Abstract
Camouflage and crypsis are key survival strategies for many animals, enabling them to evade predators or enhance their ability to ambush prey. For cryptic species, effective camouflage requires not only morphological adaptations but also behavioral tactics that ensure individuals select appropriate microhabitats. However, how polymorphic cryptic species match their phenotype to their environment remains poorly understood. In this study, we investigated the morphological and behavioral plasticity of the polymorphic mantis Sphodromantis viridis from Mediterranean and desert populations to determine whether habitat selection and coloration occur when nymphs are reared in the absence of environmental color. We reared naïve individuals in color-neutral conditions and examined their morphology, development time, body length, and microhabitat preferences. Color in S. viridis is sex-linked, with males exhibiting higher proportions of brown coloration than females, and color change occurs unidirectionally from green to brown. Desert males displayed greater variance in body size and development time, forming two distinct morphs: one is smaller, greener, and matures before females, and another is larger, browner, and matures after females, suggesting alternative reproductive strategies. Neither nymphal nor adult coloration of naïve mantids influenced color choice in controlled conditions. Our findings suggest that S. viridis nymphs showed ontogenetic color change in the absence of environmental cues, which may reflect evolutionary adaptations to seasonal changes: green in spring when the vegetation is still green and brown later on when annuals dry out. Further studies on background-dependent rearing conditions could clarify how experience influences microhabitat choice.
Introduction
Animals evolve camouflage or crypsis for two main reasons: either to avoid predation or to gain an advantage in hunting prey by being undetected until they attack (Bond, 2007; Forsman et al., 2011; Pembury Smith and Ruxton, 2020; Dubiner et al., 2023). Species across myriad taxa, ranging from Uroplatus geckos with skin that mimics lichen-coated bark to Phyllium leaf insects with veined wings that replicate the minute details of leaves, have developed intricate and complex mimicry to match their particular microhabitat (Raxworthy et al., 2008; Bank et al., 2021). For these morphological adaptations to be advantageous, the animal must choose suitable backgrounds and microhabitats. In other words, if cryptic animals were to ambush prey or hide from predators in microhabitats that do not match their camouflage, their adaptation would be irrelevant (Stevens and Ruxton, 2019). Cryptic species often choose the appropriate microhabitat, or are at least attracted to the correct place, to camouflage effectively. Alternatively, individuals may develop an imprinted recognition of a habitat and maintain a preference for that specific substrate throughout life. They may also change their color with development to match the surroundings. Whatever the mechanism is, it requires having some perception of their own coloration and that of their surroundings.
Furthermore, as many cryptic species are polymorphic (Bond, 2007; Tsurui et al., 2010; Duarte et al., 2016), individuals must somehow take into account their specific morphology, which may differ from that of conspecifics and match it to the appropriate environment. The Thomisus crab spider, for example, has yellow, white, and pink morphs. While a yellow morph crab spider could potentially survive finding itself on a white flower by gradually transforming into a white morph over several days or weeks due to its polymorphic plasticity, it would stand out until then, putting it at a higher risk of predation and decreasing its ability to ambush prey (Lo et al., 2023; Théry et al., 2004). How cryptic animals choose matching habitats, and the mechanisms by which a polymorphic cryptic animal selects the microhabitat that best matches its phenotype, are poorly understood.
Two recent studies, on the caterpillars of a polymorphic species of peppered moth, Biston betularia, and a species of polymorphic mantid, demonstrated that individuals reared on green or brown substrates will gradually develop matching green or brown coloration (Eacock et al., 2019; Burke and Holwell, 2023). Eacock et al. (2019) observed that those same green or brown caterpillars, when subsequently given the choice, preferentially move to matching substrates instead of mismatched alternative substrates. Furthermore, Eacock et al. (2019) found that adaptive color change and microhabitat choice can be mediated by extraocular photoreception. When the caterpillars’ eyes are obscured and they are presented with a color choice, they consistently move to the same color; nevertheless, this means that color perception and decision-making are mediated by “light-sensing tissues distinct from the eyes,” and do not require vision.
Burke and Holwell (2023) found that while humidity and temperature played a role in color development in the springbok mantis Miomantis caffra, the strongest factor in color development was the microhabitat’s background. The role of background in the color development of polymorphic cryptic insects is consistent with another study, documenting mantids turning from brown to green corresponding to seasonal changes from summer to autumn (Battiston and Fontana, 2010), and studies documenting the rapid evolution of fire melanism in grasshoppers, whereby black or melanistic animals proliferate following a fire (Forsman et al., 2011; de Alcantra Viana et al., 2024) although this could be a product of natural selection as well. Furthermore, Llandres et al. (2013) determined that background coloration is a major factor responsible for phenotypic change in crab spiders and even identified hormones that induce this change, illustrating how background color triggers hormonal responses that bring about phenotype shifts.
Our goal was to shed light on several morphological and behavioral questions, using Mediterranean and desert populations of a polymorphic mantis (Sphodromantis viridis). Only a handful of studies have compared natural-history aspects of Mediterranean and desert populations of the same species (Volis et al., 2002; Scharf et al., 2008, 2009; Cohen et al., 2022; Schwarz et al., 2023). We sought to determine to what extent morphology is variable with ontogeny, between sexes, within populations, and between populations. We also examined if habitat selection depends on the individual’s color relative to the surrounding microhabitat by checking if “naive” mantids reared in a color-neutral environment still exhibit a preference for their own color. Given that feeding, temperature, and humidity are kept constant, and that mantids are not exposed to microhabitats of any natural color (but rather white), we examined whether habitat-of-origin (Mediterranean or desert) and sex impacted color, body size, and development time, or if all of these factors are plastic and mostly environmentally controlled. We also explored whether S. viridis preferentially rest on substrates where their individual phenotypes are better camouflaged (i.e., green mantids on green substrates and brown ones on brown substrates), after being raised in a “neutral” environment. Lastly, we examined whether color changes followed a directional trend and whether individuals consequently adjusted their microhabitat choices.
We expected microhabitat selection to be influenced by the mantids’ color, enabling them to adapt to rapidly changing habitats, following studies showing that mantids and lepidopteran larvae both react to background color and change color to match their background (Eacock et al., 2019; Burke and Holwell, 2023). We also expected the Mediterranean population to be larger and greener than the desert population. This is in accordance with the temperature-size rule and previous studies on insects in such areas (Kingsolver and Huey, 2008; Scharf et al., 2008), as well as the assumption that vegetation remains green longer in Mediterranean regions than in deserts.
Materials and Methods
We raised 61 naïve S. viridis (Fig. 1a) in color-neutral conditions (i.e., a white background), not allowing for any acquired preference for specific colors, to test innate color preference unrelated to growth conditions. Individuals were raised from oothecae and consisted of stock from two localities with significantly differing climates: We raised 31 individuals from the stock originating from a humid, coastal region near Tel Aviv (32 °06’49.2” N 34 °48’30.8” E), hereafter “Mediterranean population,” and 30 individuals from a stock originating from the hyper-arid desert region near Eilat (29 °30’03.6” N 34 °54’55.8” E), hereafter “desert population.” The two sites greatly differ in climate. For instance, the average yearly precipitation in Tel Aviv and Eilat is 524 mm and 32 mm, respectively, and the average number of hours per day with temperatures above 28 °C in July–August is 0 and 28, respectively (Goldreich, 2003). This should lead to greener vegetation in Tel Aviv and browner-yellower vegetation in Eilat. All mantids in each population stemmed from a single ootheca. The mantids were raised in clear, transparent containers (14 × 12 cm) with a mesh lid and on a white background, where humidity and light were kept constant: ~55% relative humidity, 25 °C, and a 12-12 L-D cycle. Relative humidity was kept as constant as possible via a water reservoir placed under container racks, and by daily misting. Individuals were fed two crickets (Acheta domesticus) twice a week, increasing in size following the mantids’ growth cycle, and with molting or freshly molted individuals temporarily excluded from feeding. We had 93.6% (29 of 31) and 90% (27 of 30) survival rates to adulthood in the Mediterranean and desert populations, respectively.

(a) A green female (left) and a brown male (right) adult mantis individuals of the studied species Sphodromantis viridis. (b) Bristol papers that were used in the color choice experiment.
Citation: Israel Journal of Ecology and Evolution 71, 4 (2025) ; 10.1163/22244662-bja10115
Behavioral tests
At the sub-adult stage, when individual color was noticeable but before wing development, each mantid was placed in a 30 × 30 cm mesh enclosure for 30 minutes and given a choice between two equidistant habitat options: green and brown (Fig. 1b). The enclosure was evenly lit from overhead by the same lights used during rearing and placed in the same climate-controlled incubator. One side was covered in brown paper and the other in green paper, both made of the same Bristol Paper. The direction of each habitat option (left or right) was randomly assigned. At the start of each trial, the mantid was placed at the neutral center of the enclosure, facing forward and equally exposed to both colors. Its choice, defined by the color it rested on at the end of the 30-minute period, was recorded, along with its phenotype (green or brown, which is a clear distinction). The trial was repeated seven days after adulthood was reached to assess fidelity in individual habitat preference between developmental stages. We conducted only two microhabitat tests, in the subadult and adult stages, to avoid any learning processes that might have induced bias towards one of the two microhabitats.
In addition to these trials, we recorded each mantid’s color at subadult and adult stages, as well as any color change, date of maturation, and adult body length. Maturation date (molting into winged adults) and adult length were used to compare developmental differences between the two populations. The color measures aimed to determine whether habitat preference is consistent over time and influenced by life stage or sex.
Statistical analysis
We conducted the following statistical tests: (1) We examined whether population (Mediterranean/desert) and sex affected development times using a two-way ANOVA (n = 55). Next, we examined whether population, sex, and development time affected body size using a two-way ANCOVA (n = 55). In both cases, we began with a saturated model and gradually removed non-significant interactions. We verified that the test residuals did not deviate from a normal distribution. We noticed that variances in body size and development time might differ among groups. To test it, we separately coded each population and sex (4 levels) and used two Bartlett’s tests with body size and development time as response variables. (2) Nymphal color was examined using logistic regression with population and sex as explanatory variables (n = 61). We then examined whether population, sex, development time, and their two-way interactions affected adult color using a logistic regression (n = 55). (3) To examine the effect of nymphal and adult color on microhabitat color choice (green/brown), we used two
Results
Development time: Neither population nor sex affected development time (F1,52 = 0.079, P = 0.780 and F1,52 = 1.370, P = 0.247, respectively). The two-way interaction term was not significant as well (F1,51 = 0.001, P = 0.975).
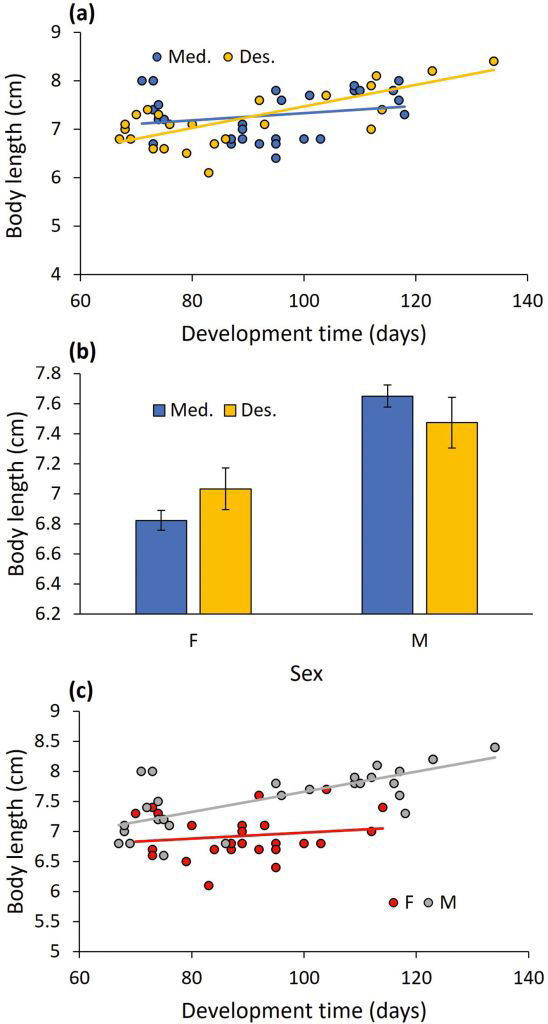
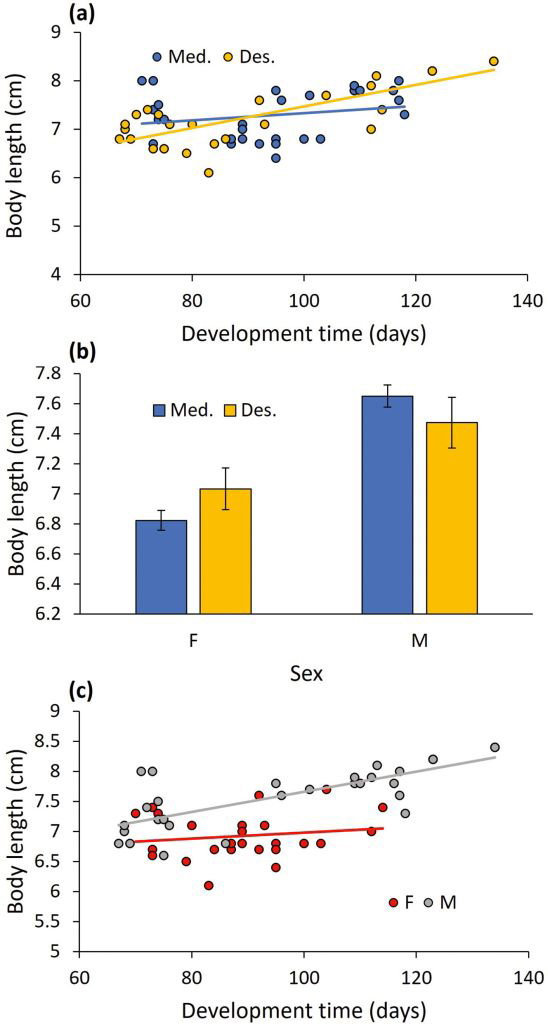
Body size: All two-way interactions were significant. Population interacted with development time to affect body size (F1,48 = 16.736, P < 0.001; Fig. 2a). The positive link between development time and body size was steeper for the desert population than for the Mediterranean population. Population interacted with sex to affect body size (F1,48 = 7.796, P = 0.008; Fig. 2b). Although females were generally shorter than males, the difference in body length between the sexes was greater in the Mediterranean population than in the desert population. The development time x sex interaction was significant too (F1,48 = 4.750, P = 0.034; Fig. 2c), indicating that the increase in body size with development time was steeper for males than females. To complete the analysis, population and development time were significant (F1,48 = 15.491, P < 0.001, and F1,48 = 6.012, P = 0.018, respectively), whereas sex was not (F1,48 = 1.192, P = 0.280).
The interactive effect of (a) development time and population (Mediterranean vs. Desert), (b) sex and population (means ± 1 SE), and (c) development time and sex on the adult body length.
Citation: Israel Journal of Ecology and Evolution 71, 4 (2025) ; 10.1163/22244662-bja10115
Variance in development time and body size: Males of the desert population exhibited the highest variance in development time, followed by males of the Mediterranean population, and then by the two female populations (
The effect of population (Mediterranean vs. Desert) and sex on (a) development time, and (b) body length. Boxplots (box: interquartile range, horizontal line inside the box: median, ×: mean, whiskers: all data without outliers).
Citation: Israel Journal of Ecology and Evolution 71, 4 (2025) ; 10.1163/22244662-bja10115
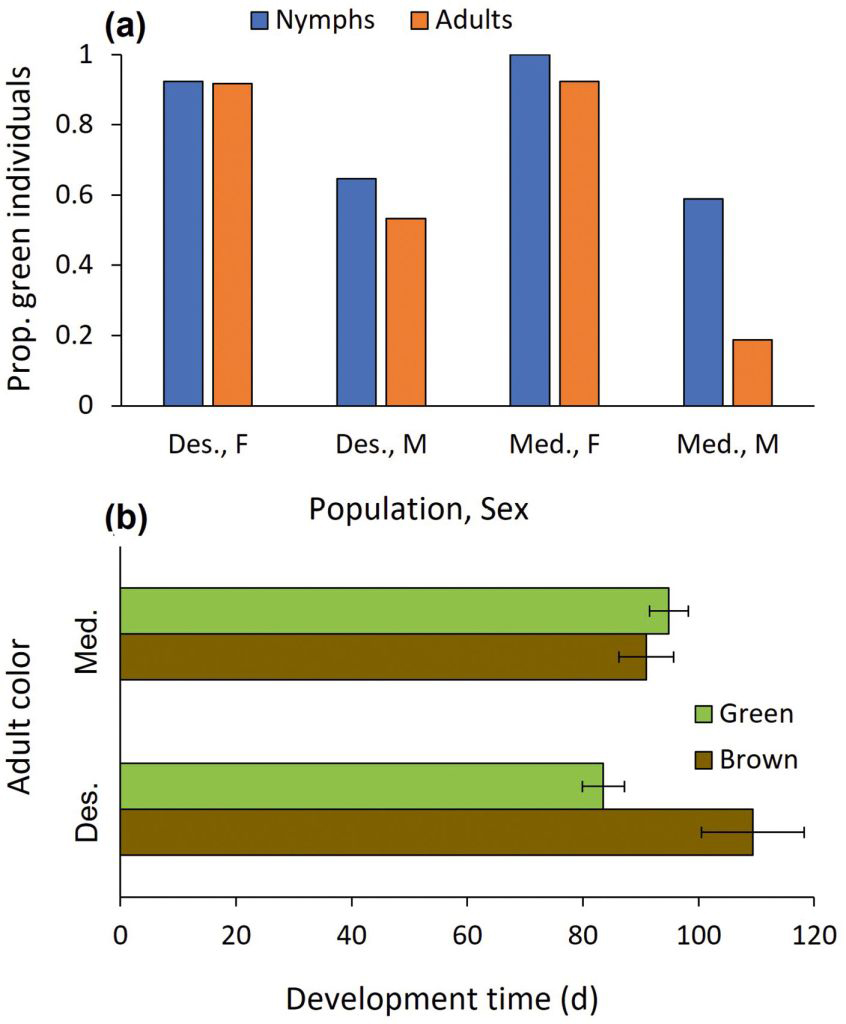
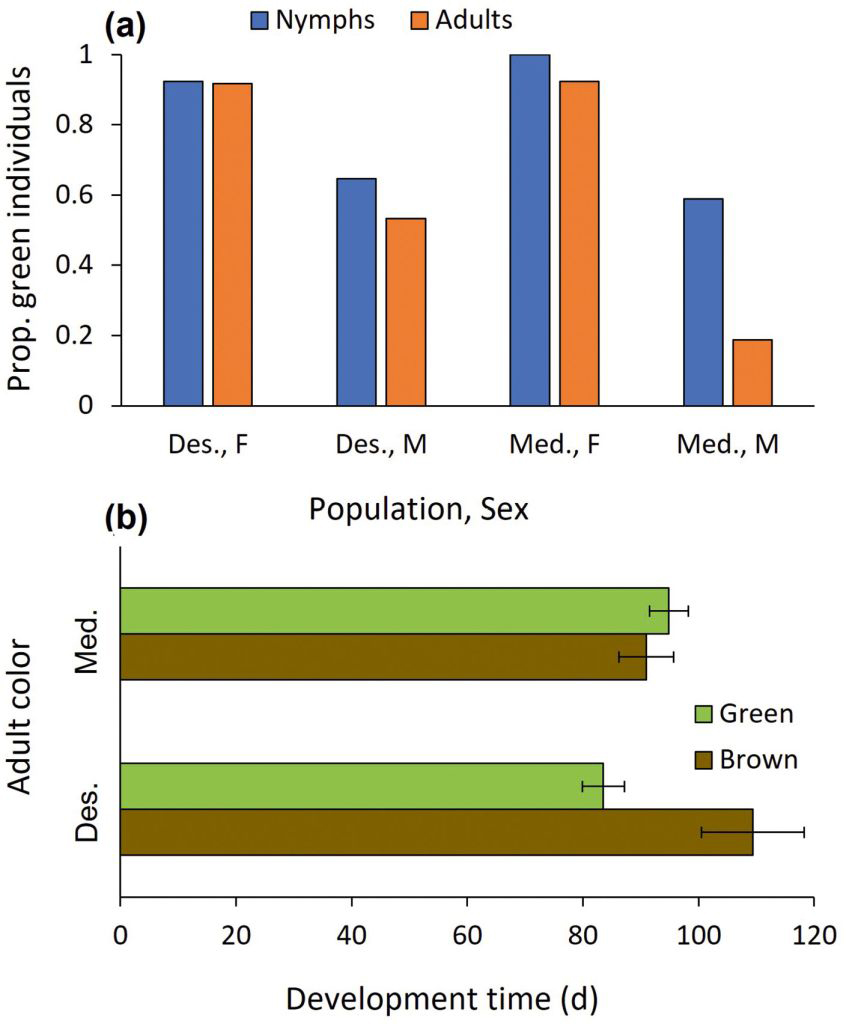
Nymphal and adult color: The nymphal color was affected by sex (z = -2.576, P = 0.010; Fig. 4a) but not by population (z = 0.012, P = 0.990). Regarding the adult color, population and development time interacted to affect color (z = 2.521, P = 0.012; Fig. 4b). Whereas development time did not affect Mediterranean individuals’ adult color, long development in the desert population was linked with a brown color morph, and short development characterized the green color morph. To complete the analysis, sex and population were significant as main effects (z = -3.584, P < 0.001 and z = -2.655, P = 0.008), but development time was not (z = -1.658, P = 0.097). While only 8% of the adult females were brown, 64.5% of the adult males were. Eight mantids changed color from green to brown, while none changed color from brown to green. Seven of these eight green-to-brown change events belonged to the Mediterranean population.
(a) The effect of population (Mediterranean vs. Desert) and sex on the proportion of green individuals at the nymphal and adult stages, and (b) of population and development time on adult color (means ± 1 SE).
Citation: Israel Journal of Ecology and Evolution 71, 4 (2025) ; 10.1163/22244662-bja10115
Color choice: Nymphal color did not affect color choice (
Discussion
Our study explored color change with ontogeny, body size, development time, and the microhabitat’s color choice of the polymorphic mantis S. viridis. We found that color in S. viridis was sex-linked, its change was unidirectional from green to brown, and that it, along with length, varied between populations even when environmental conditions were controlled for. Color choice, however, was unaffected by the nymphal or adult color when individuals were raised in controlled neutral conditions. Finally, the variance in development time and body size was greater in the desert population, whereby the males appeared to split into two sub-groups: one was smaller, greener, and matured before females of the same brood, while the other was larger, primarily comprised of brown individuals, and matured after the females. To the best of our knowledge, this phenomenon has not been observed in mantids previously. That said, our results and conclusions are only preliminary, as we had only one ootheca for each of the two populations. Future studies should examine whether the patterns detected here are general and hold across the steep climatic gradient of Israel.
Because the sexes differed in colors even in the absence of any background color during growth, we suggest that while numerous studies show the impact of the environment on color development (Eacock et al., 2019; Burke and Howell, 2023), genetics has a strong baseline influence as well. Color change in S. viridis was unidirectional: green individuals sometimes became brown, but brown individuals never became green. S. viridis has an annual lifecycle, beginning in early spring when vegetation in Israel is green, and reaching maturity in the summer or autumn when some vegetation has already dried and become brown. Accordingly, a staggered transformation to brown among part of a population may allow for gradual colonization of new brown microhabitats as they become available via seasonal drying. The turn from green when young to brown when adult matches the findings of at least two other studies on mantis species (Hierodula patellifera and Mantis religiosa; Okay, 1953; Wang et al., 2017). In both cases, as well as in our study, the majority of individuals were green and not brown.
Increased melanization in summer occurs in other insects too, for the sake of better camouflage (Haque et al., 2024). Alternatively, insects may be brighter in summer or warmer temperatures to avoid overheating (Trullas et al., 2007; Zeuss et al., 2014). Supporting this suggestion, a previous study on mantids found brown morphs to be around 1.5 °C warmer than green morphs (Köhler and Schielzeth, 2020). Regardless, unidirectional change from green to brown differs from the findings of Battiston and Fontana (2010), who recorded that polymorphic Mantis religiosa in Vicenza, Italy, is generally brown in the summer and then turns green in the fall, perhaps to match the influx of green vegetation due to autumn rains in the region. Autumn rains and the subsequent greening of vegetation begin significantly earlier in Vicenza than in central or southern Israel. This climatic difference puts selection pressure on Italian mantids to turn green later in life, while conferring no such advantage to their Israeli counterparts.
Unlike the findings of Battiston and Fontana (2010), however, we found that color-morph, at least in S. viridis, is sex-linked. Our data showed that sex is a strong predictor of adult coloration, with males exhibiting significantly more brown coloration than females in all groups (similar to Okay, 1953). More specifically, in adults, females from both populations were primarily green, whereas males were more varied. While the Mediterranean population exhibited significantly more brown males than the desert population, many of the Mediterranean males only turned brown upon their molt to adulthood.
Rosenheim et al. (2025) found a different but related phenomenon in the polymorphic mantis Stagmomantis limbata, whereby adult males were heterogeneous in color with both green and brown components, while females were homogenous in color, ranging from green to brown. Rosemheim et al. hypothesized that males possess more general coloration (i.e., a color that matches sufficiently well to a variety of backgrounds) due to their increased mobility, with the more sedentary females having camouflage specialized to their immediate area. Perhaps a similar mechanism also holds in the studied S. viridis: while green better fits stationary females camouflaging against live plants, shades of brown may be more useful for active males that are walking across dry leaves, ground, and rocks searching for females. Alternatively, greater variation in coloration may be helpful in disrupting predator learning and search image formation (Edmunds, 1990; Théry and Gomez, 2010), something that would be more relevant to active and exposed males than to females.
Unexpectedly, we found that while Mediterranean males fell into a single-size group with low variance and relatively uniform distribution, desert males exhibited a higher variance and showed two distinct length clusters, suggesting the presence of two subgroups within the same brood. Furthermore, the smaller-group individuals in the male desert population matured before the females. They primarily exhibited green coloration, while larger-group individuals matured after the females and primarily exhibited brown coloration. Females did not differ on average in either size or color (primarily green) between the two groups. We propose that S. viridis males have developed two distinct male strategies: the first strategy is to mature quickly, enabling early access to females, potentially less detectable to cannibalistic females, and requiring less food, but at the price of being smaller, senescing earlier in the season, and being less competitive against larger males. The second strategy allows males to become larger and better competitors, but at the price of maturing later, requiring more food, and reaching reproductive age once smaller males have already had access to potential mates. This fits with the phenomenon of mature individuals being greener when there is more seasonal green vegetation in their habitat, perhaps for better camouflage: the smaller, fast-maturing males were primarily green while the larger males, which matured later in the season, were primarily brown.
The presence of two separate male morphs in certain S. viridis populations is potentially reminiscent of alpha and beta male polymorphism in other insects, such as stag beetles, scarabs, and Diopsid flies. In these examples, a larger and more aggressive male morph monopolizes access to females, while a smaller “minor” morph relies on mobility and stealth to access females instead (Warren and Smith, 2007; Matsumoto and Knell, 2017; Parrett et al., 2021). In addition to insects, distinct male adaptations have been observed in certain isopods, prawns, reptiles, and fish, among others (Shuster, 1987; Stuart-Fox et al., 2020; Karplus and Barki, 2019; Partridge, 2021). However, this may be the first time two distinct male mating strategies have been identified in a mantis species. Alternatively, the two male morphs could have been explained by mixed paternity, the possibility that the female mated with two males. This could be examined and controlled under lab conditions.
Due to finding no pattern of fidelity in individual color choice for any of our populations, we assess that mantids reared in neutral conditions exhibit no preference for colors that more closely resemble their own phenotype. Ronsenheim et al. (2025) similarly found that when given a choice between green and brown, adult mantids do not match background color to their own color. In contrast, caterpillars raised in brown or green environments both turn that color and prefer it as a microhabitat (Eacock et al., 2019). The fact that we did not see a similar effect suggests that preference, if it exists, may come from gradual habituation to the particular color of its microhabitat, instead of being innately linked to the individual’s possessing the matching color morph. In a similar experiment with wild-caught polymorphic grasshoppers, Heinze et al. (2021) demonstrated that green morph grasshoppers prefer green microhabitats over brown ones, but that brown individuals have no such preference. The presence of color preference in wild-caught polymorphic insects in a laboratory setting supports the explanation that the development of color preference may be a gradual process. This is a potential explanation as to why individuals reared with distinct background colors seem to prefer them, while our experiment’s mantids, which were raised in neutral environments, did not. To confirm this, we suggest a follow-up study whereby S. viridis is reared in green or brown conditions, much like in Burke and Holwell (2023). Alternatively, it is possible that this mantid species does not exhibit background color choice.
Understanding the mechanism behind polymorphism development and color preference, as well as how it interacts with habitat selection, may offer insights into how some cryptic insects adapt to urbanization, as well as habitat and climate change. Polymorphic species are often quite tolerant of human activities and able to survive in urban environments. For instance, Sphodromantis viridis and Miomantis paykullii, two strongly polymorphic species, are common mantids in cities, towns, and gardens in Israel. Accordingly, we believe that S. viridis may be an ideal model to study adaptations to changing environments, such as those reflected by body color, size, and development time, particularly in urban habitats.
Funding
This study was supported by the Israel Science Foundation (grant no. 699/24 to IS).
Supplementary Material
The dataset is attached as an Excel file.
Conflict of interest
We declare no conflict of interest.
References
Bank, S., Cumming, R. T., Li, Y., Henze, K., Le Tirant, S., and Bradler, S. (2021). A tree of leaves: Phylogeny and historical biogeography of the leaf insects (Phasmatodea: Phylliidae). Communications Biology 4: 932.
Battiston, R. and Fontana, P. (2010). Colour change and habitat preferences in Mantis religiosa, Bulletin of Insectology 63: 85–89.
Bond, A. B. (2007). The evolution of color polymorphism: crypticity, searching images, and apostatic selection. Annual Review of Ecology, Evolution, and Systematics 38: 489–514.
Burke, N. W. and Holwell, G. (2023). Plastic background colour matching in the Springbok mantis. Functional Ecology 38: 449–464.
Cohen, N., Volov, M., Bodner, L., Bouchebti, S., and Levin, E. (2022). Body size, metabolic rate, and diapause in the Oriental hornet (Vespa Orientalis), in two extreme climatic regions. Ecological Entomology 8: 1–10.
de Alcantara Viana, J. V., Duarte, R. C., Lambertini, C., Capoccia, F., Martins, A. L. O., Vieira, C., and Romero, G. Q. (2024). Differential survival and background selection in cryptic trunk-dwelling arthropods in fire-prone environments. American Naturalist 204: E128–E145.
Duarte, R. C., Stevens, M., and Flores, A. A. (2016). Shape, colour plasticity, and habitat use indicate morph-specific camouflage strategies in a marine shrimp. BMC Evolutionary Biology 16: 1–15.
Dubiner, S., Namir, I., Chen, R., and Levin, E. (2023). Distance from human settlements favors wild-type appearance of feral cats (Felis catus) in Mediterranean woodland. Ecology and Evolution 13: e10261.
Eacock, A., Rowland, H. M., van’t Hof, A. E., Yung, C. J., Edmonds, N., and Saccheri, I. J. (2019). Adaptive colour change and background choice behaviour in peppered moth caterpillars is mediated by extraocular photoreception. Communications Biology 2: 286.
Edmunds, M. (1990). The evolution of cryptic coloration. In: Evans, D. L. and Schmidt, J. O. (eds.) Insect defenses: Adaptive mechanisms and strategies of prey and predators. State University of New York Press, Albany, NY, USA, pp. 3–21.
Forsman, A., Karlsson, M., Wennersten, L., Johansson, J., and Karpestam, E. (2011). Rapid evolution of fire melanism in replicated populations of pygmy grasshoppers. Evolution 65: 2530–2540.
Goldreich, Y. (2003). The Climate of Israel: Observations, Research and Application. Springer, New York, NY, USA.
Haque, M. T., Khan, M. K., and Herberstein, M. E. (2024). Current evidence of climate-driven colour change in insects and its impact on sexual signals. Ecology and Evolution 14: e11623.
Heinze, P., Dieker, P., Rowland, H. M., and Schielzeth, H. (2021). Evidence for morph-specific substrate choice in a green-brown polymorphic grasshopper. Behavioral Ecology 33: 17–26.
Karplus, I., and Barki, A. (2019). Male morphotypes and alternative mating tactics in freshwater prawns of the genus Macrobrachium: a review. Reviews in Aquaculture 11: 925–940.
Kingsolver, J., and Huey, R. (2008). Size, temperature, and fitness: three rules. Evolutionary Ecology Research 10: 251–268.
Köhler, G., and Schielzeth, H. (2020). Green-brown polymorphism in alpine grasshoppers affects body temperature. Ecology and Evolution 10: 441–450.
Kutiel, H. (2024). Climate of Israel. In: Frumkin, A., Shtober-Zisu, N. (eds.) Landscapes and Landforms of Israel (pp. 39–47). Springer, Cham, Germany.
Llandres, A. L., Figon, F., Christidès, J.-P., Mandon, N., and Casas, J. (2013). Environmental and hormonal factors controlling reversible colour change in crab spiders. Journal of Experimental Biology 216: 3886–3895.
Lo, Y. C., Blamires, S. J., Liao, C. P., and Tso, I. M. (2023). Nocturnal and diurnal predator and prey interactions with crab spider color polymorphs. Behavioral Ecology and Sociobiology 77: 19.
Matsumoto, K., and Knell, R. J. (2017). Diverse and complex male polymorphisms in Odontolabis stag beetles (Coleoptera: Lucanidae). Scientific Reports 7: 16733.
Okay, S. (1953). Formation of green pigment and colour changes in Orthoptera. Bulletin of Entomological Research 44: 299–315.
Parrett, J. M., Slade, E. M., and Knell, R. J. (2021). Morph-specific investment in testes mass in a trimorphic beetle, Proagoderus watanabei. Journal of Zoology 316: 169–177.
Partridge, C. (2021). Alpha, Beta, and Gamma Males. In: Shackelford, T. K., Weekes-Shackelford, V. A. (eds.) Encyclopedia of Evolutionary Psychological Science. Springer, Cham, Germany.
Pembury Smith, M. Q., and Ruxton, G. D. (2020). Camouflage in predators. Biological Reviews 95: 1325–1340.
Raxworthy, C. J., Pearson, R. G., Zimkus, B. M., Reddy, S., Deo, A. J., Nussbaum, R. A., and Ingram, C. M. (2008). Continental speciation in the tropics: Contrasting biogeographic patterns of divergence in the Uroplatus leaf-tailed gecko radiation of Madagascar. Journal of Zoology 275: 423–440.
Rosenheim, L. Y., Rosenheim, J. A., and Maxwell, M. R. (2025). Coloration in a praying mantis: Color change, sexual color dimorphism, and possible camouflage strategies. Ecology and Evolution 15: e70398.
Scharf, I., Filin, I., Ben-Yehoshua, D., and Ovadia, O. (2009). Phenotypic plasticity and variation in morphological and life-history traits of antlion adults across a climatic gradient. Zoology 112: 139–150.
Scharf, I., Filin, I., Golan, M., Buchshtav, M., Subach, A., and Ovadia, O. (2008). A comparison between desert and Mediterranean antlion populations: differences in life history and morphology. Journal of Evolutionary Biology 21: 162–172.
Schwarz, R., Stark, G., Levin, E., Chapple, D. G., and Meiri, S. (2023). Ecophysiological trait variation in desert versus Mediterranean populations of a gecko. Journal of Zoology 321: 260–277.
Shuster, S. M. (1987). Alternative reproductive behaviors: Three discrete male morphs in Paracerceis sculpta, an intertidal isopod from the northern Gulf of California. Journal of Crustacean Biology 7: 318–327.
Stevens, M., and Ruxton, G. D. (2019). The key role of behaviour in animal camouflage. Biological Reviews 94: 116–134.
Stuart-Fox, D., Aulsebrook, A., Rankin, K. J., Dong, C. M., and McLean, C. A. (2020). Convergence and divergence in lizard colour polymorphisms. Biological Reviews 96: 289–309.
Théry, M., Debut, M., Gomez, D., and Casas, J. (2004). Specific color sensitivities of prey and predator explain camouflage in different visual systems. Behavioral Ecology 16: 25–29.
Théry, M., and Gomez, D. (2010). Insect colours and visual appearance in the eyes of their predators. Advances in Insect Physiology 38: 267–353.
Trullas, S. C., van Wyk, J. H., and Spotila, J. R. (2007). Thermal melanism in ectotherms. Journal of Thermal Biology 32: 235–245.
Tsurui, K., Honma, A., and Nishida, T. (2010). Camouflage effects of various colour-marking morphs against different microhabitat backgrounds in a polymorphic pygmy grasshopper Tetrix japonica. PLoS One 5: e11446.
Volis, S., Mendlinger, S., and Ward, D. (2002). Adaptive traits of wild barley plants of Mediterranean and desert origin. Oecologia 133: 131–138.
Wang, Y. J., Wu, J., and Zhao, D. X. (2017). Factors influencing color changes in Hierodula patellifera (Mantodea: Mantidae). Journal of Entomological Science 52: 92–93.
Warren, I., and Smith, H. (2007). Stalk-eyed flies (Diopsidae): Modelling the evolution and development of an exaggerated sexual trait. BioEssays 29: 300–307.
Zeuss, D., Brandl, R., Brändle, M., Rahbek, C., and Brunzel, S. (2014). Global warming favours light-coloured insects in Europe. Nature Communications 5: 3874.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}