascorbate serves as an antioxidant, which is particularly crucial in a trace metal enriched medium. Ascorbic acid, also known as vitamin C, is believed to have a multifaceted role in cell metabolism and is likely to be a significant component in chemically defined insect cell media (Wang et al., 2001). Further, the importance of both ethanolamine and selenium for growth has been demonstrated in numerous serum-free media for mammalian cell culture, its necessity in insect cell culture has not been extensively studied (Yang et al., 2016). In serum-free medium, where the cells are not supplemented with serum containing growth factors, the autocrine regulation of cell proliferation becomes particularly important. Metalloproteinases from Trichoplusia ni High Five and BTI-Tn-5B1-4 cell lines, can play a crucial role in providing the necessary growth factors for cell growth and proliferation by autocrine regulation (Eriksson et al., 2005). There is also a documented resemblance between insect prothoracicotropic hormone (PTTH) and insulin, suggesting a possible functional similarity or connection in cell growth regulation. In insect cell cultures, insulin plays a key role as a growth factor that promotes cell proliferation and viability (Nagaraj et al., 2022; Svensson, n.d.). Table 1 enlists growth regulating compounds which may be used when culturing insect cells.
Possible growth promoters in insect cell culture
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Bioreactors
Insect cells are commonly cultured in fixed systems like culture vessels, but they can also be adjusted to grow in suspension conditions in shaker flasks and bioreactors with 1000-L capacity (Pamboukian et al., 2008). When insect cells are grown in stationary monolayer cultures, the surface area to volume ratio decreases as the culture expands, which can cause challenges when trying to produce cells on a large scale efficiently (Gotoh et al., 2004). Sophisticated bioreactor systems, including rotating wall, airlift, perfusion, and semi-continuous reactors, have been studied alongside conventional batch bioreactors for applications in insect cell culture for the synthesis of recombinant proteins. These systems typically maintain a high level of hygiene to ensure optimal cell growth and minimize the risk of contamination (Kaiser et al., 2022). Perfusion reactors might be used during the muscle differentiation stages, when the cells would be adherent to a substrate as multinucleated myofibers, whereas simple bioreactor may be effective during the cell proliferation phase. Further, in large-scale bioreactors, oxygen is typically supplied by sparging, which involves bubbling oxygen into the culture medium. However, for most insect cell culture systems, surface aeration is sufficient and provides enough oxygen for cell growth. In addition, to reduce shear sensitivity in airlift fermentation systems, surfactants like Pluronic F-68 can be added which can help strengthen the cells, thus minimizing the damage caused by mechanical forces (Strobl et al., 2021). Furthermore, insect bioreactor systems, can yield cell density levels ranging in between 2 to 8 à 106 cells/mL, which is considerably greater than mammalian cells. Perfusion systems may further boost this productivity and yield densities up to 3.5 à 107 cells/mL (Imseng et al., 2014). Figure 2 shows a diagrammatic representation of in vitro cultured insect meat in a bioreactor.
Production of insect-based cultured meat in a bioreactor.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
3 Nutritional content of edible insects
One of the most significant advantages of edible insects is its high protein content (Supplementary Table S5). Insects are often containing more protein per gram than traditional meat sources. As examples, Allomyrina dichotoma (Coleoptera: Dynastidae), Protaetia brevitarsis (Coleoptera: Cetoniidae), Tenebrio molitor (Coleoptera: Tenebrionidae), Teleogryllus emma (Orthoptera: Gryllidae), and Gryllus bimaculatus (Ortoptera: Gryllidae) are reported contained higher amounts of protein compared to conventional animal meats and chicken eggs (Ghosh et al., 2017). Even though the protein content can vary significantly among different insect species, but it generally ranges from 35% to 80% of their dry weight (KourÌimskaÌ and AdaÌmkovaÌ, 2016). In addition, protein content for chicken and beef around 20.85-72.43% (Li et al., 2021). So, with the protein content of insects, which is not that far from the protein content of chicken and beef, it can be an alternative protein source. This makes these insects an excellent protein source for both humans and animals. The high protein content of insects is particularly significant in the context of global food security. In addition to that, edible insects also contain fat, which can range from 10% to 50% of their dry weight (KourÌimskaÌ and AdaÌmkovaÌ, 2016). While the fat content in insect meat may seem high, it is important to note that insect fats are predominantly healthy unsaturated fats, including monounsaturated and polyunsaturated fats (Mohamed, 2015). Lee et al. (2008) reported that chitin was antivirally active against tumorigenesis. Furthermore, insectsâ fatty acid profiles (Table 2) are diverse and include essential fatty acids such as omega-3 and omega-6 (Paul et al., 2017). Edible insects also offer a well-balanced amino acid profile (Supplementary Table S6). Amino acids are the building blocks of proteins and play crucial roles in various physiological processes, including muscle development, immune function, and neurotransmitter synthesis (Dimou et al., 2022). Some insects have a higher protein ratio than beef, hog, chicken, and lamb (Akhtar and Isman, 2018), with values ranging from 13% to 77% of dry matter. For example, the protein contents of the top edible insect orders on the basis of dry matter are reported as Rhynchophorus phoenicis (50.01%), Allomyrina dichotoma (54.18%), Ruspolia differens (44.59%), Aeschna multicolor (54.24%), Teleogryllus emma (55.65%), Gynanisa maja (55.92%), and Gryllus bimaculatus (58.32%) (Kim et al., 2019).
Fatty acid composition of some common edible insects
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Further research is needed to better understand the protein content, amino acid composition, and digestibility of cultured meat, as well as the factors influencing these parameters (Parodi et al., 2019). Although the protein content of in vitro meat has not been quantified, morphological observations suggest that current culturing methods produce meat with cytoskeletal proteins similar to traditional meat (ZidaricÌ et al., 2020). Current research focuses on optimizing and adjusting the nutritional content of the medium used to grow protein-rich cells. However, greater explanation is required about the flow of nutrients to growing cells and the changes that occur in retrieved cells (tissue) (Miller, 2020). These scaffolds can be protein-based (e.g. collagen), and when used as a component in the final product, they can have a direct impact on the amino acid composition (Ianovici et al., 2022). Animal-derived polymers, such as collagen, are thought to be well-suited for cultured meat production, though the essential amino profile of the in vitro meat may suffer as collagen contains a higher amount of non-essential amino acid glycine (Ahmad et al., 2021). However, the extent to which different scaffolds alter the amino acid composition of in vitro meat has not yet been investigated. Edible insects are also rich in minerals (Supplementary Table S7) and essential vitamins (Supplementary Table S8), making them a valuable addition to the diet. However, the specific micronutrient content can vary among different insect species. Crickets contain levels of iron comparable to or even higher than traditional meat sources (Nowakowski et al., 2022). Iron is crucial for maintaining healthy blood and preventing anemia (Miller, 2013). Thus, incorporating insects into the diet can be particularly beneficial for individuals with iron-deficiency anemia. These micronutrients contribute to the overall nutritional value of edible insects, making them a well-rounded source of essential vitamins and minerals.
Comparison of traditional protein feed sources with generic protein feed
Implementing life cycle assessments in insect farming operations can help determine the most optimal protein feed sources, contributing to the development of sustainable and environmentally friendly insect farming practices, thus addressing the challenges of global food security and sustainable protein production. When considering the costs, water usage, and electricity consumption associated with protein feed sources for edible insect farming, the utilization of generic protein feed sources appears to be a more sustainable choice (Nijdam et al., 2012). These sources offer advantages such as lower costs, reduced water usage, and decreased electricity consumption compared to traditional protein feed sources. Smetana et al. (2016) reported that the production of insect-based protein powder and meat substitute, based on food by-products, is 2-5 times more environmentally beneficial than that of traditional products. In addition to that, traditional protein feed sources, such as soybean meal and fishmeal, are known to be expensive due to high demand and limited availability. Conversely, generic protein feed sources, including plant-based grains and waste byproducts, can be obtained at a relatively lower cost due to their wider availability. Insect farmers can leverage cost savings by utilizing local, abundant, and inexpensive protein feed sources, minimizing financial burdens. Generic protein feeds also provide longer-term cost stability and independence from global commodity price fluctuations.
When comparing traditional and generic protein feed sourcesâ water usage in insect farming, it is important to consider both the feed production process and the water requirements of insects themselves. Traditional protein feed sources, such as soybeans and fish, require substantial irrigation and aquaculture practices, consuming significant amounts of water. On the other hand, generic protein feed sources tend to have lower water requirements, especially when utilizing non-irrigated agricultural residues or byproducts with minimal additional water needs. This reduction in water use contributes to the overall sustainability of edible insect farming systems (Miglietta et al., 2015). Electricity consumption also plays a vital role in the life cycle assessment of protein feed sources for insect farming. Traditional protein feed sources, such as soybean meal and fishmeal, often involve energy-intensive processes like drying, milling, and transportation. These processes require considerable electricity, resulting in a substantial carbon footprint (Bosch et al., 2019). On the contrary, generic protein feed sources usually require less energy during processing, handling, and transportation. Feeding insects with locally sourced crops or organic waste offers a significant advantage in terms of reducing electricity consumption and associated greenhouse gas emissions. Over the years, many studies have been carried out to examine the Life Cycle Assessment (LCA) of insect production for use as food and feed. This assessment is required to evaluate the environmental impacts associated with the entire life cycle of insect-based food products. This assessment considers the inputs (e.g. feed, water, energy) and outputs (e.g. greenhouse gas emissions, waste) associated with the different stages of the insect production process, including breeding, rearing, processing, packaging, distribution, consumption, and waste management (Figure 3).
System boundary for the production of lab grown insect meat.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Comparison of insect cell culture to mammalian and avian cell culture
In recent decades, cell culture has become an indispensable tool in many fields of biology and biotechnology. It allows scientists to study the behavior and properties of cells under controlled laboratory conditions. However, not all cells are created equal. Insect cell culture, in particular, has gained attention and popularity due to its unique characteristics and potential applications. On the other hand, mammalian cell culture is by far the most extensively used form of cell culture due to its relevance to human biology. Mammalian cells, such as human cells, are highly similar in terms of structure, physiology, and genetics. This similarity makes mammalian cell culture a reliable tool for studying the mechanisms of diseases, drug toxicity, and efficacy testing (Doke and Dhawale, 2015). Mammalian cells are also preferred for the production of therapeutic proteins and vaccines due to their ability to process complex post-translational modifications (Dumont et al., 2016). Avian cell culture, on the other hand, offers unique advantages and applications. Avian cells are closer to mammals in terms of evolutionary relationship as both belong to the class Aves. However, avian cell culture has been less widely used compared to mammalian cell culture. Avian cells can serve as a model system for studying viral infections, as birds are susceptible to a wide range of viruses, including avian influenza (Shriner and Root, 2020). Additionally, avian cells are commonly used for vaccine production against avian-specific diseases (Lee et al., 2023).
Positive environmental gains of insect cultured meat
Insect cultured meat presents numerous positive environmental gains that make it a promising alternative to conventional livestock farming. Alexander et al. (2017) and Hadi and Brightwell (2021) reported that one of the most significant environmental gains of insect cultured meat is its reduced carbon footprint compared to conventional meat production. Livestock farming, particularly beef production, is a leading contributor to greenhouse gas emissions, deforestation, and water pollution (Sakadevan and Nguyen, 2017). Insect cultivation, on the other hand, emits fewer greenhouse gases and requires significantly less land, water, and resources. Research by Vauterin et al. (2021) suggest that insect protein has the greatest potential to reduce the carbon footprints of European consumers, if insects are directly consumed as food. In addition to that, it is suggested that not only that the use of insects as animal feed can substantially contribute to the sustainability of broiler production systems with regard to lowering global warming potential, but also that low-value side streams are key for improving this potential. To put it into perspective, producing a kilogram of beef requires around 25 kilograms of feed, 15,000 liters of water, and releases approximately 26 kilograms of greenhouse gas emissions (Vergunst and Savulescu, 2017). In contrast, insect cultured meat utilizes only a fraction of these resources. For instance, cricket farming requires minimal land, water, and feed inputs, producing much lower emissions (Cannon and Britt, 2019). Embracing insect cultured meat could, therefore, alleviate the environmental strain caused by conventional livestock farming and contribute to mitigating climate change.
Moreover, cultivating insects as an alternative protein source can also mitigate deforestation and biodiversity loss. It is estimated that livestock farming is responsible for over 70% of deforestation in the Amazon rainforest alone (Bustamante et al., 2012). As demand for meat rises, more land is cleared for grazing and growing animal feed crops, resulting in habitat destruction and decreased biodiversity. Insect cultured meat production, with its smaller land requirements, could help alleviate this pressure on ecosystems, allowing forests to regrow and habitats to recover. Another notable environmental advantage of insect cultured meat is its efficient use of water resources. Traditional livestock farming often requires substantial amounts of water for animal hydration and feed irrigation. Insects, however, have an impressive water usage efficiency, as they require significantly less water than conventional livestock farming. For example, crickets need approximately 200 times less water than cows per kilogram of protein produced (Alexander, 2022). This reduction in water consumption is especially important in regions where water scarcity is a growing concern, benefiting both the environment and local populations. Furthermore, insect cultured meat also has the potential to minimize the use of harmful pesticides and antibiotics. Conventional livestock farming often relies on large quantities of pesticides to control pests and diseases that can spread in crowded and unhygienic conditions. The overuse of antibiotics is also prevalent in the livestock industry, leading to antibiotic resistance, a grave concern for human health (Ma et al., 2021). In contrast, insect cultivation poses fewer risks in terms of disease transmission, reducing the need for pesticides and antibiotics, and thus preventing their negative impacts on the environment and public health.
Negative environmental impact of insect cultured meat
Even though the concept of insect cultured meat has emerged as a promising alternative to traditional livestock farming, it is crucial to critically examine and explore the various detrimental factors associated with insect cultured meat and argues that caution should be exercised before widely adopting this technology (Raheem et al., 2019). Some of the research that have been done to investigate the LCA of edible insects as shown in Supplementary Table S1. One significant environmental concern related to insect cultured meat is the potential impact on biodiversity. Insects play a crucial role in pollination, nutrient cycling, and pest control in ecosystems worldwide. Large-scale insect farming for meat production could lead to the loss of diverse species and disrupt delicate ecological balances when insect is collected and genetically modified to fulfil the industrial needs (Zhang et al., 2022a). Although insect cultured meat has been promoted as a more environmentally friendly option, its production is not without drawbacks. Insect cultured meat production involves substantial energy consumption, including heating, lighting, and maintaining controlled environments for breeding (Barrett et al., 2023). This energy demand utilizes non-renewable resources, making the process unsustainable in the long run, especially if the industry scales up. However, from our comprehensive literature review, it can be concluded that no research was done to investigate the negative environmental impact of insect cultured meat; thus open an opportunity for the researchers to look into it.
4 Technological advancements and bio-robotics in insect cell culture
Technological advancements have revolutionized the field of bio-robotics in insect cell culture, enabling researchers to explore new frontiers in biotechnology and industrial applications. Moreover, the integration of advanced sensors and actuators in bio-robotic systems has enabled real-time monitoring and control of insect cell culture parameters (Reyes et al., 2020). Sensors can measure parameters such as dissolved oxygen, pH, temperature, glucose concentration, and cell viability, providing valuable data for process optimization and quality control (Reardon, 2021). Actuators can adjust these parameters in real-time, ensuring the optimal growth conditions for insect cells. Consequently, these advancements have not only increased productivity but also improved the product quality and consistency. In addition to process automation and monitoring, developments in data analytics and artificial intelligence have enhanced our understanding of insect cell culture (Altenburg et al., 2023). Machine learning algorithms can analyze vast amounts of data collected during the culture process, identifying patterns, correlations, and optimal culture conditions. These insights can be used to optimize culture parameters, media composition, and process design, thus increasing the overall efficiency and yield in insect cell culture.
Furthermore, the combination of bio-robotics and gene editing technologies such as Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas9 has unlocked new possibilities in insect cell engineering (Tyagi et al., 2020). Gene editing allows for the precise modification of insect cell lines, introducing desired genetic traits, improving protein expression, and enhancing productivity (Hillary and Ceasar,2022). Bio-robotics systems can automate the process of gene editing, making it faster, more accurate, and efficient. This advancement allows scientists to explore techniques for genetically modifying insect species to have higher protein content, improved amino acid profiles, and reduced allergenic compounds. The application of 3D and 4D bioprinting technologies has transformed the field of insect meat culture (Kumar et al., 2022). These techniques enable precise control and organization of cellular structures, enhancing the quality of insect meat products. 3D bioprinting involves layer-by-layer deposition of cellular structures to create three-dimensional objects. In the context of insect meat production, this technology allows scientists to create scaffolded cells, promoting tissue growth and development. It enables the generation of complex muscle fibers and organoids, mimicking the textures and tastes of traditional meat products (Balasubramanian et al., 2021). This technology serves as a bridge between traditional insect farming techniques and the creation of desirable food products. The 4D bioprinting takes 3D bioprinting a step further by integrating the dimension of time. This means that the printed structures have the ability to change shape or function over time, mimicking the natural growth and development process. Employing 4D bioprinting techniques can potentially streamline the process of insect meat production by allowing cells to self-organize and differentiate into specific tissues.
Among the innovative technologies employed in insect meat culture platforms, Magnetic-Based 3D (m3D) Cell Culture Technology stands out. This approach utilizes magnetic nanoparticles to enable the three-dimensional assembly of cells into functional tissues. By introducing magnetic forces, the cells can form compact structures, closely resembling native tissues (Tognato et al., 2019). m3D technology provides several advantages, including enhanced cell-cell communication and improved cell viability. It offers a sophisticated control system by manipulating the strength and direction of the magnetic field. This technique facilitates the organization of cells into intricate, meat-like structures, opening new possibilities for insect-based meat production. In order to sustain and ensure the long-term viability of lab-grown insects cell culture, proper preservation technology becomes crucial. This technology involves the techniques and methods used to maintain insect specimens in stable and viable conditions over extended periods. As an example, cryopreservation technique preserves the insectâs cell in suspended animation, essentially halting their biological processes until they are thawed for further use (Taylor et al., 2019). This technique helps prevent the aging and deterioration of organisms and facilitates their long-term storage. Another preservation technique for lab-grown insect cell culture is lyophilization. This technique involves removing the water content from the cells through sublimation, wherein the frozen water directly transforms from the solid state to the gaseous state without passing through the liquid phase (Merivaara et al., 2021). This process is achieved by subjecting the cells to a vacuum while gradually raising the temperature. The remaining freeze-dried cells can then be stored at room temperature for long periods, minimizing the need for freezers and reducing energy consumption.
Today, numerous companies are actively involved in this cultured meat industry, thus, revolutionizing the way we perceive insect cells and their applications (Table 3). As an example, UPSIDE Foods located in California was known for its innovative work in the field of cultured meat production. There are many other companies around the world such as Vowfood in Australia, Mewery in Czecz Republic that involve in this industry. Most of this companiesâ success was built on cutting-edge technology advancements in cellular agriculture and bioprocessing. In addition to that, they utilized sensory analysis and consumer feedback to continuously improve the taste, texture, and overall experience of their products. As an example, Steakholderfoods invented 3D printed meat that is produced via a 3D bio-printer. To create a cut of meat via 3D printing, animal cells are transformed into bio-ink and printed in accordance with a specific design scheme that layers all of the ingredients together to look and taste like a steak. Meanwhile, Israel-based food tech innovator Ever After Foods has launched its patented bioreactor platform for producing cultivated meat. This manufacturing plants require significantly lower capital and lower production costs, which it says results in a 700% increase in productivity when compared with other cultivated meat technology platforms (Table 4).
Comparison of traditional protein feed sources with generic protein feed
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Companies involved in cultured meat industry
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
5 Embryonic and adult myogenesis
Myogenesis, the process of muscle formation, is a fundamental biological process that has been extensively studied in numerous organisms, including insects. Furthermore, advances in muscle cell culture techniques have paved the way for improving insect cell culture systems, enabling the production of functional muscle tissue. Figure 4 shows the process involved in producing the lab grown insect meat. Myogenesis is a complex biological process essential for the development, growth, and function of muscle tissue in organisms (Mohammadabadi et al., 2021). Research on myogenesis offers tremendous potential for various fields, such as regenerative medicine, agriculture, and bioengineering. Understanding the mechanisms governing myogenesis in insects can lead to the development of strategies to manage insect pests, enhance agricultural productivity, and facilitate novel therapeutics for muscular disorders.
Process to produce lab grown insect meat.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Insect myogenesis research has progressed significantly in recent years, shedding light on the molecular and cellular events underlying muscle formation in insects. Studies utilizing model organisms, including Drosophila melanogaster and Tribolium castaneum, have revealed key regulatory genes and signaling pathways involved in insect muscle development (Zhang et al., 2022c). One significant discovery was the identification of Myogenic Regulatory Factors (MRFs), a family of transcription factors crucial for muscle specification and differentiation. In insects, this family includes myogenic factors known as MyoD, MyoR, and Twist, among others (Andrikou et al., 2013). These factors regulate the expression of various genes involved in muscle formation and play critical roles in determining muscle identity. Furthermore, recent studies have elucidated the role of signaling pathways in insect myogenesis, such as the Notch and Fibroblast Growth Factor (FGF) signaling pathways (Ludwig and Trimmer, 2021). These pathways influence the fate determination and differentiation of muscle cells by regulating cell-cell communication and downstream target genes.
Insect cell culture systems have become invaluable tools for understanding insect biology and facilitating various applications. The development of functional muscle tissue in insect cell cultures can provide novel avenues for exploring muscle-related disorders, insect physiology, and bioengineering of muscle tissues. Recent advancements in insect cell culture techniques have focused on promoting myogenesis and enhancing muscle maturation in vitro. One significant development is the incorporation of tissue engineering principles to optimize cell culture conditions for insect muscle formation. Mimicking the native muscle environment, researchers have engineered three-dimensional scaffolds using natural or synthetic biomaterials. These scaffolds provide structural support and facilitate the alignment and maturation of muscle cells, leading to improved functional tissue formation (Rao et al., 2023). In addition to scaffold-based approaches, the utilization of growth factors and inducers has proven effective in enhancing myogenesis in insect cell cultures (Chen et al., 2020). For instance, the addition of myogenic differentiation factors, such as transforming growth factor-beta (TGF-β) and insulin-like growth factor (IGF), has shown promising results in promoting muscle-specific gene expression and myotube formation. This allows the culture system to closely mimic the natural muscle development process.
The continual progress in myogenesis research and the development of insect cell culture techniques holds immense potential for various applications. Firstly, these advances can aid in understanding the mechanisms of muscle-related disorders in insects, leading to the identification of novel therapeutic targets. Additionally, insect cell-based muscle tissue engineering can contribute to regenerative medicine applications, including the development of muscle grafts and potential treatments for muscular dystrophies. Moreover, insect muscle tissue culture systems can serve as an efficient platform for testing drugs, toxicological studies, and high-throughput screening of potential therapeutic compounds. Insect-based muscle tissue engineering may also contribute to the production of cultured meat.
Insect cell culture allows researchers to study myogenesis in a controlled laboratory setting. However, several factors can affect myogenesis and the molecular pathways involved in insect cell culture. Understanding these factors is crucial for successful cultivation of insect cells and furthering our knowledge of muscle development. The choice of insect cell line plays a crucial role in myogenesis. Different insect species exhibit variations in their genetic makeup and cellular machinery, which can affect the efficiency and extent of myocyte differentiation. For example, Spodoptera frugiperda (Sf9) cells derived from fall armyworms are commonly used in insect cell culture due to their ease of cultivation and high transfection efficiency (Dhandapani et al., 2022). Other popular insect cell lines include Drosophila melanogaster (S2) cells and Trichoplusia ni (Hi5) cells (Stolt-Bergner et al., 2018). Researchers must carefully select the appropriate cell line for their specific study to ensure optimal myogenesis.
In addition to that, the culture conditions provided to the insect cells have a significant impact on myogenesis (Ludwig and Trimmer, 2021). The medium composition, nutrient availability, temperature, and pH can all affect the cellular environment and influence the differentiation process (Choi et al., 2021). For example, the addition of specific growth factors, such as transforming growth factor-beta (TGF-β) or fibroblast growth factor (FGF), can promote myogenesis in some insect cell lines (Shi and Chen, 2014). Additionally, optimizing the oxygen and carbon dioxide levels in the culture environment is crucial for adequate cell respiration and metabolism (Place et al., 2017). Researchers must carefully modulate these culture conditions to support myocyte development and ensure proper functioning of molecular pathways involved in muscle formation. Furthermore, the presence of extracellular matrix (ECM) components is vital for myogenesis in insect cell culture. The ECM provides structural support and signaling cues to the developing myocytes, guiding their differentiation and organization into functional muscle tissue (Jana et al., 2016). Various ECM proteins, including collagen, laminin, and fibronectin, are necessary for myocyte adhesion, migration, and alignment (Li et al., 2017). Additionally, integrin receptors on the surface of myocytes interact with these ECM proteins, transducing signaling events that regulate myogenesis (Grzelkowska-Kowalczyk, 2016). Proper ECM deposition in the culture environment is, therefore, crucial for the successful development of functional muscle tissue.
Moreover, molecular pathways play a central role in myogenesis in insect cell culture. The Notch, Wnt, and Hedgehog signaling pathways are some of the key pathways involved in muscle development. For example, the activation of Notch signaling inhibits myogenesis, while its inhibition promotes myocyte differentiation (Arya et al., 2013). The Wnt pathway promotes myogenesis by activating the transcription factor MyoD (Cirillo et al., 2017), which is essential for myocyte differentiation. The Hedgehog pathway mediates muscle patterning and differentiation by regulating the expression of important myogenic transcription factors such as Pax3 and Myf5 (HernaÌndez-HernaÌndez et al., 2017). Understanding and modulating these molecular pathways are critical for controlling myogenesis in insect cell culture.
6 Importance of insect body fat and muscle cells in culture
Muscle cells
In the field of insect tissue engineering for food production, one of the biggest challenges is developing a reliable and scalable supply of insect muscle and fat cells. Although insect cell culture has gained prominence in biotechnology, the establishment of stable muscle cell lines has received relatively less attention. Nevertheless, there have been reports on the isolation of primary and immortalized myoblasts from Drosophila melanogaster, as well as the generation of contractile myotubes from Manduca sexta (Baryshyan et al., 2012; DequeÌant et al., 2015). These approaches have been helpful in the research of insect muscle cells in the laboratory; however, in order to satisfy the needs of industrial-scale manufacturing, it is crucial to establish lines of muscle progenitor cells that are capable of prolonged differentiation and proliferation (Rubio et al., 2019b).
Although insect embryos are the most common source for isolating myogenic cells, they have also been successfully extracted from the dorsal vessel of larvae, pupal leg tissue, and ovarian tissue (Biofabrication with Insect Cells, n.d.). Primary muscle cell cultures are developed by disrupting tissues or embryos and transferring the cells to a culture tube with an insect growth media with either foetal bovine serum or the use of serum-free medium (DequeÌant et al., 2015). In most cases, the resulting cultured cell population is rather heterogeneous and contains several cell types. However, the purity of myogenic cells can be enhanced by coating the culture vessel with a substance like protamine and incorporating medium additives like insulin. Insulin is a common medium addition because it stimulates protein synthesis and the development of myotubes (muscle fibres) (Baryshyan et al., 2012). Ecdysone, an insect steroidal hormone, is utilised to induce cell differentiation, as researchers can trigger specific cellular changes, such as the development of distinct cell types or the initiation of specific cellular functions. Insect life cycles depend heavily on ecdysone, especially during development and metamorphosis (Xu et al., 2020).
A number of studies have documented the development of contractile cells in culture, with contractions persisting for weeks to months. For instance, embronic cell lines, T. exiguum IPLB-Tex2 and A. pernyi NISES-AnPe-426, achieved spontaneous immortalization, while the D. melanogaster R1-R7 cell line was genetically immortalized using the Rasv12 oncoprotein (Gunage et al., 2017). More studies must be conducted to investigate if the age-proliferation ability relationship in insects resembles that observed in mammalian cells, where cell multiplication declines as animals age. However, for mass production, it is essential to regulate the growth and maturation of cultured muscle cells. The steroid hormones known as ecdysteroids have been studied for their potential significance in skeletal muscle cells of insects. Research shows that 20-hydroxyecdysone stimulates myoblast proliferation at low concentrations, but that higher concentrations decrease development (Dinan et al., 2021). Methoprene, a juvenile hormone analogue, can prevent cell differentiation induced by high amounts of ecdysteroids. These hormonal controls allow for the production-stage regulation of muscle cell development (Parthasarathy and Palli, 2021).
Fat cells
Insect fat tissue is valuable in cellular agriculture due to its high content of essential fats and other nutrients. Culturing insect fat tissue alongside muscle is important to produce nutritious and flavorful food products (Sugii et al., 2023). Similar to adipose tissue in vertebrates, insect fat body tissue stores and metabolizes energy and nutrients (Li et al., 2022). In vitro culture of insect cells benefits from the presence of fat body cells, which support muscle cell survival and contraction and promote embryonic development. Fat body cells also synthesize important proteins such as vitellogenin, yolk protein precursors, and juvenile hormone binding protein (Hoshizaki, 2012).
Fat body tissue was among of the very initial insect tissues to be cultivated, and now more than 60 cell lines have been produced from different insect species. Fat cells are obtained from the abdomen of insect species which are then cultured using a wide range of culture media formulations including foetal bovine serum or serum free conditions (Li et al., 2022). Most approaches included culturing of the explants without digesting the tissue, however, one study reported digestion of the tissue explants using Dispase I (Rubio et al., 2019a). The cultures obtained from the culturing of fat cells typically consist of a mixture of different cell types. Initially, cells isolated from fat body tissue exhibit slow growth, but they can eventually be established as continuous cell lines (Letcher et al., 2022). Table 5 enlists different muscle as well as fat cell lines isolated from various insect species. Figure 5 shows the process of cell culture differentiation and proliferation of insect muscle and fat cells to obtain fat containing insect meat. However, it must be noted that the fat content and composition can affect the perceived richness, and overall flavor profile of the cultured insect meat. Different insect species have varying fat content and composition, which can influence taste characteristics. Muscle cells, on the other hand, play a role in the texture and tenderness of the meat. The presence of intramuscular fat can also be considered which can contribute to the overall taste perception (Starowicz et al., 2022).
Cells lines isolated from muscle and fat cells of different insect species
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Cell culture differentiation and proliferation of insect muscle and fat cells to obtain fat containing insect meat.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
7 Mutations in lab-grown cells
In lab-grown cells, mutations can occur due to a variety of reasons, including spontaneous errors during DNA replication, exposure to mutagenic agents, or unintentional genetic manipulations (Graham et al, 2020). Mutations within lab-grown cells can have both positive and negative consequences, influencing the outcomes of biomedical research (Figure 6). On one hand, mutations can hinder the reliability of experimental results. Unintended genetic alterations may introduce confounding variables or distort the intended objective of the research, rendering the findings less accurate or reproducible. However, on the other hand, mutations can lead to the development of novel traits or characteristics, offering opportunities for breakthrough discoveries. These mutations may provide insights into disease mechanisms, drug resistance, or cellular behavior, leading to advancements in medical treatments and therapies.
Effect of cell mutation in lab-grown insect cell to human and environment.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Nowadays, insect cell mutation for human health benefit is an emerging field of research with the potential to offer several advantages in medicine and biotechnology. As an example, insect cells, particularly from the baculovirus expression system, are used in the production of vaccines (Contreras-GoÌmez et al., 2014). Genetic mutations can enhance their ability to produce viral antigens or other vaccine components more efficiently. Improved vaccine production can lead to increased vaccine availability, lower production costs, and faster response to emerging infectious diseases, ultimately benefiting human health. In addition to that, insect cells are employed for the production of recombinant proteins and biopharmaceuticals. Genetic modifications can enhance their protein expression capabilities, resulting in higher yields of therapeutic proteins (OâFlaherty et al., 2020). This is particularly valuable for producing essential therapeutic proteins such as insulin, monoclonal antibodies, and enzyme replacement therapies, which are crucial for managing various medical conditions.
Mutated insect cells can also serve as valuable tools in drug discovery and development. They can be engineered to express specific human genes involved in diseases or drug metabolism (Brouwer et al., 2013), allowing researchers to study drug candidates more effectively. These cells can be used for high-throughput screening assays, toxicity testing, and target validation, expediting the drug development process. In term of disease research and modelling, insect cells can be genetically modified to carry human disease-associated mutations (Huynh and Vale, 2017). This enables researchers to create cellular models for studying genetic disorders, neurodegenerative diseases, and various other conditions. Such models can provide insights into disease mechanisms, potential therapeutic targets, and drug testing platforms. Genetic modification of insects like mosquitoes can be explored to reduce their ability to transmit diseases such as malaria, dengue, or Zika virus (Wang et al., 2021). This approach has the potential to mitigate the spread of these diseases and protect human populations.
Mutations also have significant implications on the insect microbiome and gene expression for lab-grown insect meat production (Table 6). These mutations can affect the composition and function of the microbiome, which in turn can impact the nutritional quality, growth rates, and overall health of the lab-grown insects. Mutations affecting the expression of antimicrobial peptides (AMPs) in the insect gut can alter the balance between beneficial and harmful bacteria. As an example, Zhao et al. (2023) reported that deletion of uracil in Bt GS57 strain changed the diversity of gut bacteria in Spodoptera exigua. In addition to that, mutations that affect the expression of genes involved in nutrient acquisition and utilization can impact the nutritional content of lab-grown insect meat (Siddiqui et al., 2022). Thus, researchers can take this opportunity in targeting specific genetic mutations to improve nutrient synthesis by the microbiota. However, extensive research and safety assessments are necessary to ensure that such genetic modifications are safe for both the insects and the consumers of lab-grown insect meat. While the ability to culture and manipulate cells in the laboratory has revolutionized scientific research and medical treatments, it also brings with it the potential for unintended consequences when cells undergo genetic mutations that lead to uncontrolled growth (Cretu et al., 2016). One of the most alarming possibilities is the development of cancerous growths. These mutations may arise spontaneously or be induced by the in vitro culturing process itself, exposure to mutagenic agents, or genetic engineering techniques. As a consequence, it may lead to contamination of cell cultures. This can compromise the integrity of experiments and, in a worst-case scenario, spread mutated cells to other cultures or research settings.
Role of insect microbiome in gene expression profile and health benefit
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001131
Drawbacks of insect labriculture
Although insect labriculture has gained significant attention as a sustainable and alternative source of protein, it comes with its own set of food safety risks. One of the key concerns regarding insects as a food source is the potential for microbial contamination. Bacteria, such as Salmonella and Escherichia coli, can pose a significant health risk if ingested (Pava-Ripoll et al., 2015). Similarly, mycotoxins produced by fungi can contaminate insect feed and subsequently be transferred to humans (Evans and Shao, 2022). Parasites, such as mites and worms, may also be present in insect farming environments and can cause infection if consumed (GaÅęcki and SokoÌÅ, 2019). Furthermore, allergenic proteins, commonly found in insects, may trigger allergic reactions in susceptible individuals (Barre et al., 2018). In addition to microbial hazards, heavy metals like lead, mercury, and cadmium may accumulate in insects if reared in contaminated environments (Meyer et al., 2021). These metals possess significant health risks and can lead to long-term health implications upon consumption. Moreover, anti-nutrients, naturally occurring compounds that interfere with nutrient absorption, may be present in insects. High levels of anti-nutrients can impact the nutritional value of insects and compromise the quality of the final products (Kumar et al., 2022).
Insect labriculture, although often touted as environmentally friendly, can contribute to CO2 evolution. The temperature and humidity required for insect rearing can increase energy demands and subsequent CO2 emissions. Additionally, the production and transportation of feedstock, such as grain and vegetables, may result in greenhouse gas emissions (Cadinu et al., 2020). These factors should be carefully considered to minimize the environmental impact of insect labriculture. In contrast to traditional livestock farming, insect labriculture is often viewed as a more sustainable option due to its lower land, water, and feed requirements. Insects have a higher feed conversion efficiency (Van Broekhoven et al., 2015), meaning they require less feed to produce a comparable amount of protein than conventional livestock (Supplementary Table S4). Furthermore, insect farming can utilize organic waste products, effectively reducing waste and promoting circular economies. These sustainability aspects make insect labriculture an attractive solution to current food security issues. While insect labriculture presents numerous benefits, cost-effectiveness remains a crucial consideration. Currently, the high cost of insect farming equipment and infrastructure can hinder widespread adoption. Moreover, the production of insect feed can be expensive, especially if specific requirements for optimal growth are necessary. However, as the industry develops and scales up, the associated costs are expected to decrease, making it more affordable and accessible to consumers. Considering all of this drawback, it is essential to recognize and address the food safety risks it presents to ensure the health and well-being of consumers. Proper monitoring and implementation of safety measures are required to mitigate microbial contamination, heavy metal accumulation, and other potential hazards. Simultaneously, efforts should be made to minimize the environmental impact and promote cost-effectiveness of insect labriculture. With careful consideration and regulatory oversight, insect farming can offer a viable solution to protein production while maintaining food safety standards.
8 Conclusion
This review paper provides a comprehensive overview of the burgeoning field of lab-grown insect meat production. This review delves deep into the various facets of this innovative and sustainable approach to addressing the global challenges of food security, environmental sustainability, and ethical concerns associated with traditional meat production. One of the key takeaways from this review is the immense potential of lab-grown insect meat to revolutionize the food industry. Insects are a highly efficient source of protein, requiring significantly less land, water, and feed compared to traditional livestock. Moreover, the paper highlights the nutritional benefits of insect-based proteins, which are rich in essential amino acids, vitamins, and minerals. Lab-grown insect meat has the potential to become a crucial component of a balanced and sustainable diet for a growing global population. From a chemical perspective, the paper provides valuable insights into the composition of lab-grown insect meat, detailing the proteins, lipids, and other macronutrients that make up this alternative food source. It also discusses the methods and techniques used in the cultivation of insect cells, shedding light on the challenges and opportunities in optimizing their growth and nutritional content.
Furthermore, the biological aspects explored in this review offer a comprehensive understanding of the environmental benefits of lab-grown insect meat. By reducing the carbon footprint, conserving resources, and minimizing greenhouse gas emissions associated with traditional livestock farming, this innovative approach aligns with the urgent need for sustainable food production to combat climate change. Moreover, lab-grown insect meat still has challenge for examine and explores the various detrimental factors associated with insect cultured meat and argues that caution should be exercised before widely adopting this technology, Large-scale insect farming for meat production could lead to the loss of diverse species and disrupt delicate ecological balances, and Insect cultured meat production involves substantial energy consumption (heating, lighting, and maintaining controlled environments for breeding). In addition, the findings of this review will contribute to a better understanding of lab-grown insect meat, offering insights into its potential as a sustainable alternative to conventional meat production. As lab-grown insect meat continues to attract attention from researchers, environmentalists, and policymakers, this comprehensive review is a timely and critical resource to inform future developments and discussions in the field. Ethical considerations surrounding lab-grown insect meat are also addressed, emphasizing the reduction of animal suffering and the potential to alleviate the ethical dilemmas faced by those who choose to follow vegetarian or vegan diets. This research paper underscores the importance of lab-grown insect meat as a promising and sustainable solution to the challenges of the 21st century food system. As the world faces increasing pressure to feed a growing population while minimizing environmental impact, this review serves as a valuable resource for scientists, policymakers, and industry stakeholders working to harness the potential of lab-grown insect meat as a transformative force in the future of food. With continued research and development, lab-grown insect meat may well emerge as a cornerstone of a more sustainable and ethical global food supply.
Corresponding authors; e-mail:Â s.siddiqui@dil-ev.de; ibrah001@ncat.edu
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.25303876
Authors contribution statement
S.A.S.: conceptualization, methodology, writing â original draft, writing â review and editing, validation, formal analysis, resources, visualization, data curation, project administration, investigation, supervision. N.N.: writing â original draft. Y.S.W.: formal analysis. T.K.: writing â original draft, conceptualization. B.Y.: review and editing. S.A.I.: validation, funding.
Conflict of interest
The authors declare no conflict of interest.
Funding
This publication was made possible by grant numbers NC.X-267-5-12-170-1 and NC.X 359-5-24-170-1 from the National Institute of Food and Agriculture (NIFA) and the Department of Family and Consumer Sciences and the Agriculture Research Station at North Carolina Agriculture and Technical State University (Greensboro, NC, USA 27411). This work was also supported, in part, by 1890 Capacity Building Program grant no. (2020-38821-31113/project accession no. 021765).
References
Adedibu, P.A., 2023. Ecological problems of agriculture: impacts and sustainable solutions. ScienceOpen preprints.
Agathos, S.N., 2014. Insect cell culture. In: Manual of industrial microbiology and biotechnology, pp. 212-222. https://doi.org/10.1128/9781555816827.CH15
Ahmad, K., Lim, J.H., Lee, E.J., Chun, H.J., Ali, S., Ahmad, S.S., Shaikh, S. and Choi, I., 2021. Extracellular matrix and the production of cultured meat. Foods 10: 3116. https://doi.org/10.3390/foods10123116
Akhtar, Y. and Isman, M.B., 2018. Insects as an alternative protein source. In: Yada, R.Y. (ed.) Proteins in food processing. Woodhead Publishing, pp. 263-288. https://doi.org/10.1016/B978-0-08-100722-8.00011-5
Akiyama, Y., Sakuma, T., Funakoshi, K., Hoshino, T., Iwabuchi, K. and Morishima, K., 2013. Atmospheric-operable bioactuator powered by insect muscle packaged with medium. Lab on a Chip 13: 4870-4880. https://doi.org/10.1039/C3LC50490E
Alam, M.K., Koomson, E., Zou, H., Yi, C., Li, C.W., Xu, T. and Yang, M., 2018. Recent advances in microfluidic technology for manipulation and analysis of biological cells (2007-2017). Analytica Chimica Acta 1044: 29-65.
Alexander, P., 2022. Eat insects to save the planet? This academic feels it could help in achieving a sustainable food system. Available at: https://economictimes.indiatimes.com/magazines/panache/eat-insects-to-save-the-planet-this-academic-feels-it-could-help-in-achieving-a-sustainable-food-system/articleshow/94181392.cms?utm_source=contentofinterest&utm_medium=text&utm_campaign=cppst
Alexander, P., Brown, C., Arneth, A., Dias, C., Finnigan, J., Moran, D. and Rounsevell, M.D., 2017. Could consumption of insects, cultured meat or imitation meat reduce global agricultural land use? Global Food Security 15: 22-32.
Altenburg, J.J., Klaverdijk, M., Cabosart, D., Desmecht, L., Brunekreeft-Terlouw, S.S., Both, J. and Martens, D.E., 2023. Real-time online monitoring of insect cell proliferation and baculovirus infection using digital differential holographic microscopy and machine learning. Biotechnology Progress 39: 3318.
An, Y., Rininger, J.A., Jarvis, D.L., Jing, X., Ye, Z., Aumiller, J.J., Eichelberger, M. and Cipollo, J.F., 2013. Comparative glycomics analysis of influenza Hemagglutinin (H5N1) produced in vaccine relevant cell platforms. Journal of Proteome Research 12: 3707-3720. https://doi.org/10.1021/PR400329K
Arumugam, M.K., Paal, M.C., Donohue, T.M., Ganesan, M., Osna, N.A. and Kharbanda, K.K., 2021. Beneficial effects of betaine: a comprehensive review. Biology 10: 450-456. https://doi.org/10.3390/BIOLOGY10060456
Arya, M.A., Tai, A.K., Wooten, E.C., Parkin, C.D., Kudryavtseva, E. and Huggins, G.S., 2013. Notch pathway activation contributes to inhibition of C2C12 myoblast differentiation by ethanol. PLOS ONE 8: 71632.
Barre, A., Simplicien, M., Cassan, G., Benoist, H. and RougeÌ, P., 2018. Food allergen families common to different arthropods (mites, insects, crustaceans), mollusks and nematods: cross-reactivity and potential cross-allergenicity. Revue Francaise dâallergologie 58: 581-593.
Barrett, M., Chia, S.Y., Fischer, B. and Tomberlin, J.K., 2023. Welfare considerations for farming black soldier flies, Hermetia illucens (Diptera: Stratiomyidae): a model for the insects as food and feed industry. Journal of Insects as Food and Feed 9: 119-148.
Baryshyan, A.L., Domigan, L.J., Hunt, B., Trimmer, B.A. and Kaplan, D.L., 2014. Self-assembled insect muscle bioactuators with long term function under a range of environmental conditions. RSC Advances 4: 39962. https://doi.org/10.1039/C4RA08438A
Baryshyan, A.L., Woods, W., Trimmer, B.A. and Kaplan, D.L., 2012. Isolation and maintenance-free culture of contractile myotubes from Manduca sexta embryos. PLOS ONE 7: 31598. https://doi.org/10.1371/journal.pone.0031598
Bhat, Z.F., Kumar, S. and Bhat, H.F., 2017. In vitro meat: a future animal-free harvest. Critical Reviews in Food Science and Nutrition 57: 782-789.
Biofabrication with insect cells. (n.d.). Available at: https://zenodo.org/records/7245722
Boegel, S.J., 2020. Towards a chemically defined medium for Sf-9 cell culture: micronutrients reduce dependence on yeast extract. UWSpace. Available at: http://hdl.handle.net/10012/15430
Brackney, D.E., Scott, J.C., Sagawa, F., Woodward, J.E., Miller, N.A., Schilkey, F.D., Mudge, J., Wilusz, J., Olson, K.E., Blair, C.D. and Ebel, G.D., 2010. C6/36 Aedes albopictus cells have a dysfunctional antiviral RNA interference response. PLOS Neglected Tropical Diseases 4: 856. https://doi.org/10.1371/journal.pntd.0000856
Broderick, N.A., Buchon, N. and Lemaitre, B., 2014. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. MBio 5: 10-1128.
Brouwer, K.L., Keppler, D., Hoffmaster, K.A., Bow, D.A., Cheng, Y., Lai, Y. and International Transporter Consortium, 2013. In vitro methods to support transporter evaluation in drug discovery and development. Clinical Pharmacology & Therapeutics 94: 95-112.
Bustamante, M.M., Nobre, C.A., Smeraldi, R., Aguiar, A.P., Barioni, L.G., Ferreira, L.G. and Ometto, J.P., 2012. Estimating greenhouse gas emissions from cattle raising in Brazil. Climatic Change 115: 559-577.
Butler, M., 2015. Serum and protein free media. In: Al-Rubeai, M. (ed.) Animal cell culture. Springer International Publishing, Cham, Switzerland, pp. 223-236. https://doi.org/10.1007/978-3-319-10320-4_8
Campuzano, S. and Pelling, A.E., 2019. Scaffolds for 3D cell culture and cellular agriculture applications derived from non-animal sources. Frontiers in Sustainable Food Systems 3: 30-38. https://doi.org/10.3389/FSUFS.2019.00038/BIBTEX
Cannon, K.M. and Britt, D.T., 2019. Feeding one million people on Mars. New Space 7: 245-254.
Chakravorty, J., Ghosh, S. and Meyer-Rochow, V.B., 2013. Comparative survey of entomophagy and entomotherapeutic practices in six tribes of Eastern Arunachal Pradesh (India). Journal of Ethnobiology and Ethnomedicine 9: 1-12.
Chan, L.C.L. and Reid, S., 2016. Development of serum-free media for lepidopteran insect cell lines. Methods in Molecular Biology 1350: 161-196. https://doi.org/10.1007/978-1-4939-3043-2_8
Chang, L., Li, Z., Guo, H., Zhang, W., Lan, W., Wang, J., Shen, G., Xia, Q. and Zhao, P., 2021. Function of polyamines in regulating cell cycle progression of cultured silkworm cells. Insects 12: 6614-6624. https://doi.org/10.3390/insects12070624
Chen, X.Y., Chen, J.Y., Tong, X.M., Mei, J.G., Chen, Y.F. and Mou, X.Z., 2020. Recent advances in the use of microcarriers for cell cultures and their ex vivo and in vivo applications. Biotechnology Letters 42: 1-10.
Choi, K.H., Yoon, J.W., Kim, M., Lee, H.J., Jeong, J., Ryu, M. and Lee, C.K., 2021. Muscle stem cell isolation and in vitro culture for meat production: a methodological review. Comprehensive Reviews in Food Science and Food Safety 20: 429-457.
Cirillo, F., Resmini, G., Ghiroldi, A., Piccoli, M., Bergante, S., Tettamanti, G. and Anastasia, L., 2017. Activation of the hypoxia-inducible factor 1a promotes myogenesis through the noncanonical Wnt pathway, leading to hypertrophic myotubes. The FASEB Journal 31: 2146-2156.
Contreras-GoÌmez, A., SaÌnchez-MiroÌn, A., GarcıÌa-Camacho, F., Molina-Grima, E. and Chisti, Y., 2014. Protein production using the baculovirus-insect cell expression system. Biotechnology Progress 30: 1-18.
Cowan, N., Ferrier, L., Spears, B., Drewer, J., Reay, D. and Skiba, U., 2022. CEA systems: the means to achieve future food security and environmental sustainability? Frontiers in Sustainable Food Systems 6: 891256.
Cretu, C., SchmitzovaÌ, J., Ponce-Salvatierra, A., Dybkov, O., De Laurentiis, E.I., Sharma, K. and Pena, V., 2016. Molecular architecture of SF3b and structural consequences of its cancer-related mutations. Molecular Cell 64: 307-319.
Dambuza, I.M., He, C., Choi, J.K., Yu, C.R., Wang, R., Mattapallil, M.J. and Egwuagu, C.E., 2017. IL-12p35 induces expansion of IL-10 and IL-35-expressing regulatory B cells and ameliorates autoimmune disease. Nature Communications 8: 709-719.
de Carvalho, N.M., Walton, G.E., Poveda, C.G., Silva, S.N., Amorim, M., Madureira, A.R. and Jauregi, P., 2019. Study of in vitro digestion of Tenebrio molitor flour for evaluation of its impact on the human gut microbiota. Journal of Functional Foods 59: 101-109.
DequeÌant, M.L., Fagegaltier, D., Hu, Y., Spirohn, K., Simcox, A., Hannon, G.J. and Perrimon, N., 2015. Discovery of progenitor cell signatures by time-series synexpression analysis during Drosophila embryonic cell immortalization. Proceedings of the National Academy of Sciences of the United States of America 112: 12974-12979. https://doi.org/10.1073/pnas.151772911
Dhandapani, R.K., Gurusamy, D. and Palli, S.R., 2022. Protamine-lipid-dsRNA nanoparticles improve RNAi efficiency in the fall armyworm, Spodoptera frugiperda. Journal of Agricultural and Food Chemistry 70: 6634-6643.
Dimou, A., Tsimihodimos, V. and Bairaktari, E., 2022. The critical role of the branched chain amino acids (BCAAs) catabolism-regulating enzymes, branched-chain aminotransferase (BCAT) and branched-chain α-keto acid dehydrogenase (BCKD), in human pathophysiology. International Journal of Molecular Sciences 23: 4022.
Dinan, L., Dioh, W., Veillet, S. and Lafont, R., 2021. 20-Hydroxyecdysone, from plant extracts to clinical use: therapeutic potential for the treatment of neuromuscular, cardio-metabolic and respiratory diseases. Biomedicines 9: 492. https://doi.org/10.3390/BIOMEDICINES9050492
Doke, S.K. and Dhawale, S.C., 2015. Alternatives to animal testing: a review. Saudi Pharmaceutical Journal 23: 223-229.
Dong, Z.Q., Chen, T.T., Zhang, J., Hu, N., Cao, M.Y., Dong, F.F. and Pan, M.H., 2016. Establishment of a highly efficient virus-inducible CRISPR/Cas9 system in insect cells. Antiviral Research 130: 50-57.
Drosophila cultured cell lines | DRSC/TRiP functional genomics resources & DRSC-BTRR. (n.d.). Available at: https://fgr.hms.harvard.edu/drosophila-cultured-cell-lines
Dumont, J., Euwart, D., Mei, B., Estes, S. and Kshirsagar, R., 2016. Human cell lines for biopharmaceutical manufacturing: history, status, and future perspectives. Critical Reviews in Biotechnology 36: 1110-1122.
Eriksson, U., Blomqvist, J., Lüllau, E. and Häggström, L., 2005. Metalloproteinase activity is the sole factor responsible for the growth-promoting effect of conditioned medium in Trichoplusia ni insect cell cultures. Journal of Biotechnology 119: 76-86. https://doi.org/10.1016/j.jbiotec.2005.05.020
ESF 921 Insect Cell Culture Medium, protein free â expression systems. (n.d.). Available at: https://expressionsystems.com/product/esf-921-insect-cell-culture-medium-protein-free/
Ettinger, A. and Wittmann, T., 2014. Fluorescence live cell imaging. Methods in Cell Biology 123: 77-94.
Evans, N.M. and Shao, S., 2022. Mycotoxin metabolism by edible insects. Toxins 14: 210-217.
ExpiSfTM CD Medium. (n.d.). Available at: https://doi.org/10.3390/toxins14030217
Express FiveTM SFM. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/10486025
GaÅęcki, R. and SokoÌÅ, R., 2019. A parasitological evaluation of edible insects and their role in the transmission of parasitic diseases to humans and animals. PLOS ONE 14: 219303.
Ghosh, A., Dhall, H., Dietzgen, R.G. and Jain, R.K., 2020. Insect cell culture as a tool in plant virus research: a historical overview. Phytoparasitica 48: 287-303. https://doi.org/10.1007/S12600-020-00795-7/METRICS
Ghosh, S., Lee, S.M., Jung, C. and Meyer-Rochow, V.B., 2017. Nutritional composition of five commercial edible insects in South Korea. Journal of Asia-Pacific Entomology 20: 686-694.
Gomes, A.R., Byregowda, S.M., Veeregowda, B.M. and Balamurugan, V., 2016. An overview of heterologous expression host systems for the production of recombinant proteins. Advances in Animal and Veterinary Sciences 4: 346-356. http://dx.doi.org/10.14737/journal.aavs/2016/4.7.346.356
Gotoh, T., Chiba, K. and Kikuchi, K.I., 2004. Oxygen consumption profiles of Sf-9 insect cells and their culture at low temperature to circumvent oxygen starvation. Biochemical Engineering Journal 17: 71-78. https://doi.org/10.1016/S1369-703X(03)00140-2
Grace, T.D.C., 1962. Establishment of four strains of cells from insect tissues grown in vitro. Nature 195: 4843. https://doi.org/10.1038/195788a0
Graceâs Insect Medium, supplemented. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/11605094
Graceâs Insect Medium, unsupplemented. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/11595030
Graham, N., Patil, G.B., Bubeck, D.M., Dobert, R.C., Glenn, K.C., Gutsche, A.T. and Morrell, P.L., 2020. Plant genome editing and the relevance of off-target changes. Plant Physiology 183: 1453-1471.
Grzelkowska-Kowalczyk, K., 2016. The importance of extracellular matrix in skeletal muscle development and function. In: Travascio, F. (ed.) Composition and function of the extracellular matrix in the human body. IntechOpen, Croatia, pp. 3-24.
Gunage, R.D., Dhanyasi, N., Reichert, H. and Raghavan, K.V., 2017. Drosophila adult muscle development and regeneration. Seminars in Cell and Developmental Biology 72: 56-66. https://doi.org/10.1016/j.semcdb.2017.11.017
Hadi, J. and Brightwell, G., 2021. Safety of alternative proteins: technological, environmental and regulatory aspects of cultured meat, plant-based meat, insect protein and single-cell protein. Foods 10: 1226.
Hawkey, K.J., Lopez-Viso, C., Brameld, J.M., Parr, T. and Salter, A.M., 2021. Insects: a potential source of protein and other nutrients for feed and food. Annual Review of Animal Biosciences 9: 333-354.
HernaÌndez-HernaÌndez, J.M., GarcıÌa-GonzaÌlez, E.G., Brun, C.E. and Rudnicki, M.A., 2017. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. In: Seminars in cell & developmental biology. Academic Press, Cambridge, MA, USA.
Hillary, V.E. and Ceasar, S.A., 2022. Prime editing in plants and mammalian cells: mechanism, achievements, limitations, and future prospects. BioEssays 44: 2200032.
Hoshizaki, D.K., 2012. Fat body. The Insects 5: 132-146. https://doi.org/10.1017/CBO9781139035460.009
Huynh, W. and Vale, R.D., 2017. Disease-associated mutations in human BICD2 hyperactivate motility of dynein-dynactin. Journal of Cell Biology 216: 3051-3060.
Ianovici, I., Zagury, Y., Redenski, I., Lavon, N. and Levenberg, S., 2022. 3D-printable plant protein-enriched scaffolds for cultivated meat development. Biomaterials 284: 121487. https://doi.org/10.1016/j.biomaterials.2022.121487
Imseng, N., Steiger, N., Frasson, D., Sievers, M., Tappe, A., Greller, G., Eibl, D. and Eibl, R., 2014. Single-use wave-mixed versus stirred bioreactors for insect-cell/BEVS-based protein expression at benchtop scale. Engineering in Life Sciences 14: 264-271. https://doi.org/10.1002/ELSC.201300131
Insect cell culture media. (n.d.). Available at: https://www.thermofisher.com/de/en/home/life-science/cell-culture/insect-cell-culture.html
Insect media â Gibco insect cell culture media | Thermo Fisher Scientific â IN. (n.d.). Available at: https://www.thermofisher.com/de/en/home/life-science/cell-culture/insect-cell-culture/insect-cell-culture-misc/serum-free-media.html
IPL-41 Insect Medium (1X). (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/11405081
Jana, S., Cooper, A. and Zhang, M., 2013. Chitosan scaffolds with unidirectional microtubular pores for large skeletal myotube generation. Advanced Healthcare Materials 2: 557-561. https://doi.org/10.1002/ADHM.201200177
Jana, S., Levengood, S.K.L. and Zhang, M., 2016. Anisotropic materials for skeletal-muscle-tissue engineering. Advanced Materials 28: 10588-10612.
Jensen, K., Kristensen, T.N., Heckmann, L.H. and Sørensen, J.G., 2017. Breeding and maintaining high-quality insects. Insects as food and feed: from production to consumption. Wageningen Academic Publishers, Wageningen, the Netherlands, pp. 175-198.
Jiang, Y., He, Z., Zhao, M., Wang, C., Sun, L. and Feng, Y., 2017. Oil content and fatty acid composition of six kinds of common edible dragonfly naiads. China Oils and Fats 42: 135-139.
Kaiser, S.C., Decaria, P.N., Seidel, S. and Eibl, D., 2022. Scaling-up of an insect cell-based virus production process in a novel single-use bioreactor with flexible agitation. Chemie Ingenieur Technik 94: 1950-1961. https://doi.org/10.1002/CITE.202200103
Ker, D.F.E., Weiss, L.E., Junkers, S.N., Chen, M., Yin, Z., Sandbothe, M.F. and Campbell, P.G., 2011. An engineered approach to stem cell culture: automating the decision process for real-time adaptive subculture of stem cells. PLOS ONE 6: 27672.
Kim, T.K., Yong, H.I., Kim, Y.B., Kim, H.W. and Chi, Y.S., 2019. Edible insects as a protein source: a review of public perception, processing technology, and research trends. Food Science of Animal Resources 39: 521-540. https://doi.org/10.5851/kosfa.2019.e53
KourÌimskaÌ, L. and AdaÌmkovaÌ, A., 2016. Nutritional and sensory quality of edible insects. NFS Journal 4: 22-26.
Kraham, S.J., 2017. Environmental impacts of industrial livestock production. In: Steier, G. and Patel, K. (eds.) International farm animal, wildlife and food safety law. Springer, Cham, Switzerland. https://doi.org/10.1007/978-3-319-18002-1_1
Kumar, P., Mehta, N., Abubakar, A.A., Verma, A.K., Kaka, U., Sharma, N. and Lorenzo, J.M., 2022. Potential alternatives of animal proteins for sustainability in the food sector. Food Reviews International 39: 5703-5728. https://doi.org/10.1080/87559129.2022.2094403
Lee, C.W., Kc, M., Ngunjiri, J.M., Ghorbani, A. and Lee, K., 2023. TLR3 and MDA5 knockout DF-1 cells enhance replication of avian orthoavulavirus 1. Avian Diseases 67: 94-101.
Lee, K.P., Simpson, S.J. and Wilson, K., 2008. Dietary protein-quality influences melanization and immune function in an insect. Functional Ecology 22: 1052-1061.
Letcher, S.M., Rubio, N.R., Ashizawa, R.N., Saad, M.K., Rittenberg, M.L., Mccreary, A., Ali, A., Calkins, O.P., Trimmer, B.A. and Kaplan, D.L., 2022. In vitro insect fat cultivation for cellular agriculture applications. ACS Biomaterials Science and Engineering 8: 3785-3796. https://doi.org/10.1021/acsbiomaterials.2c00093
Li, C.H., Yang, I.H., Ke, C.J., Chi, C.Y., Matahum, J., Kuan, C.Y., Celikkin, N., Swieszkowski, W. and Lin, F.H., 2022. The production of fat-containing cultured meat by stacking aligned muscle layers and adipose layers formed from gelatin-soymilk scaffold. Frontiers in Bioengineering and Biotechnology 10: 540-550. https://doi.org/10.3389/FBIOE.2022.875069/BIBTEX
Li, Y., Xiao, Y. and Liu, C., 2017. The horizon of materiobiology: a perspective on material-guided cell behaviors and tissue engineering. Chemical Reviews 117: 4376-4421.
Liu, W., Hao, Z., Florkowski, W.J., Wu, L. and Yang, Z., 2022. A review of the challenges facing global commercialization of the artificial meat industry. Foods 11: 3609.
Ludwig, J.C. and Trimmer, B.A., 2021. Metamorphosis in insect muscle: insights for engineering muscle-based actuators. Tissue Engineering Part B: Reviews 27: 330-340.
Lynn, D.E., 2002. Methods for maintaining insect cell cultures. Journal of Insect Science 2: 1-9. https://doi.org/10.1093/JIS/2.1.9
Ma, D., Wang, Y. and Dai, W., 2018. Silk fibroin-based biomaterials for musculoskeletal tissue engineering. Materials Science & Engineering: C 9: 456-469. https://doi.org/10.1016/J.MSEC.2018.04.062
Ma, F., Xu, S., Tang, Z., Li, Z. and Zhang, L., 2021. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosafety and Health 3: 32-38.
Ma, H., Nandakumar, S., Bae, E.H., Chin, P.J. and Khan, A.S., 2019. The Spodoptera frugiperda Sf9 cell line is a heterogeneous population of rhabdovirus-infected and virus-negative cells: isolation and characterization of cell clones containing rhabdovirus X-gene variants and virus-negative cell clones. Virology 536: 125-133. https://doi.org/10.1016/J.VIROL.2019.08.001
Mendoza, T.C., 2023. Transforming meat based to plant-based diet is addressing food security and climate crisis in this millenium: a review. International Journal of Agricultural Technology 19: 517-540.
Merivaara, A., Zini, J., Koivunotko, E., Valkonen, S., Korhonen, O., Fernandes, F.M. and Yliperttula, M., 2021. Preservation of biomaterials and cells by freeze-drying: change of paradigm. Journal of Controlled Release 336: 480-498.
Meyer-Rochow, V.B., Gahukar, R.T., Ghosh, S. and Jung, C., 2021. Chemical composition, nutrient quality and acceptability of edible insects are affected by species, developmental stage, gender, diet, and processing method. Foods 10: 1036.
Miglietta, P.P., De Leo, F., Ruberti, M. and Massari, S., 2015. Mealworms for food: a water footprint perspective. Water 7: 6190-6203.
Miller, J.L., 2013. Iron deficiency anemia: a common and curable disease. Cold Spring Harbor Perspectives in Medicine 3: 011866.
Miller, R.K., 2020. A 2020 synopsis of the cell-cultured animal industry. Animal Frontiers 10: 64-72. https://doi.org/10.1093/af/vfaa031
MilliporeSigmaTM NovagenTM BacVectorTM Insect Cell Medium. (n.d.). Available at: https://www.fishersci.com/shop/products/emd-millipore-novagen-bacvector-insect-cell-medium/705903
Mitsuhashi, J., 1998. Polyamine as a growth promoter for cultured insect cells. In vitro cellular and developmental biology. Animal 34: 619-621. https://doi.org/10.1007/S11626-996-0007-9/METRICS
Mnisi, C.M., Oyeagu, C.E. and Ruzvidzo, O., 2022. Mopane worm (Gonimbrasia belina Westwood) meal as a potential protein source for sustainable quail production: a review. Sustainability 14: 5511. https://doi.org/10.3390/su14095511
Mohamed, E.H., 2015. Fatty acids contents of the edible migratory locust Locusta migratoria, Linnaeus, 1758 (Orthoptera: Acrididae). International Journal Advance Pharmautical Biology Chemistry 4: 746-750.
Mohammadabadi, M., Bordbar, F., Jensen, J., Du, M. and Guo, W., 2021. Key genes regulating skeletal muscle development and growth in farm animals. Animals 11: 835.
Moraes, AÌ.M., Jorge, S.A.C., Astray, R.M., Suazo, C.A.T., CalderoÌn Riquelme, C.E., Augusto, E.F.P., Tonso, A., Pamboukian, M.M., Piccoli, R.A.M., Barral, M.F. and Pereira, C.A., 2012. Drosophila melanogaster S2 cells for expression of heterologous genes: from gene cloning to bioprocess development. Biotechnology Advances 30: 613-628. https://doi.org/10.1016/J.BIOTECHADV.2011.10.009
Nagaraj, K., Sarfstein, R., Laron, Z. and Werner, H., 2022. Long-term IGF1 stimulation leads to cellular senescence via functional interaction with the thioredoxin-interacting protein, TXNIP. Cells 11: 3260. https://doi.org/10.3390/CELLS11203260/S1
Narzari, S. and Sarmah, J., 2017. Nutritional aspects of an aquatic edible insect Sympetrum sp. (Odonata: Libellulidae) of Assam, northeast India. International Journal of Food Sciences and Nutrition 2: 38-42.
Nguyen, Q., Qi, Y.M., Wu, Y., Chan, L.C., Nielsen, L.K. and Reid, S., 2011. In vitro production of Helicoverpa baculovirus biopesticides â automated selection of insect cell clones for manufacturing and systems biology studies. Journal of Virological Methods 175: 197-205.
Nijdam, D., Rood, T. and Westhoek, H., 2012. The price of protein: review of land use and carbon footprints from life cycle assessments of animal food products and their substitutes. Food Policy 37: 760-770.
Nowakowski, A.C., Miller, A.C., Miller, M.E., Xiao, H. and Wu, X., 2022. Potential health benefits of edible insects. Critical Reviews in Food Science and Nutrition 62: 3499-3508.
OâFlaherty, R., Bergin, A., Flampouri, E., Mota, L.M., Obaidi, I., Quigley, A. and Butler, M., 2020. Mammalian cell culture for production of recombinant proteins: a review of the critical steps in their biomanufacturing. Biotechnology Advances 43: 107552.
Pamboukian, M.M., Jorge, S.A.C., Santos, M.G., Yokomizo, A.Y., Pereira, C.A. and Tonso, A., 2008. Insect cells respiratory activity in bioreactor. Cytotechnology 57: 37. https://doi.org/10.1007/S10616-007-9118-8
Park, S., Kuo, J.C.H., Reesink, H.L. and Paszek, M.J., 2022. Recombinant mucin biotechnology and engineering. Advanced Drug Delivery Reviews 6: 114618.
Parthasarathy, R. and Palli, S.R., 2021. Stage-specific action of juvenile hormone analogs. Journal of Pesticide Science 46: 16. https://doi.org/10.1584/JPESTICS.D20-084
Paul, A., Frederich, M., Megido, R.C., Alabi, T., Malik, P., Uyttenbroeck, R. and Danthine, S., 2017. Insect fatty acids: a comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. Journal of Asia-Pacific Entomology 20: 337-340.
Pava-Ripoll, M., Pearson, R.E.G., Miller, A.K., Tall, B.D., Keys, C.E. and Ziobro, G.C., 2015. Ingested Salmonella enterica, Cronobacter sakazakii, Escherichia coli O157: H7, and Listeria monocytogenes: transmission dynamics from adult house flies to their eggs and first filial (F1) generation adults. BMC Microbiology 15: 1-12.
Payne, C., Caparros Megido, R., Dobermann, D., FreÌdeÌric, F., Shockley, M. and Sogari, G., 2019. Insects as food in the global north â the evolution of the entomophagy movement. Edible Insects in the Food Sector: Methods, Current Applications and Perspectives 5: 11-26.
Pinotti, L. and Ottoboni, M., 2021. Substrate as insect feed for bio-mass production. Journal of Insects as Food and Feed 7: 585-596.
Place, T.L., Domann, F.E. and Case, A.J., 2017. Limitations of oxygen delivery to cells in culture: an underappreciated problem in basic and translational research. Free Radical Biology and Medicine 113: 311-322.
Quattrociocchi, M., 2017. Partial characterization of yeast hydrolysates for insights on chemically defined media for Sf-9 insect cells. Masterâs thesis, University of Waterloo.
Raheem, D., Raposo, A., Oluwole, O.B., Nieuwland, M., Saraiva, A. and Carrascosa, C., 2019. Entomophagy: nutritional, ecological, safety and legislation aspects. Food Research International 126: 108672.
Raksakantong, P., Meeso, N., Kubola, J. and Siriamornpun, S., 2010. Fatty acids and proximate composition of eight Thai edible terricolous insects. Food Research International 43: 350-355. https://doi.org/10.1016/j.foodres.2009.10.014
Rao, K.M., Choi, S.M. and Han, S.S., 2023. A review on directional muscle cell growth in scaffolding biomaterials with aligned porous structures for cultivated meat production. Food Research International 5: 112755.
Reardon, K.F., 2021. Practical monitoring technologies for cells and substrates in biomanufacturing. Current Opinion in Biotechnology 71: 225-230.
Reid, W. and OâBrochta, D.A., 2016. Applications of genome editing in insects. Current Opinion in Insect Science 13: 43-54.
Reyes, S.J., Durocher, Y., Pham, P.L. and Henry, O., 2022. Modern sensor tools and techniques for monitoring, controlling, and improving cell culture processes. Processes 10: 180-189.
Rubio, N.R., Fish, K.D., Trimmer, B.A. and Kaplan, D.L., 2019a. In vitro insect muscle for tissue engineering applications. ACS Biomaterials Science and Engineering 5: 1071-1082. https://doi.org/10.1021/acsbiomaterials.8b01261
Rubio, N.R., Fish, K.D., Trimmer, B.A. and Kaplan, D.L., 2019b. Possibilities for engineered insect tissue as a food source. Frontiers in Sustainable Food Systems 3: 20-24. https://doi.org/10.3389/fsufs.2019.00024
Rumpold, B.A. and Schlüter, O.K., 2013. Nutritional composition and safety aspects of edible insects. Molecular Nutrition and Food Research 57: 802-823. https://doi.org/10.1002/mnfr.201200735
Sakadevan, K. and Nguyen, M.L., 2017. Livestock production and its impact on nutrient pollution and greenhouse gas emissions. Advances in Agronomy 141: 147-184.
Santana, A., Salido, E., Torres, A. and Shapiro, L.J., 2003. Primary hyperoxaluria type 1 in the Canary Islands: a conformational disease due to I244T mutation in the P11L-containing alanine:glyoxylate aminotransferase. Proceedings of the National Academy of Sciences of the United States of America 100: 7277-7282. https://doi.org/10.1073/pnas.1131968100
Scala, A., Cammack, J.A., Salvia, R., Scieuzo, C., Franco, A., Bufo, S.A. and Falabella, P., 2020. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae produced at an industrial scale. Scientific Reports 10: 19448.
Schneiderâs Drosophila Medium. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/21720024
Schoch, C.L., Ciufo, S., Domrachev, M., Hotton, C.L., Kannan, S., Khovanskaya, R., Leipe, D., McVeigh, R., OâNeill, K., Robbertse, B., Sharma, S., Soussov, V., Sullivan, J.P., Sun, L., Turner, S. and Karsch-Mizrachi, I., 2020. NCBI taxonomy: a comprehensive update on curation, resources and tools. Database 2020: baaa062. https://doi.org/10.1093/DATABASE/BAAA062
SeÌreÌ, A., Bougma, A., BazieÌ, B.S.R., TraoreÌ, E., Parkouda, C., GnankineÌ, O. and BassoleÌ, I.H.N., 2021. Chemical composition, energy and nutritional values, digestibility and functional properties of defatted flour, protein concentrates and isolates from Carbula marginella (Hemiptera: Pentatomidae) and Cirina butyrospermi (Lepidoptera: Saturniidae). BMC Chemistry 15: 46. https://doi.org/10.1186/s13065-021-00772-z
Sf-900TM II SFM. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/10902088
Sf-900TM III SFM. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/12659017
Shantibala, T., Lokeshwari, R.K. and Debaraj, H., 2014. Nutritional and antinutritional composition of the five species of aquatic edible insects consumed in Manipur, India. Journal of Insect Science 14: 1-10. https://doi.org/10.1093/jis/14.1.14
Shi, N. and Chen, S.Y., 2014. Mechanisms simultaneously regulate smooth muscle proliferation and differentiation. Journal of Biomedical Research 28: 40.
Shriner, S.A. and Root, J.J., 2020. A review of avian influenza A virus associations in synanthropic birds. Viruses 12: 1209.
Siddiqui, S.A., Khan, S., Farooqi, M.Q.U., Singh, P., Fernando, I. and Nagdalian, A., 2022. Consumer behavior towards cultured meat: a review since 2014. Appetite 179: 106314. https://doi.org/10.1016/j.appet.2022.106314
Simoni, A., Siniscalchi, C., Chan, Y.S., Huen, D.S., Russell, S., Windbichler, N. and Crisanti, A., 2014. Development of synthetic selfish elements based on modular nucleases in Drosophila melanogaster. Nucleic Acids Research 42: 7461-7472.
Skora, L., Shrestha, B. and Gossert, A.D., 2015. Isotope labeling of proteins in insect cells. In: Kelman, Z. (ed.) Methods in enzymology. Academic Press, Cambridge, MA, USA, pp. 245-288. https://doi.org/10.1016/bs.mie.2015.05.013
Smetana, S., Bhatia, A., Batta, U., Mouhrim, N. and Tonda, A., 2023. Environmental impact potential of insect production chains for food and feed in Europe. Animal Frontiers 13: 112-120.
Smetana, S., Palanisamy, M., Mathys, A. and Heinz, V., 2016. Sustainability of insect use for feed and food: life cycle assessment perspective. Journal of Cleaner Production 137: 741-751.
Stanley, D.W., Goodman, C., An, S., McIntosh, A. and Song, Q., 2008. Prostaglandins A1 and E1 influence gene expression in an established insect cell line (BCIRL-HzAM1 cells). Insect Biochemistry and Molecular Biology 38: 275-284. https://doi.org/10.1016/J.IBMB.2007.11.004
Stolt-Bergner, P., Benda, C., Bergbrede, T., Besir, H., Celie, P.H., Chang, C. and Suppmann, S., 2018. Baculovirus-driven protein expression in insect cells: a benchmarking study. Journal of Structural Biology 203: 71-80.
Strobl, F., Duerkop, M., Palmberger, D. and Striedner, G., 2021. High shear resistance of insect cells: the basis for substantial improvements in cell culture process design. Scientific Reports 11: 1-11. https://doi.org/10.1038/s41598-021-88813-4
Su, Y., Lu, M.X., Jing, L.Q., Qian, L., Zhao, M., Du, Y.Z. and Liao, H.J., 2021. Nutritional properties of larval epidermis and meat of the edible insect Clanis bilineata tsingtauica (Lepidoptera: Sphingidae). Foods 10: 2895. https://doi.org/10.3390/foods10122895
Sugii, S., Wong, C.Y.Q., Lwin, A.K.O. and Chew, L.J.M., 2023. Alternative fat: redefining adipocytes for biomanufacturing cultivated meat. Trends in Biotechnology 41: 686-700. https://doi.org/10.1016/J.TIBTECH.2022.08.005
Svensson, I., (n.d.). Physiological effects of conditioned medium and passage number on Spodoptera frugiperda Sf9 serum free cultures. Available at: https://pubmed.ncbi.nlm.nih.gov/16599552/
Tauber, J.P., Nguyen, V., Lopez, D. and Evans, J.D., 2019. Effects of a resident yeast from the honeybee gut on immunity, microbiota, and Nosema disease. Insects 10: 296.
Taylor, M.J., Weegman, B.P., Baicu, S.C. and Giwa, S.E., 2019. New approaches to cryopreservation of cells, tissues, and organs. Transfusion Medicine and Hemotherapy 46: 197-215.
TC-100 insect Medium. (n.d.). Available at: https://www.thermofisher.com/order/catalog/product/de/en/13055025
The buzzing future of food: an introduction to entomoculture | protein report. (n.d.). Available at: https://www.proteinreport.org/articles/buzzing-future-food-introduction-entomoculture/
Tognato, R., Armiento, A.R., Bonfrate, V., Levato, R., Malda, J., Alini, M. and Serra, T., 2019. A stimuli-responsive nanocomposite for 3D anisotropic cell-guidance and magnetic soft robotics. Advanced Functional Materials 29: 1804647.
Torino, S., Corrado, B., Iodice, M. and Coppola, G., 2018. Pdms-based microfluidic devices for cell culture. Inventions 3: 65.
Treich, N., 2021. Cultured meat: promises and challenges. Environmental and Resource Economics 79: 33-61. https://doi.org/10.1007/s10640-021-00551-3
Tyagi, S., Kumar, R., Das, A., Won, S.Y. and Shukla, P., 2020. CRISPR-Cas9 system: a genome-editing tool with endless possibilities. Journal of Biotechnology 319: 36-53.
United Nation, 2023. Population. Available at: https://www.un.org/en/global-issues/population
US7829309B1 â cell culture performance with betaine â Google patents. (n.d.). Available at: https://patents.google.com/patent/US7829309B1/en
Van Broekhoven, S., Oonincx, D.G., Van Huis, A. and Van Loon, J.J., 2015. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. Journal of Insect Physiology 73: 1-10.
Van Huis, A., 2017. Edible insects and research needs. Journal of Insects as Food and Feed 3: 3-5.
van Huis, A., 2020. Edible insects. In: Meiselman, H. (ed.) Handbook of eating and drinking. Springer, Cham, Swotserland. https://doi.org/10.1007/978-3-030-14504-0_123
Van Huis, A. and Oonincx, D.G., 2017. The environmental sustainability of insects as food and feed. A review. Agronomy for Sustainable Development 37: 1-14.
Vauterin, A., Steiner, B., Sillman, J. and Kahiluoto, H., 2021. The potential of insect protein to reduce food-based carbon footprints in Europe: the case of broiler meat production. Journal of Cleaner Production 320: 128799.
Vergunst, F. and Savulescu, J., 2017. Five ways the meat on your plate is killing the planet. The Conversation. Available at: https://theconversation.com/five-ways-the-meat-on-your-plate-is-killing-the-planet-76128
Wang, G.H., Gamez, S., Raban, R.R., Marshall, J.M., Alphey, L., Li, M. and Akbari, O.S., 2021. Combating mosquito-borne diseases using genetic control technologies. Nature Communications 12: 388.
Wang, Y., Oberley, L.W. and Murhammer, D.W., 2001. Antioxidant defense systems of two lipidopteran insect cell lines. Free Radical Biology and Medicine 30: 1254-1262. https://doi.org/10.1016/S0891-5849(01)00520-2
Xu, T., Jiang, X., Denton, D. and Kumar, S., 2020. Ecdysone controlled cell and tissue deletion. Cell Death and Differentiation 27: 1-14. https://doi.org/10.1038/S41418-019-0456-9
Yang, H., Xiong, X., Li, T. and Yin, Y., 2016. Ethanolamine enhances the proliferation of intestinal epithelial cells via the mTOR signaling pathway and mitochondrial function. In vitro cellular & developmental biology. Animal 52: 562-567. https://doi.org/10.1007/S11626-016-0002-8
Zhang, C., Guan, X., Yu, S., Zhou, J. and Chen, J., 2022a. Production of meat alternatives using live cells, cultures and plant proteins. Current Opinion in Food Science 43: 43-52. https://doi.org/10.1016/J.COFS.2021.11.002
Zhang, J., Zhang, Y., Li, J., Liu, M. and Liu, Z., 2016. Midgut transcriptome of the cockroach Periplaneta americana and its microbiota: digestion, detoxification and oxidative stress response. PLOS ONE 11: e0155254.
Zhang, X., Feng, H., He, J., Muhammad, A., Zhang, F. and Lu, X., 2022b. Features and colonization strategies of enterococcus faecalis in the gut of Bombyx mori. Frontiers in Microbiology 13: 921330.
Zhang, Y., Yu, R., Tang, J., Du, L., Wang, Y., Wang, J. and Li, B., 2022c. Three cytochrome P450 CYP4 family genes regulated by the CncC signaling pathway mediate phytochemical susceptibility in the red flour beetle, Tribolium castaneum. Pest Management Science 78: 3508-3518.
Zhao, D., Wu, H., Li, Y., Wang, Q., Ji, Y., Guo, X. and Guo, W., 2023. Effects of the pyrE deletion mutant from Bacillus thuringiensis on gut microbiota and immune response of Spodoptera exigua. Frontiers in Microbiology 14: 1182699.
Zhou, Y., Wang, D., Zhou, S., Duan, H., Guo, J. and Yan, W., 2022. Nutritional composition, health benefits, and application value of edible insects: a review. Foods 11: 3961. https://doi.org/10.3390/foods11243961
ZidaricÌ, T., MilojeviÌc, M., Vajda, J., Vihar, B. and Maver, U., 2020. Cultured meat: meat industry hand in hand with biomedical production methods. Food Engineering Reviews 12: 498-519.
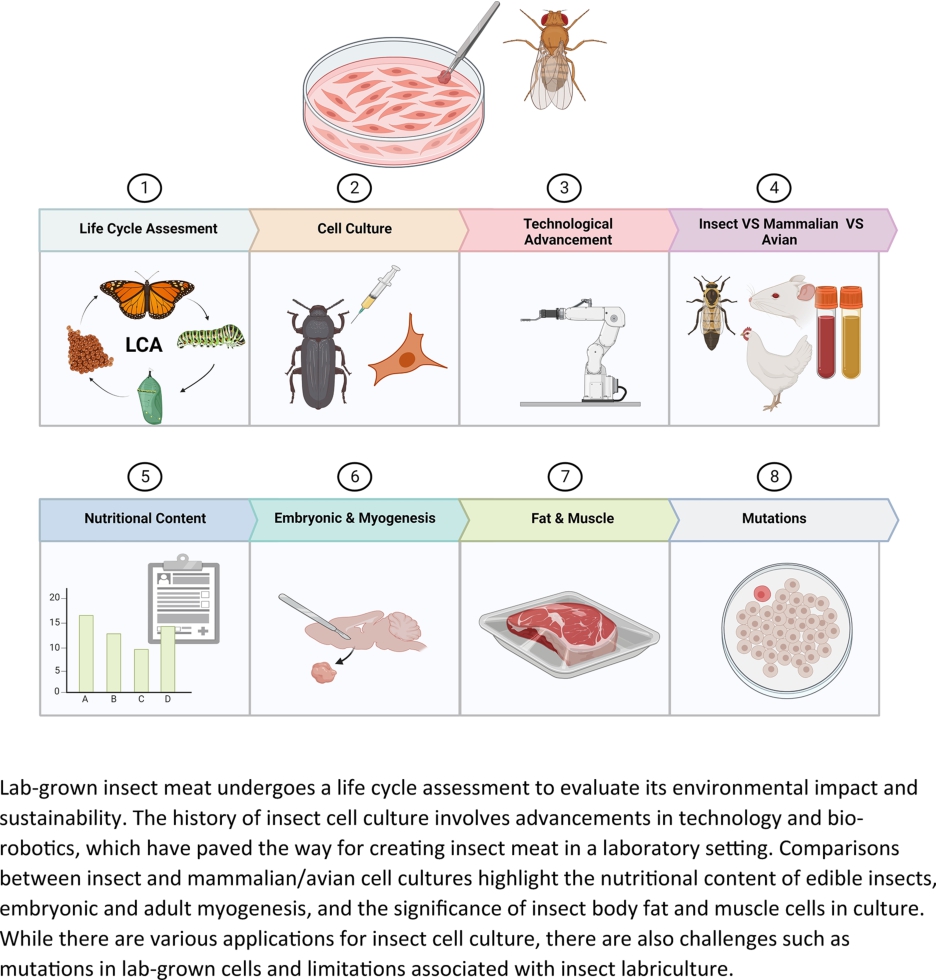
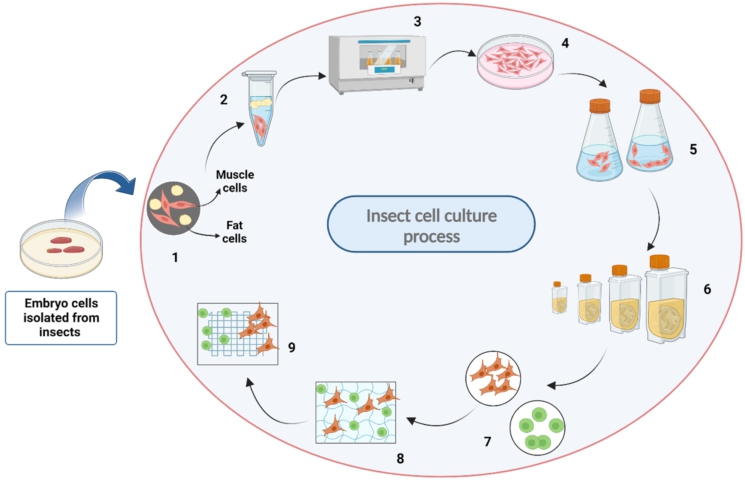



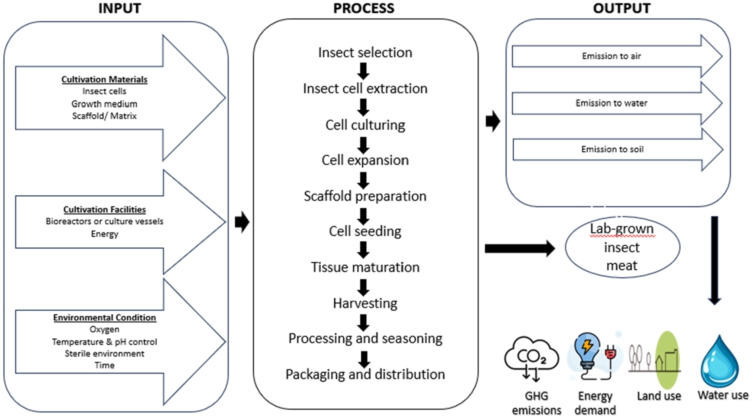


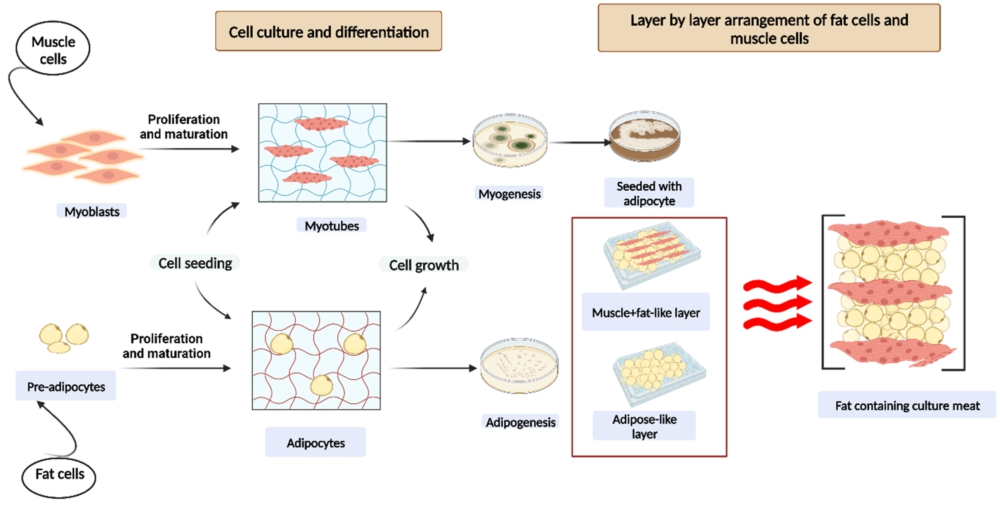


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}