Abstract
In this study, black soldier fly (BSF) dried larvae were fractionated into their defatted, lipid, protein, and chitin-rich fractions. The samples were then characterised in terms of proximate composition, analysis of amino acids, analysis of the molecular weight distribution of proteins via gel electrophoresis, and fatty acids analysis of the lipid fraction. The antibacterial activity of all fractions was determined against Escherichia coli F4 and Streptococcus suis, two common swine pathogens. Finally, the prebiotic activity of the whole larvae and of their protein isolate and chitin-rich pellet was determined for Limosilactobacillus reuteri, a naturally occurring species in the gastro-intestinal tract of the pig. The results on amino acid profile showed that the protein extraction method used was not fully effective in cleaving chitin-bound proteins, as expected, as the true protein content of the chitin-rich sample was 48.5%. This was in accordance with the SDS-PAGE analysis, where the molecular weight distribution of proteins in the protein isolate sample did not show band at higher molecular weight, which were instead present in the chitin-rich sample. The analysis of fatty acid (FA) profile showed that lauric acid was the most abundant FA, in accordance with the literature. The results on microbiological analyses highlighted how all nitrogen-containing samples showed antibacterial activity against E. coli, the highest being the defatted fraction at 1% with a decrease of −0.77 log CFU/mL, while only the lipid fraction against S. suis (−0.48 log CFU/mL at 1%). All tested samples showed prebiotic activity for L. reuteri, but it was noticed that the increase in chitin concentration resulted in the decrease of the prebiotic effect. By combining the results of both antibacterial and prebiotic tests, it could be inferred that chitin might be considered as an antibacterial compound.
This article is dedicated to the memory of Stefano Sforza
1 Introduction
The sustainability of the animal breeding sector is highly determined by the natural resources used for feed production (Hawkey et al., 2021; Sogari et al., 2019; Spranghers et al., 2018). Soybean is currently the most used protein source in animal feed worldwide; the widespread monoculture use of this crop has led to a worrying increase in deforestation and to biodiversity losses. Therefore, interest in novel, more sustainable sources of protein for animal feed has recently grown (Pam Ismail et al., 2020; Spranghers et al., 2018).
Another issue related with animal husbandry is the inappropriate use of antibiotics by livestock farmers, which has led to antimicrobial resistance (AMR) (Xia et al., 2021). Despite the fact that nowadays the use of antimicrobials as growth promoters in feed is forbidden in many areas (Auza et al., 2020), AMR is still on the rise worldwide (Choi et al., 2012).
AMR is defined as the acquired resistance of pathogens to antimicrobial agents (antibacterials, antifungals, antivirals). This process naturally takes place over time, but practices such as the improper use of these drugs may accelerate the phenomenon (Murray et al., 2022; World Health Organization, 2017). As a matter of fact, the abuse of antimicrobials is considered the main factor contributing to AMR spread, in animals and humans (Jadeja and Worrich, 2022).
Since the second half of the 20th century, antimicrobials have been extensively used, as they were supposed to promote livestock productivity, and nowadays more than 70% of antibiotics sold worldwide are used for animal husbandry (Jadeja and Worrich, 2022; Zhu et al., 2022). As a result of the use and abuse of antimicrobial drugs, several strains of swine-derived pathogens have been found to be drug-resistant to a multiple range of antibiotics, representing a hazard for both animals and humans. In 2017, the World Health Organization (WHO) reported a list of “priority pathogens”, which have been clustered based on the urgency of need for new antibiotics; among these, Enterobacteriaceae such as Escherichia coli are considered of critical priority (Jadeja and Worrich, 2022; Zhu et al., 2022).
Due to this, in recent years there has been a growing interest towards discovering new and novel alternatives to antibiotics. Among these, the use of probiotics and prebiotics has gained attention as a potential strategy, as they have showed several benefits when used in pigs’ diet. In fact, they demonstrated to regulate and improve the growth of the intestinal microbiota, prevent pathogenic infections in the gastrointestinal tract, improve digestion, and produce antimicrobial compounds (Dowarah et al., 2017; Hou et al., 2015; Yang et al., 2015). Another alternative to traditional antimicrobials is the exploitation of naturally occurring compounds with antimicrobial activity; these can include plant extracts, such as curcumin, animal-derived extracts and products, such as chitosan and honey, and extracts obtained from microorganisms, the majority of which derive from moulds and actinobacteria. More recently, insect-derived extracts have been of interest, as their innate immune system is extensively made up of antimicrobial peptides (Chen et al., 2023; Ghosh et al., 2008; Koutsos et al., 2019; Taylor, 2013).
Insect farming has been gaining attention for many reasons: they represent a great source of essential nutrients such as high-quality proteins and polyunsaturated fatty acids, they can be reared on a multitude of organic waste and efficiently convert it into valuable biomass, and they need a relatively low amount of resources in terms of use of water, land, and energy to be reared (Hawkey et al., 2021; Leni et al., 2021; van Huis and Gasco, 2023). Moreover, they contain bioactive compounds with potential antimicrobial activity, such as lauric acid and antimicrobial peptides, and chitin, which might be a potential novel prebiotic (Auza et al., 2020; Refael et al., 2022; Sogari et al., 2019; Xia et al., 2021). The BSF is among the most promising species for insect farming: its high feed conversion rate and short reproductive cycle allow to obtain biomass of high nutritional value in a short span of time. Moreover, BSF has low maintenance costs, since it can grow easily on many substrates derived from agri-food side streams (Lu et al., 2022).
From the available data in the literature, there is no clarity as to which type of bacteria can be mostly inhibited by BSF whole or fractionated larvae: a number of studies showed that protein extracts and antimicrobial peptides obtained from BSF larvae inhibited the growth of gram-negative bacteria (Auza et al., 2020; Harlystiarini et al., 2019; Van Moll et al., 2022), but not gram-positive ones (Choi et al., 2012), while other authors observed antibacterial activity of whole BSF digesta against gram-positive bacteria (Dong et al., 2021). In addition, different studies demonstrated the efficacy of the lipid fraction of BSF against gram-positive bacteria, mainly related to the high lauric acid content (Hurtado-Ribeira et al., 2023; Li et al., 2022; Zabulionė et al., 2023). Studies have shown that the efficacy of antimicrobial peptides on gram-negative or positive bacteria depends largely on their amino acid profile: specifically, cysteine-rich peptides were shown to be mainly active against gram-positive bacteria, while peptides rich in glycine residues against gram-negative ones (Veldkamp et al., 2022).
Notably, relatively little to no research has been carried out on livestock pathogens, thus emphasizing the importance of exploring this topic. Moreover, there is a gap in the research concerning the characterization of BSF main components (protein, lipid, and chitin fractions) in relation to their potentially antibacterial and prebiotic compounds. In fact, to our knowledge, no study has ever attempted to determine the antibacterial and prebiotic activity of the whole larvae and its fractions, and further characterise the isolated compounds that may contribute to their antibacterial and prebiotic potential.
Therefore, in this study, Hermetia illucens L. whole and defatted larvae, and their extracted protein, lipid, and chitin-rich fractions underwent in-depth molecular characterization, and their antibacterial and prebiotic activity were evaluated. Antibacterial tests were carried out against a gram-negative bacterium, Escherichia coli F4, and a gram-positive one, Streptococcus suis, being among the most common swine pathogens (Aarestrup et al., 2008; Abubakar et al., 2019; Obradovic et al., 2021). Prebiotic tests were carried out with Limosilactobacillus reuteri, being one of the few naturally occurring species identified in the gastro-intestinal tract of pigs (Hou et al., 2015).
2 Materials and methods
Solvents and reagents
The RAPID’E. coli 2 agar, the Criterion XT Bis-Tris Gel at 10%, and the buffer XT MES 20× were supplied by the company BioRad (Hercules, CA, USA). The Dithiothreitol (DTT) was supplied by the company ITW Reagents Division by PanReac AppliChem (Darmstadt, Germany). Bromocresol Green, Ethylenediaminetetraacetic acid (EDTA), L-Ascorbic acid, TRIS Hydrochloride, Sodium chloride, Sodium hydroxide, DL-Norleucine, L-Cysteic acid monohydrate, DL-Methionine sulfone, Chitin from shrimp shells, L-Tryptophan, α-Methyl-DL-Tryptophan, D-(+)-Galactosamine hydrochloride, and ammonium bicarbonate were supplied by Sigma-Aldrich Co. (St. Louis, MO, USA). The Methyl Red ACS reagent, the brain heart infusion broth, the KF Streptococcus agar, the MRS Broth (DeMan, Rogosa and Sharpe), and the MRS Agar were supplied by Thermo Fisher Scientific Inc. (Brussels, Belgium). Copper (II) oxide powder, sulfuric acid 95%, Kjeldahl Catalyst CK 3.9 g, Kjeldahl Catalyst KS 5.05 g, boric acid, sodium hydroxide, hydrochloric acid 0.1 mol/l (0.1 N), hydrochloric acid 0.01 mol/l (0.01 N), acetic acid, urea, diethyl ether, acetonitrile, methanol, and ethanol were supplied by VWR International (Milan, Italy). The AccQ-Fluor Reagent Kit and the AccQ∙Tag™ were supplied by Waters (Milford, MA, USA).
Samples
Hermetia illucens L. dried larvae were purchased from the company Société APPI (Pont St Martin, France). The larvae were stored in ziplock bags at room temperature and in the dark until use. Before carrying out the analyses, the larvae were finely ground with the GRINDOMIX GM 200 knife mill (Retsch, Germany).
Bacterial strains and culture conditions
Escherichia coli F4 was supplied by the company SSI Diagnostica (Leiden, the Netherlands). Streptococcus suis was supplied by the Animal Health Care Flanders association (Dierengezondheidszorg Vlaanderen – DGZ, Torhout, Belgium). Limosilactobacillus reuteri supsp. porcinus was supplied the Belgian Coordinated Collections of Microorganisms (BCCM). To obtain plated pure bacterial culture, aliquots of the liquid pure bacterial cultures were incubated in 10 mL of brain heart infusion (BHI) broth in case of the pathogenic bacteria, or MRS broth in case of lactic acid bacteria (LAB), both previously prepared according to the manufacturer’s instruction, at 37 °C for 24 hours (E. coli F4) and 48 hours (S. suis and L. reuteri). Then, the bacterial solutions were plated on E. coli agar for E. coli, on Streptococcus agar for S. suis, and on MRS agar for L. reuteri. The plates were incubated for 24 hours at 37 °C for E. coli, and for 48 hours at 37 °C for S. suis and L. reuteri. Finally, they were stored as pure cultures on Petri dishes at 4 °C until use. Before each analysis, one bacterial colony from the plated pure cultures was incubated in 10 mL of BHI broth in case of the pathogenic bacteria, or MRS broth in case of LAB, both previously prepared according to the manufacturer’s instruction, at 37 °C for 24 hours (E. coli F4) and 48 hours (S. suis and L. reuteri), to obtain a fresh stock of concentrated bacterial solution of 107 to 108 CFU/mL. This was then diluted ×100 or ×1000 in physiological salt solution and used for the inocula in antibacterial and prebiotic tests, respectively. The number of colonies was determined by plating method on E. coli agar for E. coli F4, after incubating the plates for 24 hours at 37 °C, on Streptococcus agar for S. suis, after incubating the plates for 48 hours at 37 °C, and on MRS agar for L. reuteri, after incubating the plates for 48 hours at 37 °C.
Fractionation of proteins, lipids, and chitin
BSF dried larvae were finely ground with the GRINDOMIX GM 200 knife mill (Retsch, Germany). Then, to facilitate further protein extraction and to isolate the lipid fraction, a first step of lipid extraction following the Folch extraction procedure was performed with the modifications previously reported by Caligiani et al., 2019. The lipids extracted were collected as the isolated lipid fraction and stored at −20 °C until use, together with a part of the defatted samples. On the defatted sample, a mild stepwise protein extraction, based on Osborne fractionation, was then carried out following the method proposed by Leni et al. (2019), with some modifications: specifically, the sample:water ratio was reduced to 1:2.5 (w/v), and the globulin supernatant underwent a dialysis step with the Sigma-Aldrich dialysis tubing (pore size 2000 Da), as it was the extract containing the highest amount of salts. The four supernatants were jointly collected to obtain the isolated protein fraction, freeze-dried, and stored at −20 °C until use. Finally, chitin was obtained by demineralization of the residual pellet. Specifically, the deproteinised pellet was subjected to demineralization with 40 mL of HCl 2 M for 24 hours at room temperature. The sample was then centrifuged at 4,000 rpm for 15 minutes. The precipitate was washed with deionised water until pH neutrality and the final pellet was dried overnight at 40 °C. The dried pellet was collected as the isolated chitin-rich fraction and stored at −20 °C until use. The fractions were characterised as more comprehensively described in the following sections.
Proximate composition analysis
The proximate composition analyses were carried out on the whole larvae and on their defatted, protein, and chitin-rich fractions following standard methods (AOAC, 2002). For a precise measurement of true protein, thus excluding non-protein sources of nitrogen, protein content was determined by UPLC-ESI/MS analysis of amino acids as suggested by Caligiani et al. (2018), following the method described by Luparelli et al. (2023). Lipid content was determined by Soxhlet method using the VELP SCIENTIFICA fat extractor and diethyl ether as solvent. Ash content was determined gravimetrically after demineralization at 550 °C for 5 hours. In addition, chitin content was also determined following a method developed by Luparelli et al. (2023) based on an LC/MS determination of the chitin monomer N-acetylglucosamine after acid hydrolysis, as more comprehensively described in section “Analysis of amino acids and chitin”.
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE)
SDS-PAGE was carried out following methods described in previous studies (Leni et al., 2019). The samples were analysed with the Criterion XT Bis-Tris Gel at 10% (Biorad, Hercules, CA, USA) using MES 20× running buffer. The image of the obtained gel was acquired on a GS-800TM scanner with the QuantityOne BIO-RAD software.
Analysis of fatty acids
The analysis of fatty acids was carried out on the lipid extract. Briefly, 50 mg of lipid extract underwent an acid-catalysed transmethylation by adding 0.5 mL of sulfuric acid/methanol (1:15 v/v) in glass tubes. The samples were then heated at 100 °C for 3 hours. After cooling, 5 mL of hexane containing tetracosane at concentration 0.02 mg/mL as internal standard were added to the tubes. Finally, the samples were diluted 1:1 v/v with hexane and analysed on the Agilent Technologies 7820A gas-chromatograph (Agilent technologies, Palo Alto, CA, USA) coupled with the Agilent Technologies 5977B mass spectrometer using the low polarity capillary SLB-5MS column (30 m × 250 μm i.d., film thickness 0.25 μm) (Supelco, Bellefonte, PA, USA) with the following experimental conditions: inlet temperature: 280 °C. Carrier gas: helium at constant flow rate of 1 mL/min. Oven temperature: isothermal at 60 °C for 2 minutes, from 60 to 220 °C at a constant rate of 20 °C/min, isothermal at 220 °C for 8 minutes, from 220 to 280 °C at a constant rate of 20 °C/min, isothermal for 4 minutes. Mass spectrometer: electron impact (EI) ionization mode, 70 eV, scan range 40-400 m/z, ion source temperature 230 °C. The fatty acid content was determined in relation to the concentration of the internal standard, using the Supelco 37 Component FAME Mix as response factor, and expressed as relative percentage of fatty acid.
Analysis of amino acids and chitin
Quantification of amino acids and chitin was carried out on the whole larvae and on the defatted, protein, and chitin fractions, following procedures described in previous studies (Luparelli et al., 2023; Prandi et al., 2021), with some modifications. Briefly, 150 mg of samples were hydrolysed with 6 mL of HCl 6 M in Pyrex glass tubes for 23 h at 110 °C. Then, 0.75 mL of norleucine 50 mM in HCl 0.1 M were added to each tube as internal standard for the quantification of amino acids. The mixtures were filtered with Buchner filters in 100 mL volumetric flasks and brought up to volume with ultrapure water. 900 μL of filtered and diluted sample solution were mixed with 86 μL of ultrapure water and 14 μL of galactosamine 46 mM in HCl 0.1 M as internal standard for the quantification of chitin.
For the quantification of sulphur-containing amino acids, a preliminary treatment with performic acid to force their complete oxidation was carried out, and cysteine and methionine were thus quantified as their relative oxidated form (cystic acid and methionine sulphone, respectively). Specifically, 50 mg of samples were added to 0.2 mL of freshly prepared performic acid and incubated for 16 h in an ice bath. Subsequently, 0.03 mL of hydrobromic acid 48% were added to remove the excess of performic acid, and the samples were dried under nitrogen. Acid hydrolysis was performed as previously described by adding 0.6 mL of HCl 6 M in each tube. Then, 0.075 mL of norleucine 50 mM in HCl 0.1 M were added to each tube as internal standard. The mixtures were filtered with Buchner filters in 25 mL volumetric flasks and brought up to volume with ultrapure water.
A calibration curve for the quantification of amino acids, sulphur-containing amino acids, and chitin was prepared as previously reported by Luparelli et al. (2023), with some modifications. Specifically, in addition to the Amino Acid Standard H (Waters, Milford, MA, USA), norleucine, chitin, and galactosamine, also methionine sulfone and cystic acid were added in the same concentration as the other amino acids.
Finally, the final sample mixtures, as well as the calibration curve, were derivatised with the AccQ∙Tag Fluor Derivatization Kit (Waters, Milford, MA, USA) according to the manufacturer’s instructions and transferred into vials for UPLC analysis. Then, UPLC-ESI/MS analysis was performed using an UPLC separation system coupled to a single quadrupole SWD detector (Waters, Milford, USA) with the following experimental conditions:
- ∙ Stationary phase: Acquity UPLC Protein BEH C18 column (300Å, 1.7 μm, 2.1 mm × 150 mm) with Acquity UPLC Peptide CSH™ C18 VanGuard™ (130Å, 1.7 μm, 2.1 mm × 5 mm)
- ∙ Mobile phase: H2O Milli-Q + 0.2% CH3CN + 0.1% HCOOH (eluent A) and CH3CN + 0.1% HCOOH (eluent B).
- ∙ Instrumental parameters: column temperature 35 °C, injected volume 5 μl, flow 0.25 mL/min, ionization mode ESI+, capillary voltage 3.2 kV, cone voltage 30 V, source temperature 150 °C, desolvation temperature 300 °C, cone gas flow 100 L/Hr, desolvation gas flow 650 L/Hr
Information on gradient elution and method validation are reported in the study by Luparelli et al. (2023).
Analysis of tryptophan
The analysis of tryptophan has been carried out following the method proposed by Cutroneo et al. (2023), with some modifications. Briefly, the samples were subjected to alkaline hydrolysis by mixing 150 mg of samples to 4 mL of NaOH 4 M at 100 °C for 4 hours. 150 μl of α-methyl-tryptophan 0.5 mg/mL were also added as internal standard, and the tubes were fluxed with nitrogen for one minute before hydrolysis to avoid tryptophan oxidation. The mixtures were then neutralised using HCl 6 M and centrifuged at 4,000 rpm and 4 °C for 45 minutes. The supernatants were filtered with syringe filters of pore size 0.45 μm in 10 mL volumetric flasks, brought to volume with ultrapure water and transferred into vials for UPLC analysis. For the quantification of tryptophan, a response factor constituted by tryptophan and α-methyl-tryptophan with concentration 0.5 mg/mL was prepared in duplicate for UPLC analysis. Then, UPLC-ESI/MS analysis was performed with experimental conditions described by Cutroneo et al. (2023).
Determination of the antibacterial activity
The bacteria under investigation were Escherichia coli F4 and Streptococcus suis. The samples tested were black soldier fly dried larvae and their defatted, protein, lipid, and chitin fractions, obtained as previously described in section “Fractionation of proteins, lipids, and chitin”. Three biological replicates were made for each fraction and each bacterium.
Determination of the prebiotic activity
The samples tested were black soldier fly dried larvae and their protein and chitin fractions, obtained as previously described in section “Fractionation of proteins, lipids, and chitin”. Three biological replicates were obtained for each fraction.
Two types of broths were prepared: a normal MRS broth, prepared according to the manufacturer’s instructions, and a nutrient deprived MRS broth, prepared with the same solute quantities stated by the MRS broth manufacturer and excluding solutes containing carbon and nitrogen.

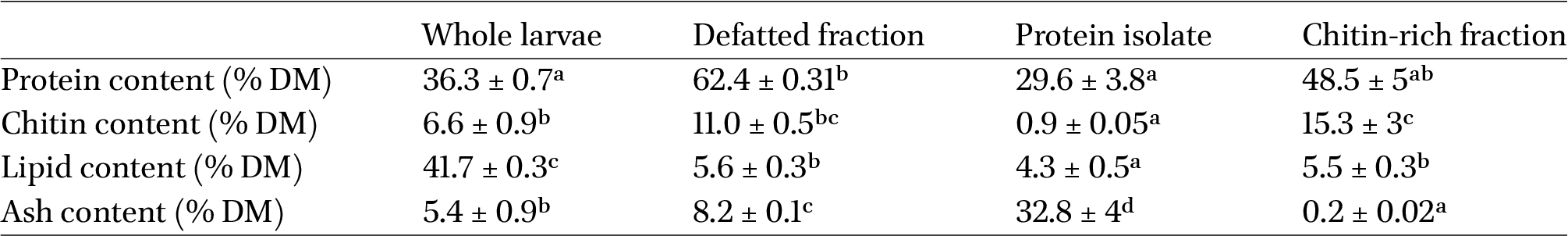
Proximate composition of black soldier fly whole larvae and of its defatted, protein, and chitin-rich fractions. The results are expressed on a dry matter basis (DM). Significant differences within each line are indicated by distinct letters (one-way ANOVA with post hoc Tukey b’s test,
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
Different tubes were set up in each test: a positive control tube, containing 10 mL of MRS broth; a negative control tube, containing 10 mL of nutrient-deprived MRS broth; and two sample tubes, containing the nutrient-deprived MRS broth and 42 g/L (100%) or 21 g/L (50%) of insect sample, corresponding to the mass of N- and C-containing solutes missing. Then, 100 μL of the 1000× diluted bacterial culture were inoculated in each tube, the negative control was plated at time 0 h (T0), and both the plates and the tubes were incubated at 37 °C for 48 hours. After the incubation time, T0 colonies were counted, all tubes were plated at time 48 h (T48) and incubated at 37 °C for 48 hours. Finally, T48 colonies were counted and the bacterial growth, expressed as log10 CFU/mL was determined.
Statistical analysis
Data analysis was performed using the software IBM® SPSS® Statistics version 28.0.1.1 (SPSS Inc., Chicago, IL, USA). The data was subjected to one-way analysis of variance (ANOVA) test followed by Tukey b’s post hoc test (
3 Results and discussion
Proximate composition of black soldier fly larvae and of their defatted and lipid fractions, protein isolate, and chitin-rich pellet
Proximate composition analyses were carried out on the whole larvae and on their defatted, protein, and chitin-rich fractions following standard methods (European Commission (EC), 2009). The Folch extraction procedure was preferred over the more commonly used Soxhlet method as it allowed to better extract the lipid fraction from animal tissues and from larger quantities of sample. The Osborne fractionation method, typically employed for cereal proteins and adapted by Leni et al. (2018) for insect matrices, was chosen over other procedures as it lowered considerably protein degradation (Leni et al., 2028). The protein content was determined by UPLC-ESI/MS analysis of amino acids instead of Kjeldahl method to avoid the overestimation of proteins depending on the standard nitrogen-to-protein conversion factors (Caligiani et al., 2018) and to simultaneously determine the chitin content of all samples.
The results are reported in Table 1.
Overall, the proximate composition of the whole and defatted larvae was in accordance with studies found in the literature (Vanqa et al., 2022; Zhou et al., 2022). The ash content of the protein isolate was 32.8%, showing that a remarkable amount of residual salts from the extraction steps was present, as expected, thus indicating that it would be appropriate to add a dialysis step on the collected protein fractions to obtain a purer isolate. Regarding the chitin-rich pellet, a high percentage of residual proteins (48.5%) was still present. As it is already known, chitin is not found in its pure form in nature, but it’s normally bound to other components, mainly proteins and minerals, which might hinder its complete extraction; in fact, chitin binds tightly to cuticular proteins after the sclerotization process, thus critically limiting the separation of chitin-bound proteins (Eggink and Dalsgaard, 2023; Rangel et al., 2024; Tetreau et al., 2015).
GC-MS determination of fatty acids profile of the lipid fraction
The results, expressed as relative percentage of fatty acid in relation to the concentration of the internal standard, are shown in Table 2.
Fatty acids profile of black soldier fly lipid fraction. The results are expressed as relative percentage of fatty acid in relation to the concentration of the internal standard
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
The most abundant fatty acid was lauric acid (C12:0), accounting for almost 32% of the total fatty acids. Lauric acid is a medium-chained fatty acid commonly found in coconut and palm kernel oils, and it has been reported to show antibacterial activity against a range of microorganisms such as bacteria and viruses. Due to these characteristics, the interest in novel lauric acid sources, among which black soldier fly, has recently grown as an alternative strategy to conventional antimicrobials (Borrelli et al., 2021; Matsue et al., 2019). Other saturated fatty acids contained in significant amounts were myristic acid (C14:0) and palmitic acid (C16:0), accounting for 10.23 and 14.46% of the total fatty acids, respectively. In addition, linoleic (C18:2) and oleic (C18:1) acids were also present, being, respectively, at 19.75 and 13.70% of the total fatty acids. The high percentage of polyunsaturated fatty acids may be attributed to the quality of the feed on which the insects were reared, as previously reported (Danieli et al., 2019; Siddiqui et al., 2022). In fact, despite the protein content is usually not much influenced by the diet, the fatty acids profile can in fact be more easily modified and improved through diet (van Huis, 2021). Overall, the fatty acids profile was in line with the literature (Caligiani et al., 2019; Ewald et al., 2020; Li et al., 2022; Suryati et al., 2023).
Profiling of black soldier fly larvae protein extracts
Electrophoretic profile
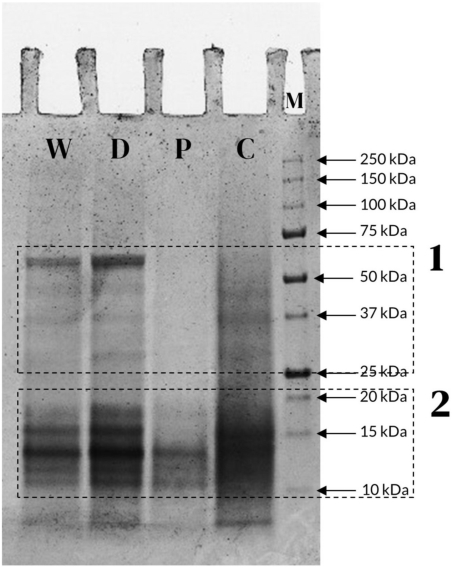
In order to assess the molecular weight distribution of the samples, SDS-PAGE gel electrophoresis was carried out as described in section “Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE)”. The electrophoretic profile is shown in Figure 1.
SDS-PAGE gel electrophoresis of black soldier fly whole larvae (W), defatted larvae (D), protein isolate (P), and chitin-rich pellet (C). The marker lane is indicated by the letter “M”. Region 1 includes bands in the range 75-25 kDa, while Region 2 includes bands below 25 kDa.
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
Multiple protein bands were visible in the range 75-10 kDa. Two main molecular weight (MW) regions were identified, in accordance with previous works on black soldier fly and on other insect species: one at higher MW, between 75 and 25 kDa, which might be attributed to structural proteins, muscle proteins, and enzymes (region 1), and one at lower MW, below 25 kDa, which might be constituted by haemolymph proteins and larval cuticle proteins (region 2) (Leni et al., 2019; Pasini et al., 2022; Queiroz et al., 2021; Rabani et al., 2019; Xu et al., 2023; Yi et al., 2013).
By comparing the whole (W) and defatted (D) larvae it can be observed that the protein profile remained unchanged, thus indicating that the separation process of the lipid fraction through Folch method did not affect the protein fraction in the defatted sample. The protein isolate (P) only showed protein bands at MW below 25 kDa, suggesting that the extraction process was not effective for proteins with higher MW, while both high and low MW proteins were present in the chitin-rich pellet (C). Additionally, when observing the latter, it can be seen that the bands in the lower MW region were smeared and less prominent compared to the other samples. This might be due to the higher chitin content of this sample: this polymer is tightly bound to insects cuticular proteins, which are present in this MW range, critically limiting their separation from chitin and thus interfering with their electrophoretic profile (Tetreau et al., 2015).
UPLC-ESI/MS analysis of amino acids
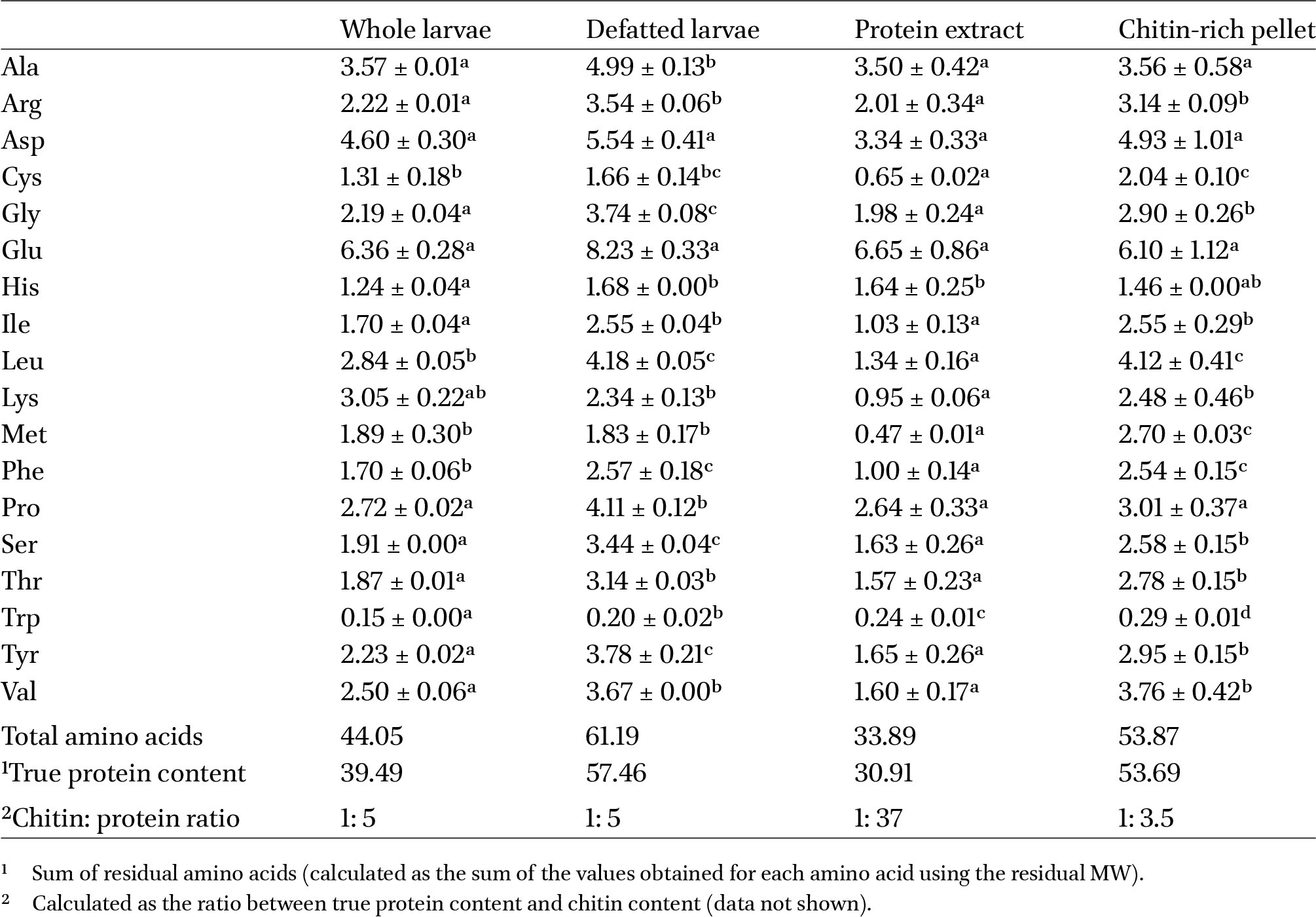
The analysis of amino acids was carried out on the whole larvae and on their defatted fraction, protein isolate, and chitin-rich fraction. The main objectives were to further investigate the possible influence of the defatting process on the protein fraction, as previously explored with the SDS-PAGE analysis, to determine the amino acidic distribution of the protein and chitin isolates, and to assess the protein quality of the four samples based on their amino acid profile. The results are shown in Table 3.
Amino acid profile of black soldier fly whole larvae, defatted larvae, protein isolate, and chitin-rich pellet. The results are expressed as g/100 g DM. Significant differences within each line are indicated by distinct letters (one-way ANOVA with post hoc Tukey b’s test,
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
By comparing the results with the literature, it can be observed that the amino acidic content of the whole larvae was overall in line with that of Caligiani et al. (2018) and Traksele et al. (2021), with few exceptions. Specifically, with respect to essential amino acids, the content of lysine and of sulphurated amino acids was higher in our sample, while the content of tryptophan was in line with Caligiani et al. (2018) and lower than the one reported in the study conducted by Traksele et al. (2021). Eggink and Dalsgaard (2023) and Spranghers et al. (2017) have also observed that this amino acid is one of the least abundant in Hermetia illucens. In addition, differences among the three sets of results were also observed for tyrosine and valine, of which Caligiani et al. (2018) reported higher values. These differences on the whole larvae makes clear that there is a certain variability in the amino acid composition, which may be attributed to the different feed substrates used for rearing the insects (Fuso et al., 2021; Leni et al., 2020). Black soldier fly can be reared on many food wastes and by-products, making it one of the most efficient in converting wastes into valuable biomass; however, the inhomogeneity and lack of standardization of the wastes binds their diet to the seasonal availability of the rearing substrate used as feed. As a matter of fact, significantly different protein contents can be observed within the same insect species according to the rearing substrate used, mainly due to the seasonality of the biomass employed (Fuso et al., 2021). Meanwhile, the amino acid profile of the defatted sample was comparable with the findings of Traksele et al. (2021). However, the current samples were high in sulphurated amino acids and low in tryptophan. Additionally, the overall trend of the amino acid profile is the same for the whole and the defatted sample, implying that the defatting process did not alter the composition of the protein pool, agreeing with what we already highlighted in section “Electrophoretic profile”.
The protein content of the protein isolate and of the chitin-rich fraction was 30.91 and 53.69% on a dry matter basis, respectively, indicating that a substantial fraction of proteins was retained by the chitin-rich pellet. As previously mentioned in section “Proximate composition of black soldier fly larvae and of its defatted and lipid fractions, protein isolate, and chitin-rich pellet”, chitin in its natural form is often tightly bound to proteins, thus hindering their detachment and complete extraction. This agreed with the gel electrophoresis profile, where the presence of chitin might have compromised the sharpness of the protein bands. By grouping the amino acids into categories based on their R group (aliphatic: glycine, alanine, proline, valine, isoleucine, leucine; aromatic: tryptophan, tyrosine, phenylalanine; acidic: aspartic acid, glutamic acid; hydroxylic: serine, threonine; sulphur-containing: cysteine, methionine; basic: histidine, arginine, lysine) it emerged that the protein isolate had slightly lower levels of aliphatic (−1%) and aromatic (−2%) amino acids and higher levels of acidic ones (+9%) compared to the chitin-rich pellet, which was in accordance with the results by Pedrazzani et al. (2024). This possibly means that the proteins of higher molecular weight contained in the chitin-rich fraction (and not in the protein isolate) might be richer in aliphatic and aromatic amino acids, while proteins of lower molecular weight in acidic ones.
Determination of the antibacterial activity
Several studies have been conducted recently to investigate the antibacterial potential of BSF larvae. Harlystiarini et al. (2019) observed antibacterial activity of BSF larvae methanolic extracts against gram-negative bacteria, specifically Salmonella sp. and E. coli. Auza et al. (2020) conducted a similar study and determined that the extracts inhibited the growth of Salmonella thypimurium, E. coli, and Pseudomonas aeruginosa. Dong et al. (2021) observed growth inhibition of Clostridium perfringens by culturing the bacteria with BSF digesta. Van Moll et al. (2022) positively evaluated the antibacterial effect of antimicrobial peptides extracted from BSF against a panel of gram-negative bacteria. However, relatively little to no research has been carried out on livestock pathogens using whole BSF larvae and their various fractions, thus emphasizing the importance of exploring this topic.
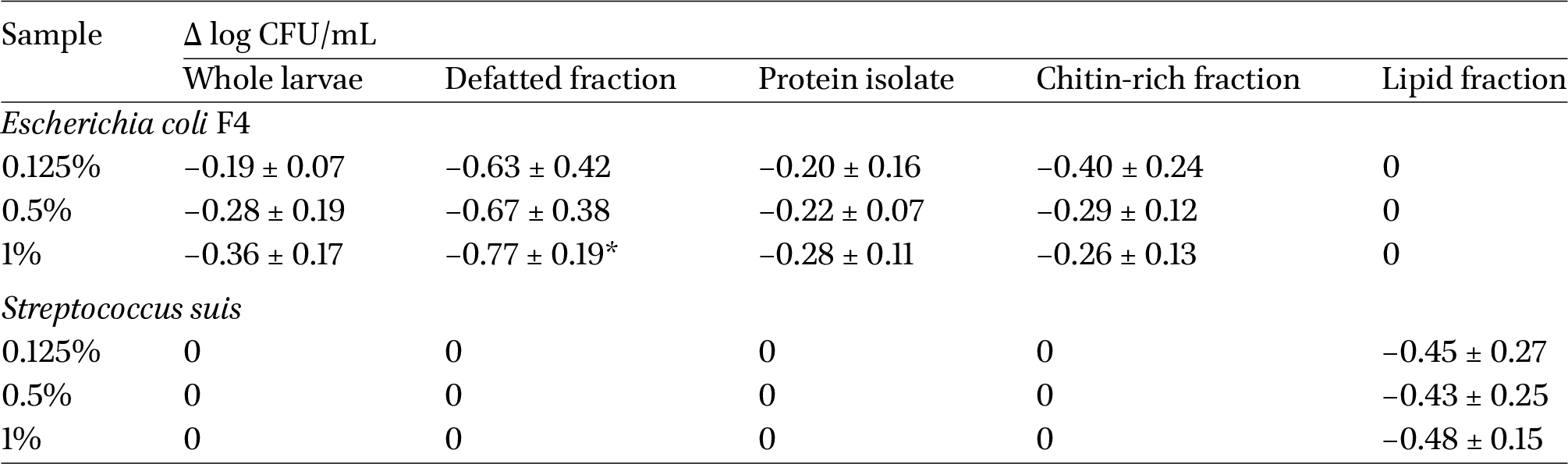
Therefore, antibacterial activity tests were carried out on the whole larvae and on their defatted fraction, protein isolate, chitin-rich fraction, and lipid fraction against E. coli F4 and S. suis, as described in section “Determination of the antibacterial activity”. The results, expressed as Δ log CFU/mL, are reported in Table 4.
Antibacterial activity of whole black soldier fly larvae, defatted larvae, and of their isolated protein, lipid, and chitin fractions against Escherichia coli F4 and Streptococcus suis. The results are expressed as Δ log CFU/mL (T3 − T0). Values are the mean of three biological replicates ± standard deviation. Significant differences within each line are indicated by an asterisk (Kruskal-Wallis non-parametric test,
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
The defatted fraction showed the highest efficacy against E. coli, resulting in a decrease of −0.63, −0.67, and −0.77 log CFU/mL in the 0.125, 0.5, and 1% samples, respectively, while the lipid fraction did not show any activity. In addition, when comparing the results of the protein isolate and the chitin-rich fraction, this last shows higher activity compared to the first. The protein isolate was expected to show the highest antibacterial potential in comparison to the other fractions because, as it is shown in the literature, antimicrobial peptides usually show higher activity against gram-negative bacteria (Auza et al., 2020; Harlystiarini et al., 2019; Van Moll et al., 2022). One hypothesis that might explain this behaviour could be a putative synergistic effect of the proteins and chitin contained in the defatted fraction: in fact, it can be observed that this sample, which had the highest protein content (57.46%) and second-highest chitin content (10.57%), showed the highest values of relative kill-off per percentage of sample added. In order to determine whether or not this difference was statistically significant, a Kruskal-Wallis non-parametric test was carried out for each sample concentration, and the results showed that the 1% defatted fraction was significantly different from all the other tested samples (
A completely different behaviour was observed for S. suis. The only fraction that exhibited antibacterial activity against this bacterium was the lipid fraction, showing the highest activity as at the highest sample concentration. This can be mainly attributed to the high concentration of lauric acid in the black soldier fly lipid fraction: in fact, the antibacterial and antiviral properties of medium chained fatty acids, and in particular of lauric acid, are widely known. Furthermore, studies showed that lauric acid was more effective on gram-positive bacteria, category that includes S. suis, than on gram-negative ones (Casillas-Vargas et al., 2021; Dong et al., 2021). Additionally, studies have shown that other fatty acids can also exhibit antibacterial activity, namely myristic acid (C14:0) and oleic acid (C18:1) (Veldkamp et al., 2022); as a matter of fact, both are present in considerable amounts in the analysed sample, constituting roughly 24% of the total fatty acids. Despite the whole larvae contained lipids, the sample did not show any antibacterial activity on S. suis: this could be due to the different percentages of lipids contained in the two samples. In fact, the whole larvae contained around 40% of lipids, while the tested lipid fraction was constituted only by fats, thus possibly meaning that a minimum amount of sample is needed in order to show efficacy. Additionally, the whole larvae also contained proteins and chitin, both of which could possibly constitute a source of carbon and nitrogen for bacterial growth, thus being in antagonism with the antimicrobial effect of the lipid fraction.
Overall, the results show that nitrogen-rich fractions were active against E. coli F4 and not against S. suis, a gram-negative and a gram-positive bacterium, respectively. In contrast, the lipid fraction was the only one active against S. suis. As the efficacy of antimicrobial peptides on gram-negative or gram-positive bacteria depends largely on the amino acid residues of which they are constituted, the results might mean that the proteins contained antimicrobial peptides rich in glycine residues, such as attacins, which are more effective on this class of bacteria (Veldkamp et al., 2022). However, in order to test this hypothesis and further study this phenomenon, an in-depth peptide characterization through high resolution mass spectrometry would be necessary to determine if and which antimicrobial peptides are present.
Determination of the prebiotic activity
In 2017, the International Scientific Association for Probiotics and Prebiotics (ISAPP) updated the definition of a prebiotic, defining it as “a substrate that is selectively utilised by host microorganisms conferring a health benefit” (Gibson et al., 2017). In recent years, one of the most studied alternatives to antibiotics in swine production is the use of in-feed prebiotics and probiotics. In fact, it has been widely demonstrated that these promote a healthy gut microbiota, which in turn improves feed conversion ratio, nutrient digestibility, and resistance to pathogenic infections (Dowarah et al., 2017; Hou et al., 2015; Yang et al., 2015). Chitin, a long-chain polymer of N-acetylglucosamine which constitutes the exoskeleton of insects, has recently gained attention due to its putative prebiotic effect on gut microorganisms: in fact, it can be a good source of dietary fibre in human and animal diets, thus promoting the growth of probiotic microorganisms in the intestines (Dowarah et al., 2017; Harkin et al., 2019; Hou et al., 2015; Liao and Nyachoti, 2017; van Huis, 2021). However, just as many studies appointed at chitin as antibacterial agent, rather than a prebiotic one, mainly depending on its degree of deacetylation (Benhabiles et al., 2012; Guarnieri et al., 2022; Srinivasan et al., 2023).
Therefore, the prebiotic activity of the whole larvae and of their protein isolate and chitin-rich fraction was assessed in order to determine which fraction was the most efficient in promoting the growth of probiotic bacteria. The tests were carried out with Limosilactobacillus reuteri, a common endogenous probiotic microorganism in pigs (Hou et al., 2015; Slizewska and Chlebicz-Wójcik, 2020), as described in section “Determination of the prebiotic activity”. The mean negative control for all samples and all trials was 4.5 ± 0.40 log10 CFU/mL at t = 0 hours, and 3.8 ± 0.34 log10 CFU/mL at t = 48 hours, meaning that, in absence of any source of carbon or nitrogen, the bacterial load decreased after 48 hours (Table 5).
Prebiotic activity of black soldier fly whole larvae and of their isolated protein and chitin fractions on Limosilactobacillus reuteri. The results are expressed as log CFU/mL. Samples were tested at a concentration of 42 g/L (100%) and 21 g/L (50%), which corresponded to the mass of N- and C-containing solutes missing from the nutrient-deprived MRS broth. Values are the mean of three biological replicates ± standard deviation. Significant differences within each column are indicated by distinct letters (one-way ANOVA with post hoc Tukey b’s test,
Citation: Journal of Insects as Food and Feed 11, 2 (2025) ; 10.1163/23524588-00001152
All samples showed prebiotic activity for L. reuteri, meaning that they served as substrate for the growth of the bacterium. The protein isolate was the most effective in promoting bacterial growth, with the final bacterial concentration of the sample at 100% at T48 9.4 log10 CFU/mL, even higher than the one of the positive control (9.2 log10 CFU/mL), followed by the whole larvae (9.2 log10 CFU/mL), and finally by the chitin-rich fraction (8.0 log10 CFU/mL). It is to be noted that the bacterial growth in chitin-rich pellet samples at both concentrations was always statistically lower than the one in the whole larvae and the protein isolate. Chitin is often described as a potential source of dietary fibre and a novel prebiotic agent in the literature (Acosta-Estrada et al., 2021; Ojha et al., 2021; Sogari et al., 2019). Indeed, a few studies investigated and confirmed the prebiotic effect of whole insects (de Carvalho et al., 2019; Stull et al., 2018); nevertheless, no studies were found on the potential prebiotic activity of the fractionated components of insects. In fact, as it can be seen from our results, the whole insect did exhibit prebiotic activity, as already shown in the literature, but this effect notably dropped with the increase of chitin concentration; specifically, the bacterial growth decreased with 1.2 log CFU/mL for the test with 100% of chitin-rich pellet and with 1.6 log CFU/mL for the one with 50% of sample. Therefore, based on these results, chitin might be considered more of an antimicrobial agent rather than a prebiotic one.
By relating the results with the protein and chitin content of the three tested samples (Table 1), it is noticeable that the chitin-rich pellet also contained proteins, and in higher amounts with respect to the isolated protein fraction: thus, we could infer that the nitrogen of the chitin-rich fraction, despite being more abundant, seemed to be less readily usable by bacteria than the nitrogen of the protein isolate alone. By also including the results on the amino acid content of the samples (Table 3) and on their molecular weight distribution (Figure 1), it is to be noted that the two fractions contained proteins of different molecular weights and had a slightly different amino acid composition. In fact, the protein isolate was made up of <25 kDa proteins and was richer in acidic amino acids (+9%), while the chitin-rich fraction contained proteins in the MW range 25-75 kDa and had slightly higher levels of aliphatic (+1%) and aromatic (+2%) amino acids. This difference leads to the hypothesis that the proteins bound to chitin are less reachable by bacterial enzymes, so less hydrolysable by bacterial proteases in comparison to the isolated proteins, implying that chitin might act like an antinutritional factor, making proteins both less extractable and less reachable by enzymes, hindering their breakdown.
4 Conclusions
At present, there is a lack of knowledge on the correlation between insect fractions and their potential antibacterial and prebiotic activity, and little to no research has been carried out on livestock pathogens. Therefore, this study aimed at assessing the antibacterial and prebiotic activity of whole BSF larvae and of their isolated defatted, protein, chitin-rich, and lipid fractions with livestock pathogens (Escherichia coli F4 and Streptococcus suis) and probiotic microorganisms (Limosilactobacillus reuteri). The analysis of amino acids showed that the protein content of the whole and defatted larvae was overall in accordance with previous studies. A high percentage of residual proteins was still present in the chitin-rich fraction, implying that chitin hindered protein extraction, in accordance with the SDS-PAGE analysis, where the protein isolate showed only bands at lower molecular weight compared to the chitin-rich fraction. The analysis of fatty acids showed that lauric acid was the most abundant, also in agreement with the literature. The antibacterial activity tests revealed that, in general, all nitrogen-containing samples showed activity against E. coli, while only the lipid fraction against S. suis. The results pointed out that chitin might potentially be an antibacterial agent, rather than a prebiotic one: this also agreed with the prebiotic activity tests, where the increase in chitin concentration resulted in the decrease of the prebiotic effect. In fact, the protein isolate showed the highest value, indicating that the nitrogen was readily available for the bacteria, while the bacterial growth was less favoured at higher chitin percentages. Overall, the results highlighted the potential use of BSF larvae and their fractions as a valuable source of proteins and lipids in animal feed. Our findings also emphasized the promising bioactivity of the analysed samples. Given the rise of antimicrobial resistance, the integration of BSF whole larvae or their fractions in animal feed could offer a novel approach to mitigate this global burden. Further studies are still required to fully elucidate chitin bioactivity.
Corresponding author; e-mail: mariagiulia.bonomini@unipr.it
Conflict of interest
The authors have no conflict of interest to declare.
References
Aarestrup, F.M., Oliver Duran, C. and Burch, D.G.S., 2008. Antimicrobial resistance in swine production. Animal Health Research Reviews 9: 135-148. https://doi.org/10.1017/S1466252308001503
Abubakar, R.H., Madoroba, E., Adebowale, O., Fasanmi, O.G. and Fasina, F.O., 2019. Antimicrobial usage in pig production: effects on Escherichia coli virulence profiles and antimicrobial resistance. Onderstepoort Journal of Veterinary Research 86: a1743. https://doi.org/10.4102/ojvr.v86i1.1743
Acosta-Estrada, B.A., Reyes, A., Rosell, C.M., Rodrigo, D. and Ibarra-Herrera, C.C., 2021. Benefits and challenges in the incorporation of insects in food products. Frontiers in Nutrition 8. https://doi.org/10.3389/fnut.2021.687712
Auza, F.A., Purwanti, S., Syamsu, J.A. and Natsir, A., 2020. Antibacterial activities of black soldier flies (Hermetia illucens L.) extract towards the growth of Salmonella typhimurium, E. coli and Pseudomonas aeruginosa. IOP Conference Series: Earth and Environmental Science 492: 012024. https://doi.org/10.1088/1755-1315/492/1/012024
Benhabiles, M.S., Salah, R., Lounici, H., Drouiche, N., Goosen, M.F.A. and Mameri, N., 2012. Antibacterial activity of chitin, chitosan and its oligomers prepared from shrimp shell waste. Food Hydrocolloids 29: 48-56. https://doi.org/10.1016/j.foodhyd.2012.02.013
Borrelli, L., Varriale, L., Dipineto, L., Pace, A., Menna, L.F. and Fioretti, A., 2021. Insect derived lauric acid as promising alternative strategy to antibiotics in the antimicrobial resistance scenario. Frontiers in Microbiology 12. https://doi.org/10.3389/fmicb.2021.620798
Caligiani, A., Marseglia, A., Sorci, A., Bonzanini, F., Lolli, V., Maistrello, L. and Sforza, S., 2019. Influence of the killing method of the black soldier fly on its lipid composition. Food Research International 116: 276-282. https://doi.org/10.1016/j.foodres.2018.08.033
Casillas-Vargas, G., Ocasio-Malavé, C., Medina, S., Morales-Guzmán, C., Del Valle, R.G., Carballeira, N.M. and Sanabria-Rı́os, D.J., 2021. Antibacterial fatty acids: an update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Progress in Lipid Research 82: 101093. https://doi.org/10.1016/j.plipres.2021.101093
Chen, C., Chen, L., Mao, C., Jin, L., Wu, S., Zheng, Y., Cui, Z., Li, Z., Zhang, Y., Zhu, S., Jiang, H. and Liu, X., 2023. Natural Extracts for Antibacterial Applications. Small 20: 2306553. https://doi.org/10.1002/smll.202306553
Choi, W.H., Yun, J.H., Chu, J.P. and Chu, K.B., 2012. Antibacterial effect of extracts of Hermetia illucens (Diptera: Stratiomyidae) larvae against gram-negative bacteria. Entomological Research 42: 219-226. https://doi.org/10.1111/j.1748-5967.2012.00465.x
Danieli, P.P., Lussiana, C., Gasco, L., Amici, A. and Ronchi, B., 2019. The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 9: 178. https://doi.org/10.3390/ani9040178
de Carvalho, N.M., Walton, G.E., Poveda, C.G., Silva, S.N., Amorim, M., Madureira, A.R., Pintado, M.E., Gibson, G.R. and Jauregi, P., 2019. Study of in vitro digestion of Tenebrio molitor flour for evaluation of its impact on the human gut microbiota. Journal of Functional Foods 59: 101-109. https://doi.org/10.1016/j.jff.2019.05.024
Dong, L., Ariëns, R.M.C., America, A.H.P., Paul, A., Veldkamp, T., Mes, J.J., Wichers, H.J. and Govers, C., 2021. Clostridium perfringens suppressing activity in black soldier fly protein preparations. LWT 149: 111806. https://doi.org/10.1016/j.lwt.2021.111806
Dowarah, R., Verma, A.K. and Agarwal, N., 2017. The use of Lactobacillus as an alternative of antibiotic growth promoters in pigs: A review. Animal Nutrition 3: 1-6. https://doi.org/10.1016/j.aninu.2016.11.002
Eggink, K.M. and Dalsgaard, J., 2023. Chitin contents in different black soldier fly (Hermetia illucens) life stages. Journal of Insects as Food and Feed 9: 855-863. https://doi.org/10.3920/JIFF2022.0142
European Commission (EC), (n.d.). Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Available at: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32009R0152
Ewald, N., Vidakovic, A., Langeland, M., Kiessling, A., Sampels, S. and Lalander, C., 2020. Fatty acid composition of black soldier fly larvae (Hermetia illucens) – possibilities and limitations for modification through diet. Waste Management 102: 40-47. https://doi.org/10.1016/j.wasman.2019.10.014
Fuso, A., Barbi, S., Macavei, L.I., Luparelli, A.V., Maistrello, L., Montorsi, M., Sforza, S. and Caligiani, A., 2021. Effect of the rearing substrate on total protein and amino acid composition in black soldier fly. Foods 10: 1773. https://doi.org/10.3390/foods10081773
Ghosh, A., Das, B.K., Roy, A., Mandal, B. and Chandra, G., 2008. Antibacterial activity of some medicinal plant extracts. Journal of Natural Medicines 62: 259-262. https://doi.org/10.1007/s11418-007-0216-x
Gibson, G.R., Hutkins, R., Sanders, M.E., Prescott, S.L., Reimer, R.A., Salminen, S.J., Scott, K., Stanton, C., Swanson, K.S., Cani, P.D., Verbeke, K. and Reid, G., 2017. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology & Hepatology 14: 491-502. https://doi.org/10.1038/nrgastro.2017.75
Guarnieri, A., Triunfo, M., Scieuzo, C., Ianniciello, D., Tafi, E., Hahn, T., Zibek, S., Salvia, R., De Bonis, A. and Falabella, P., 2022. Antimicrobial properties of chitosan from different developmental stages of the bioconverter insect Hermetia illucens. Scientific Reports 12: 8084. https://doi.org/10.1038/s41598-022-12150-3
Harkin, C., Mehlmer, N., Woortman, D.V., Brück, T.B. and Brück, W.M., 2019. Nutritional and Additive Uses of Chitin and Chitosan in the Food Industry. In: Crini, G. and Lichtfouse, E. (eds.) Sustainable Agriculture Reviews 36. Springer, Cham, Switzerland, pp. 1-43. https://doi.org/10.1007/978-3-030-16581-9_1
Harlystiarini, H., Mutia, R., Wibawan, I.W.T. and Astuti, D.A., 2019. In vitro antibacterial activity of black soldier fly (Hermetia Illucens) larva extracts against gram-negative bacteria. Buletin Peternakan 43. https://doi.org/10.21059/buletinpeternak.v43i2.42833
Hawkey, K.J., Lopez-Viso, C., Brameld, J.M., Parr, T. and Salter, A.M., 2021. Insects: a potential source of protein and other nutrients for feed and food. Annual Review of Animal Biosciences 9: 333-354. https://doi.org/10.1146/annurev-animal-021419-083930
Hou, C., Zeng, X., Yang, F., Liu, H. and Qiao, S., 2015. Study and use of the probiotic Lactobacillus reuteri in pigs: a review. Journal of Animal Science and Biotechnology 6: 14. https://doi.org/10.1186/s40104-015-0014-3
Hurtado-Ribeira, R., Silvan, J.M., Fornari, T., Vázquez, L., Martinez-Rodriguez, A.J. and Martin, D., 2023. Modulation of the lipolysis and subsequent antibacterial activity of the fat from black soldier fly (Hermetia illucens) by the combined selection of slaughtering, drying and defatting methods of the larvae. Innovative Food Science & Emerging Technologies 90: 103510. https://doi.org/10.1016/j.ifset.2023.103510
Jadeja, N.B. and Worrich, A., 2022. From gut to mud: dissemination of antimicrobial resistance between animal and agricultural niches. Environmental Microbiology 24: 3290-3306. https://doi.org/10.1111/1462-2920.15927
Koutsos, L., Mccomb, A. and Finke, M., 2019. Insect composition and uses in animal feeding applications: a brief review. Annals of the Entomological Society of America 112: 544-551. https://doi.org/10.1093/aesa/saz033
Leni, G., Caligiani, A. and Sforza, S., 2019. Killing method affects the browning and the quality of the protein fraction of Black Soldier Fly (Hermetia illucens) prepupae: a metabolomics and proteomic insight. Food Research International 115: 116-125. https://doi.org/10.1016/j.foodres.2018.08.021
Leni, G., Caligiani, A. and Sforza, S., 2021. Bioconversion of agri-food waste and by-products through insects: a new valorization opportunity. In: Rajeev Bhat, R. (ed.) Valorization of agri-food wastes and by-products. Academic Press, Cambridge, MA, USA, pp. 809-828. https://doi.org/10.1016/B978-0-12-824044-1.00013-1
Leni, G., Soetemans, L., Jacobs, J., Depraetere, S., Gianotten, N., Bastiaens, L., Caligiani, A. and Sforza, S., 2020. Protein hydrolysates from Alphitobius diaperinus and Hermetia illucens larvae treated with commercial proteases. Journal of Insects as Food and Feed 6: 393-404. https://doi.org/10.3920/JIFF2019.0037
Li, X., Dong, Y., Sun, Q., Tan, X., You, C., Huang, Y. and Zhou, M., 2022. Growth and fatty acid composition of black soldier gly Hermetia illucens (Diptera: Stratiomyidae) larvae are influenced by dietary fat sources and levels. Animals 12: 486. https://doi.org/10.3390/ani12040486
Liao, S.F. and Nyachoti, M., 2017. Using probiotics to improve swine gut health and nutrient utilization. Animal Nutrition 3: 331-343. https://doi.org/10.1016/j.aninu.2017.06.007
Lu, S., Taethaisong, N., Meethip, W., Surakhunthod, J., Sinpru, B., Sroichak, T., Archa, P., Thongpea, S., Paengkoum, S., Purba, R.A.P. and Paengkoum, P., 2022. Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: a review. Insects 13: 831. https://doi.org/10.3390/insects13090831
Luparelli, A., Leni, G., Fuso, A., Pedrazzani, C., Palini, S., Sforza, S. and Caligiani, A., 2023. Development of a quantitative UPLC-ESI/MS method for the simultaneous determination of the chitin and protein content in insects. Food Analytical Methods 16: 252-265. https://doi.org/10.1007/s12161-022-02411-2
Matsue, M., Mori, Y., Nagase, S., Sugiyama, Y., Hirano, R., Ogai, K., Ogura, K., Kurihara, S. and Okamoto, S., 2019. Measuring the antimicrobial activity of lauric acid against various Bacteria in human gut microbiota using a new method. Cell Transplantation 28: 1528-1541. https://doi.org/10.1177/0963689719881366
Murray, C.J., Ikuta, K.S., Sharara, F., Swetschinski, L., Robles Aguilar, G., Gray, A., Han, C., Bisignano, C., Rao, P., Wool, E., Johnson, S.C., Browne, A.J., Chipeta, M.G., Fell, F., Hackett, S., Haines-Woodhouse, G., Kashef Hamadani, B.H., Kumaran, E.A.P., McManigal, B., Naghavi, M., … 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 399: 629-655. https://doi.org/10.1016/S0140-6736(21)02724-0
Obradovic, M.R., Segura, M., Segalés, J. and Gottschalk, M., 2021. Review of the speculative role of co-infections in Streptococcus suis-associated diseases in pigs. Veterinary Research 52: 49. https://doi.org/10.1186/s13567-021-00918-w
Ojha, S., Bekhit, A.E.-D., Grune, T. and Schlüter, O.K., 2021. Bioavailability of nutrients from edible insects. Current Opinion in Food Science 41: 240-248. https://doi.org/10.1016/j.cofs.2021.08.003
Pam Ismail, B., Senaratne-Lenagala, L., Stube, A. and Brackenridge, A., 2020. Protein demand: review of plant and animal proteins used in alternative protein product development and production. Animal Frontiers 10: 53-63. https://doi.org/10.1093/af/vfaa040
Pasini, G., Cullere, M., Vegro, M., Simonato, B. and Dalle Zotte, A., 2022. Potentiality of protein fractions from the house cricket (Acheta domesticus) and yellow mealworm (Tenebrio molitor) for pasta formulation. LWT 164: 113638. https://doi.org/10.1016/j.lwt.2022.113638
Pedrazzani, C., Righi, L., Vescovi, F., Maistrello, L. and Caligiani, A., 2024. Black soldier fly as a new chitin source: extraction, purification and molecular/structural characterization. LWT 191: 115618. https://doi.org/10.1016/j.lwt.2023.115618
Prandi, B., Zurlini, C., Maria, C.I., Cutroneo, S., Di Massimo, M., Bondi, M., Brutti, A., Sforza, S. and Tedeschi, T., 2021. Targeting the nutritional value of proteins from legumes by-products through mild extraction technologies. Frontiers in Nutrition 8. https://doi.org/10.3389/fnut.2021.695793
Queiroz, L.S., Regnard, M., Jessen, F., Mohammadifar, M.A., Sloth, J.J., Petersen, H.O., Ajalloueian, F., Brouzes, C.M.C., Fraihi, W., Fallquist, H., de Carvalho, A.F. and Casanova, F., 2021. Physico-chemical and colloidal properties of protein extracted from black soldier fly (Hermetia illucens) larvae. International Journal of Biological Macromolecules 186: 714-723. https://doi.org/10.1016/j.ijbiomac.2021.07.081
Rabani, V., Cheatsazan, H. and Davani, S., 2019. Proteomics and lipidomics of black soldier fly (Diptera: Stratiomyidae) and blow fly (Diptera: Calliphoridae) larvae. Journal of Insect Science 19: 29. https://doi.org/10.1093/jisesa/iez050
Rangel, F., Monteiro, M., Santos, R.A., Ferreira-Martins, D., Cortinhas, R., Gasco, L., Gai, F., Pousão-Ferreira, P., Couto, A., Oliva-Teles, A., Serra, C.R. and Enes, P., 2024. Novel chitinolytic Bacillus spp. increase feed efficiency, feed digestibility, and survivability to Vibrio anguillarum in European seabass fed with diets containing Hermetia illucens larvae meal. Aquaculture 579: 740258. https://doi.org/10.1016/j.aquaculture.2023.740258
Refael, G., Riess, H.T., Levi, C.S., Magzal, F., Tamir, S., Koren, O. and Lesmes, U., 2022. Responses of the human gut microbiota to physiologically digested insect powders or isolated chitin thereof. Future Foods 6: 100197. https://doi.org/10.1016/j.fufo.2022.100197
Siddiqui, S.A., Snoeck, E.R., Tello, A., Alles, M.C., Fernando, I., Saraswati, Y.R., Rahayu, T., Grover, R., Ullah, M.I., Ristow, B. and Nagdalian, A.A., 2022. Manipulation of the black soldier fly larvae (Hermetia illucens; Diptera: Stratiomyidae) fatty acid profile through the substrate. Journal of Insects as Food and Feed 8: 837-855. https://doi.org/10.3920/JIFF2021.0162
Slizewska, K. and Chlebicz-Wójcik, A., 2020. The in vitro analysis of prebiotics to be used as a component of a synbiotic preparation. Nutrients 12: 1272. https://doi.org/10.3390/nu12051272
Sogari, G., Amato, M., Biasato, I., Chiesa, S. and Gasco, L., 2019. The potential role of insects as feed: a multi-perspective review. Animals 9: 119. https://doi.org/10.3390/ani9040119
Spranghers, T., Michiels, J., Vrancx, J., Ovyn, A., Eeckhout, M., De Clercq, P. and De Smet, S., 2018. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Animal Feed Science and Technology 235: 33-42. https://doi.org/10.1016/j.anifeedsci.2017.08.012
Spranghers, T., Ottoboni, M., Klootwijk, C., Ovyn, A., Deboosere, S., De Meulenaer, B., Michiels, J., Eeckhout, M., De Clercq, P. and De Smet, S., 2017. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. Journal of the Science of Food and Agriculture 97: 2594-2600. https://doi.org/10.1002/jsfa.8081
Srinivasan, S., Ahmed, N.S., Ashokkumar, N., Selvaraj, P., Vinothkumar, V., Pugalendhi, P., Saravanan, R., Raajasubramanian, D., Karunanidhi, M., Williams, P.G. and Murali, R., 2023. Antibacterial efficacy of natural compounds chitin and chitosan: a mechanistic disclosure. In: Sreekala, M.S., Ravindran, L., Goda, K. and Thomas, S. (eds.) Handbook of natural polymers, volume 1. Elsevier, Amsterdam, the Netherlands, pp. 177-201. https://doi.org/10.1016/B978-0-323-99853-6.00011-5
Stull, V.J., Finer, E., Bergmans, R.S., Febvre, H.P., Longhurst, C., Manter, D.K., Patz, J.A. and Weir, T.L., 2018. Impact of edible cricket consumption on gut microbiota in healthy adults, a double-blind, randomized crossover trial. Scientific Reports 8: 10762. https://doi.org/10.1038/s41598-018-29032-2
Suryati, T., Julaeha, E., Farabi, K., Ambarsari, H. and Hidayat, A.T., 2023. Lauric acid from the black soldier fly (Hermetia illucens) and its potential applications. Sustainability 15: 10383. https://doi.org/10.3390/su151310383
Taylor, P.W., 2013. Alternative natural sources for a new generation of antibacterial agents. International Journal of Antimicrobial Agents 42: 195-201. https://doi.org/10.1016/j.ijantimicag.2013.05.004
Tetreau, G., Dittmer, N.T., Cao, X., Agrawal, S., Chen, Y.R., Muthukrishnan, S., Haobo, J., Blissard, G.W., Kanost, M.R. and Wang, P., 2015. Analysis of chitin-binding proteins from Manduca sexta provides new insights into evolution of peritrophin A-type chitin-binding domains in insects. Insect Biochemistry and Molecular Biology 62: 127-141. https://doi.org/10.1016/j.ibmb.2014.12.002
van Huis, A., 2021. Prospects of insects as food and feed. Organic Agriculture 11: 301-308. https://doi.org/10.1007/s13165-020-00290-7
van Huis, A. and Gasco, L., 2023. Insects as feed for livestock production. Science 379: 138-139. https://doi.org/10.1126/science.adc9165
Van Moll, L., De Smet, J., Paas, A., Tegtmeier, D., Vilcinskas, A., Cos, P. and Van Campenhout, L., 2022. In vitro evaluation of antimicrobial peptides from the black soldier fly (Hermetia illucens) against a selection of human pathogens. Antimicrobial Chemotherapy 10: e01664-21. https://doi.org/10.1128/spectrum.01664-21
Vanqa, N., Mshayisa, V.V. and Basitere, M., 2022. Proximate, physicochemical, techno-functional and antioxidant properties of three edible insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) flours. Foods 11: 976. https://doi.org/10.3390/foods11070976
Veldkamp, T., Dong, L., Paul, A. and Govers, C., 2022. Bioactive properties of insect products for monogastric animals – a review. Journal of Insects as Food and Feed 8: 1027-1040. https://doi.org/10.3920/JIFF2021.0031
World Health Organization, 2017. Antimicrobial Resistance Fact sheets on sustainable development goals: health targets. World Health Organization, Regional Office for Europe, Copenhagen Ø, Denmark. Available at: https://iris.who.int/bitstream/handle/10665/340814/WHO-EURO-2017-2375-42130-58025-eng.pdf?sequence=3&isAllowed=y
Xia, J., Ge, C. and Yao, H., 2021. Antimicrobial peptides from black soldier fly (Hermetia illucens) as potential antimicrobial factors representing an alternative to antibiotics in livestock farming. Animals 11: 1937. https://doi.org/10.3390/ani11071937
Xu, J.H., Xiao, S., Wang, J.H., Wang, B., Cai, Y.X. and Hu, W.F., 2023. Comparative study of the effects of ultrasound-assisted alkaline extraction on black soldier fly (Hermetia illucens) larvae protein: nutritional, structural, and functional properties. Ultrasonics Sonochemistry 101: 106662. https://doi.org/10.1016/j.ultsonch.2023.106662
Yang, F., Hou, C., Zeng, X. and Qiao, S., 2015. The use of lactic acid bacteria as a probiotic in swine diets. Pathogens 4: 34-45. https://doi.org/10.3390/pathogens4010034
Yi, L., Lakemond, C.M.M., Sagis, L.M.C., Eisner-Schadler, V., van Huis, A. and Boekel, M.A.J.S.V., 2013. Extraction and characterisation of protein fractions from five insect species. Food Chemistry 141: 3341-3348. https://doi.org/10.1016/j.foodchem.2013.05.115
Zabulionė, A., Šalaševičienė, A., Makštutienė, N. and Šarkinas, A., 2023. Exploring the antimicrobial potential and stability of black soldier fly (Hermentia illucens) larvae fat for enhanced food shelf-life. Gels 9: 793. https://doi.org/10.3390/gels9100793
Zhou, Y., Wang, D., Zhou, S., Duan, H., Guo, J. and Yan, W., 2022. Nutritional composition, health benefits, and application value of edible insects: a review. Foods 11: 3961. https://doi.org/10.3390/foods11243961
Zhu, Y., Huang, W.E. and Yang, Q., 2022. Clinical perspective of antimicrobial resistance in bacteria. Infection and Drug Resistance 15: 735-746. https://doi.org/10.2147/IDR.S345574



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}