Abstract
LifeinU⢠Bacillus subtilis CU1 (BSCU1) has been previously shown to be effective in stimulating mucosal immune responses and supporting resistance to common infectious disease episodes in the elderly. The current clinical study aimed at exploring potential pathways by which BSCU1 could beneficially modulate the immune system and contribute to protection against infection in the general population. A total of 88 participants from three different age groups were supplemented with BSCU1 (2 à 109 cfu/day) for 4 weeks. The effect of the intervention on mucosal immunity was assessed by faecal sIgA levels. In addition, a series of complementary immunoassays were selected, including immune phenotyping, gene expression, basal cytokine levels, cytokine levels in lipopolysaccharide (LPS)-stimulated whole blood and phagocytosis assay. Although no significant effect was observed on faecal sIgA levels after intervention, BSCU1 showed a positive effect on a consistent set of markers of the peripheral innate immune system in adults and the elderly. Percentages of peripheral blood myeloid cells as well as the expression of the activation marker CD69 on monocytes were significantly increased after probiotic intervention. BSCU1 supplementation resulted in significant enrichment of clusters of genes involved in response to type I interferon and phagocytosis pathway. Consistently, ex vivo stimulation of whole blood with LPS resulted in a statistically significant increase in pro-inflammatory cytokines (interleukin (IL)-1beta, IL-6, interferon-gamma, IL-12, tumour necrosis factor (TNF)-alpha, macrophage inflammatory protein (MIP)-1alpha, IL-8) and phagocytosis assays showed increased capacity of monocytes to engulf bacteria as well as higher phagosome maturation. BSCU1 supplementation also had a positive effect on low-grade inflammation as significant reduction in basal levels of several serum cytokines (IL-10, TNF-alpha, MIP-1alpha, IL-8) were observed in the elderly subgroup. Overall, BSCU1 primed immune cells for a better response to microbial challenges and reduced low-grade inflammation associated with aging.
Registered at ClinicalTrials.gov with the identifier NCT05403398
1 Introduction
The human gastrointestinal tract hosts a highly diverse microbial ecosystem, collectively referred to as gut microbiota, that plays a crucial role in human health and disease (Flint et al., 2012). It affects numerous aspects of host physiology from digestion and metabolism to neurobiology (Fan and Pedersen, 2021; Sharon et al., 2016). The human gut microbiota strongly influences development and homeostasis of the immune system and this interplay has a major role in the control and response to infection (Belkaid and Harrison, 2017; Gensollen et al., 2016; Pickard et al., 2017). It contributes to the resistance and clearance of infection, both directly through mechanisms of colonisation resistance (e.g. competition for resources and space, production of antimicrobial substances or stimulation of mucus production) and indirectly through modulation of mucosal immunity (Perez-Lopez et al., 2016). The role of the gut microbiota in immunity also extends to systemic compartments and distant organs such as the lung (Budden et al., 2017; McAleer and Kolls, 2018). The symbiotic relationship between the gut microbiota and the host is a fragile equilibrium that can be affected by several factors (e.g. lifestyle habits, diet or antibiotics use). A divergence from the normal microbiota composition, generally referred to as dysbiosis, can substantially affect mucosal and systemic immunity (Levy et al., 2017). Owing to the well documented impact of the gut microbiota on immune health, there is an interest in understanding the role of microbiota modulation in health and disease. Of the available solutions, probiotics, which are defined as âLive microorganisms that, when administered in adequate amounts, confer a health benefit on the hostâ (Hill et al., 2014), have been shown to modulate immune system function (Lehtoranta et al., 2020). Probiotic effects on immune health are strain-dependent and different lactic acid bacteria and bifidobacteria have been described to induce very different maturation and cytokine production patterns in dendritic cells (DCs) and even to elicit opposing T-cell responses. Certain probiotics have been shown to induce the production of interleukin (IL)-12 by macrophages and DCs, promote interferon (IFN)-γ secretion, and favour inflammatory Th1 responses (Fujiwara et al., 2004; Mohamadzadeh et al., 2005; Pochard et al., 2005), whereas other reports show that probiotic-induced IL-10 levels play an important role in limiting Th1-mediated proinflammatory responses (Hart et al., 2004; Zeuthen et al., 2006). There are few published reports addressing the health benefits of probiotic spore-formers, mostly belonging to the genus Bacillus. Bacteria of this genus can form spores that enable them to survive harsh environmental conditions. This ability gives them a significant advantage over the most common lactic acid probiotics (Lactobacillus and Bifidobacterium), because in their spore form, Bacillus probiotics can survive the acidic conditions of the stomach and spread safely throughout the nutrient-rich small intestinal tract. In addition, Bacillus probiotics are of growing interest because they are stable in commercial foods for long periods of time (Cutting, 2011). The efficacy of Bacillus spp. as probiotics has been studied in several in vitro and in vivo animal models, and a few have also been validated in human clinical trials. Available data from these studies have demonstrated the beneficial effects of various strains of Bacillus spp. on human health. For example, several researchers have recognised the preventive role of Bacillus probiotics in intestinal disorders (Lopetuso et al., 2016; Zhang et al., 2016). Bacillus subtilis species have long been safely consumed as dietary supplements and food ingredients in various parts of the world (Lefevre et al., 2017). A specific strain of B. subtilis, CU1 (CNCM I-2745) has been described as an effective probiotic in stimulating immune responses of healthy elderly subjects (Lefevre et al., 2015). Probiotic supplementation with B. subtilis CU1 was found to decrease the occurrence of infectious episodes in healthy free-living elderly subjects during a period of common infectious disease (i.e. the winter season in France). These effects were attributed to the ability of B. subtilis CU1 to enhance peripheral as well as intestinal and respiratory mucosal immune responses, as indicated by the increased concentrations of faecal and salivary secretory IgA and serum interferon-gamma. Postulated mechanisms of actions of B. subtilis CU1 include stimulation of the immune system, production of antimicrobials and modulation of the composition of gut microbiota (DudonneÌ et al., 2015; Lefevre et al., 2015; Urdaci et al., 2004; VilleÌger et al., 2014). However, the precise mechanisms by which B. subtilis CU1 exert its positive probiotic effects remains relatively unexplored.
In the current study, we aimed to investigate whether B. subtilis CU1 can positively modulate the mucosal and peripheral immune system in three target populations representing the general population. A series of complementary immunoassays were selected, including immune phenotyping, gene expression, cytokine levels in LPS-stimulated whole blood and phagocytosis assay.
2 Materials and methods
Study design
This exploratory study was designed as a single-arm study with repeated measures, involving three different populations, in which each subject served as its own control. This clinical trial was conducted between June 2022 and July 2022. The duration of the intervention was 4 weeks, with biomarker assessments at baseline, after 2 weeks, and after 4 weeks of intervention. Participants visited the study site (NIZO, Ede, the Netherlands) four times, once for information on the trial and subjectsâ eligibility assessment (screening visit), once for baseline measurements (visit 1, V1), after two weeks of probiotic supplementation (visit 2, V2) and at the end of the four-week intervention (visit 3, V3).
Participants, ethics and trial registration
This human intervention study was approved by the Medical Ethics Committee of Brabant, Tilburg, the Netherlands and registered as protocol NL79890.000.21. In addition, the clinical trial was registered in a clinical trials registration system at ClinicalTrials.gov with the identifier NCT05403398. The clinical trial was performed according to the ethical principles of the Declaration of Helsinki. The study included three separate subgroups, primarily defined by age: healthy children between 3 and 6 years; healthy adults between 30 and 49 years; healthy elderly between 65 and 79 years. Participants were excluded if they had an acute respiratory or gastrointestinal tract infection, any vaccination, antibiotic treatment, or regular intake of probiotics within 1 month prior to the start of the study. The use of anti-inflammatory or immunosuppressive medications was also an exclusion criterion. For complete inclusion and exclusion criteria lists for the three subgroups, see Supplementary Data S1. Signed written informed consent for the study was obtained from all adults and elderly subjects and from parents/caretakers of the children before protocol-specific procedures were carried out.
Investigational product
The probiotic product consists of Bacillus subtilis CU1 (BSCU1) mixed with excipients (maltodextrin and magnesium stearate). Each probiotic capsule contains 2 billion spores of BSCU1 (LESAFFRE, Marcq en Baroeul, France). BSCU1 is a specific strain exclusive to Lesaffre and registered in the CNCM collection under the number I-2745. Capsules were taken orally, as a food supplement. The supplement was provided in a formulation for clinical study use in a daily dosage (2Â Ã 109 cfu/day). The capsules were supplied to the study subjects and parents/caretakers, respectively, during study visit 1, and participants were requested to consume 1 capsule daily in the morning (just before breakfast). For the children, caretakers were asked to empty the capsule into a small glass of a cold drink (e.g. water or milk) or yogurt and make sure that all of it was consumed. Compliance was determined through the assessment of returned packaging and interviews of the subjects at each visit. Subjects were instructed to maintain their habitual lifestyle and the consumption of any other probiotic was forbidden during the entire study period.
Study endpoints
The primary endpoint was the change after 4 weeks of probiotic supplementation in faecal secretory immunoglobulin A (sIgA) concentration. Secondary endpoints were the change from baseline to 4 weeks in circulating levels of cytokines, in ex vivo production of cytokines in a control and lipopolysaccharide (LPS) stimulated condition, and in ex vivo phagocytic capacity of monocytes and granulocytes. Additionally, the effect of 4 weeks supplementation with BSCU1 was assessed on immune cell phenotypes and activation markers and on gene expression in peripheral blood mononuclear cells (PBMC). The effect of probiotic intervention was also assessed on gut microbiota composition and activity.
Biological samples collections
Faecal samples were collected at V1, V2 and V3 in the 3 subgroups. Faecal samples were collected within 24 h before each visit. The adult subjects and the childrenâs caregivers were given all the materials and instructions to collect a faecal sample. Approximately 1-2 g of stool were collected at each visit. Of the stool samples, 100 mg was used for sIgA analysis and 250 mg for DNA isolation. Immediately after collection, the stool sample was frozen at â20 °C in the subjectâs home freezer. For this purpose, an envelope and materials for transport were provided to prevent the sample from thawing during transport.
Fasting blood samples were collected at V1 and V3 in adults and elderly only (not performed in the healthy children subgroup for ethical reason).
Immune markers
Faecal sIgA
Faecal sIgA levels were measured using the IDK sIgA ELISA kit (Immundiagnostik, Bensheim, Germany). A faecal sample of 100Â mg was diluted in 10Â ml ELISA buffer and further processed according to the manufacturerâs protocol.
Serum cytokine quantification
The following cytokines were analysed IL-1beta, IL-6, IL-8, IL-10, IL-12, IFN-gamma, macrophage inflammatory protein (MIP)-1alpha, and tumour necrosis factor (TNF)-alpha. Cytokine concentration was assessed with Bio-Plex Pro⢠Human Cytokine Assay (BioRad, Hercules, CA, USA) according to the manufacturerâs instructions. Fluorescence was measured with Bio-Plex 200 Array reader and Bio-Plex Manager software 6.1.
BD Vacutainer CAT 2Â ml PET tube with clot activator (BD Biosciences, Vianen, the Netherlands, BD 368271) were used for serum preparation. Samples were incubated for 30 minutes at room temperature and centrifuged for 10Â min at 2,000
Ex vivo cytokine quantification
Heparinized whole blood was collected and used within 1 h. Blood was diluted 1:5 in Roswell Park Memorial Institute (RPMI) 1640 Medium (ThermoFisher, Bleiswijk, the Netherlands) in a 96 wells plate and incubated for 20 h with and without 100 ng/ml LPS (LPS from Escherichia coli O111:B4, Merk Life Sciences NV, Amsterdam, the Netherlands) at 37 °C in a humidified atmosphere containing 5% CO2. After incubation, samples were centrifuged at 2,000
Ex vivo phagocytosis assay
Ex vivo phagocytosis in whole blood was studied using pHrodo⢠Green E. coli BioParticles⢠(ThermoFischer Scientific, Waltham, MA, USA) according to the manufacturerâs instructions. Fluorescence was measured using FACSCanto II (BD, Franklin Lakes, NJ, USA). Samples were analysed using FlowJo⢠Version 10 (FlowJo LLC, Ashland, OR USA).
Immune cell phenotyping
This analysis was performed at Wageningen University, Cell Biology and Immunology group (the Netherlands). Blood samples were collected in BD Vacutainer CPT tubes. After blood withdrawal, tubes were stored at room temperature and transported to Wageningen University. Tubes were centrifuged for 25Â min at 1,800

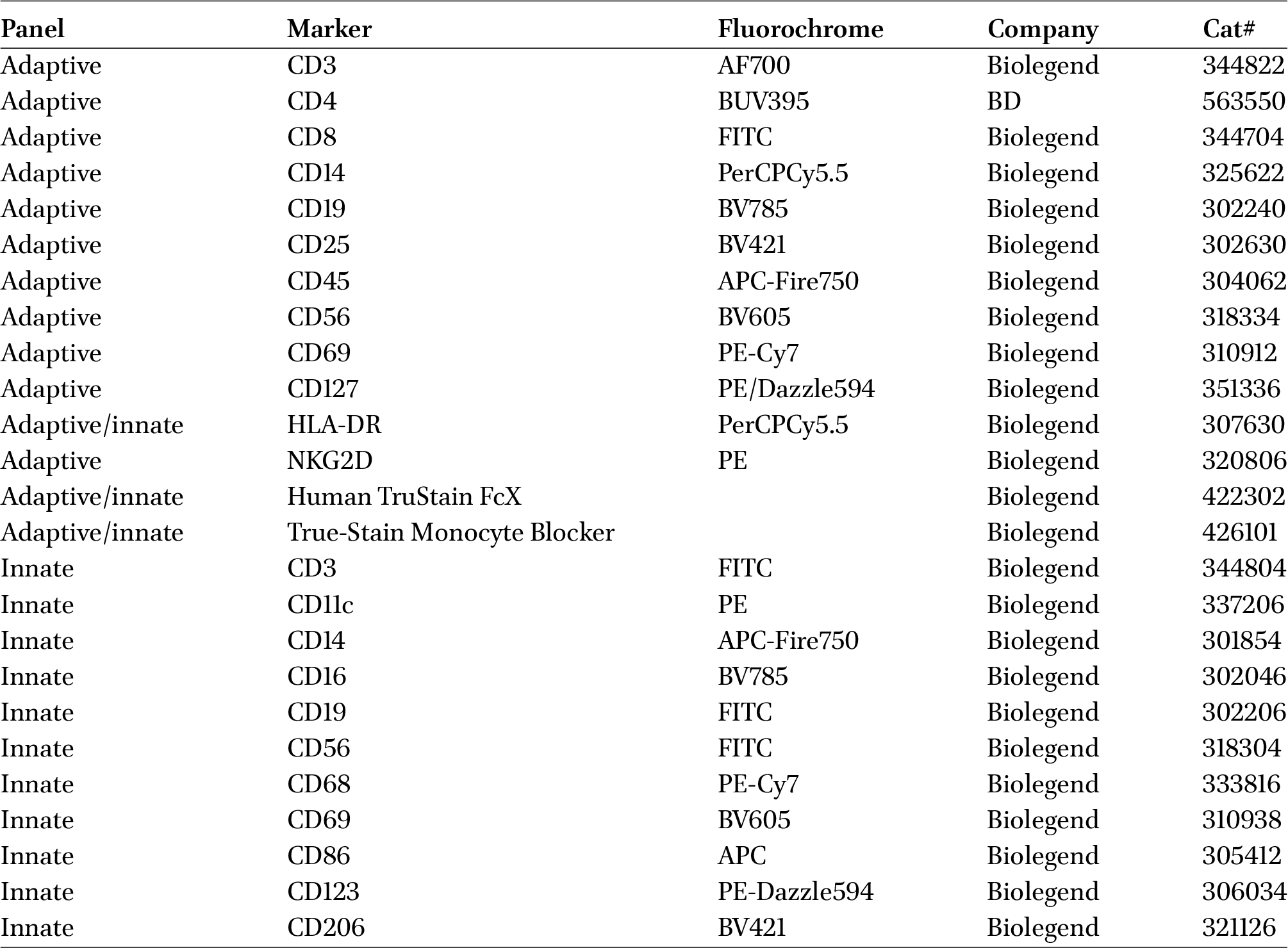
Antibodies used for immune phenotyping
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
PBMC gene expression assessment
Blood was collected in BD Vacutainer 10 ml PET heparin tubes. PBMCs were isolated using Leucosep® tubes (Greiner Bio-One, Kremsmünster, Austria), prefilled with separation medium according to the manufacturerâs instructions. Heparinized blood (10 ml) was 1:2 diluted with Dulbeccoâs phosphate-buffered saline (DPBS) and poured into the Leucosep® tube. The sample was centrifugated for 10 min at 1000
RNA isolation was performed with RNeasy Mini Kit (Qiagen, US) according to the manufacturerâs instructions. After lysis and precipitation, the samples were loaded onto the RNeasy column and subjected to DNAse treatment to remove DNA. RNA was eluted in RNAse-free water and stored at â80 °C in multiple aliquots for quality control and library preparation. RNA concentration was measured by the NanoDrop ND-1000 (Thermo Scientific). Further, 10% of the samples were analysed at the Agilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany).
RNA samples were shipped on dry ice to Novogene UK (Milton, UK). Directional mRNA libraries were prepared by Novogene (Poly A enrichment) and sequenced on an Illumina NovaSeq 6000 PE150 platform (Illumina, San Diego, CA, USA) to obtain at least 6Â Gb raw PE150 data per sample. Quality-controlled data files in FASTQ format were transferred to NIZO for bioinformatics analysis.
For RNAseq analysis, a total of 118 RNA samples were collected from two distinct populations, adults and the elderly. These samples were analysed using the nf-core/rnaseq pipeline version 3.12.0 (https://nf-co.re/rnaseq/3.12.0), which encompasses a series of steps. Initially, a preprocessing step was performed using tools such as FastQC, TrimGalore, BBsplit, and SortMeRNA to ensure quality control. Then, reads were aligned to the reference genome and gene expression levels were quantified using the STAR tool. Post-processing steps included count generation using featureCounts and Kalisto, followed by additional processing and a final quality control assessment using RSeqQC, DESEq2, and MultiQC. The resulting data were then subjected to a differential analysis using DESEq2 version 1.38.3, where genes with a log fold change greater than 1.5 or less than â1.5 and a P-value of 0.05 were considered differentially expressed. A Gene Set Enrichment Analysis (GSEA) was also conducted using the fgsea package in R version 4.1.0.
Faecal microbiota composition
The faecal DNA isolation method is based on mechanical disruption. An aliquot of 250 mg of faecal material was placed in 700 μl STAR buffer (Roche) with 0.5 g of 0.1 mm Zirconia beads. Lysis was performed with the FastPrep at 5.5 ms and room temperature for 3 à 1 min followed by heat treatment (95 °C for 15 min shaken at 100 rpm). Samples were centrifuged for 10 min at 4 °C and 21,000
DNA libraries were prepared by Novogene and sequenced on an Illumina NovaSeq 6000 PE150 platform to obtain at least 2Â Gb raw PE150 data per sample. Quality-controlled data files in FASTQ format were transferred to NIZO for bioinformatics analysis.
Raw sequencing data was quality controlled using fastqc (v0.11.9). Reports for quality controlled reads were combined using multiQC (v1.12). Quality controlled reads were then subjected to the Humann pipeline (v3.6). Multivariate analyses consisted in Principal Coordinate Analysis (PCoA) on Bray Curtis beta-diversity. Statistical significance and the proportion of microbial composition variance explained by different effects and covariates was obtained using PERMANOVA. Statistics were performed with R (v4.2.3). Multivariate analysis was performed using the Maaslin2 (v1.8.0) and FactoMineR (v1.34) packages. Graphics were produced using ggplot2 (v3.4.0). Alpha and Beta-diversity were analysed using the vegan package (v2.6-4).
Safety
Adverse events were recorded during the study by investigators.
Statistical analysis
Sample size
The primary outcome of this study was faecal sIgA level. With a sample size of n = 30 (with 20% drop-out), it is possible to detect an increase of 54% in the levels of faecal sIgA with a power of 80%, in both the adult and elderly populations (Lefevre et al., 2015). For children, a sample size of 21 subjects enables the detection of an increase of 50% in the levels of faecal sIgA with a power of 80% (BeÌghin et al., 2021).
Efficacy analyses
Analyses were performed on the per-protocol population (PP). PP population was defined per timepoint. Decisions about the allocation of subjects to the PP were taken during a data review meeting. The database was locked after the completion of the data review meeting and decisions were documented in the minutes of the data review meeting. All data were analysed with Linear Mixed Models (LMMs) with adjustment of analysis on age, gender and body mass index (BMI) at baseline. Model goodness-of-fit were assessed using the Akaike Information Criterion and model assumptions (linearity, homoscedasticity, linearity of residuals, non-collinearity of parameters) were checked. In case of non-compliance to the model assumptions, log transformations were performed. Hochberg adjustments were used to correct for multiple testing.
Statistical analyses were performed using SAS® software version 9.4 (TS1M7) (SAS Institute Inc., Cary, NC, USA) and R software version 4.2.2 (R Foundation).
3 Results
Study flowchart and baseline characteristics of the population
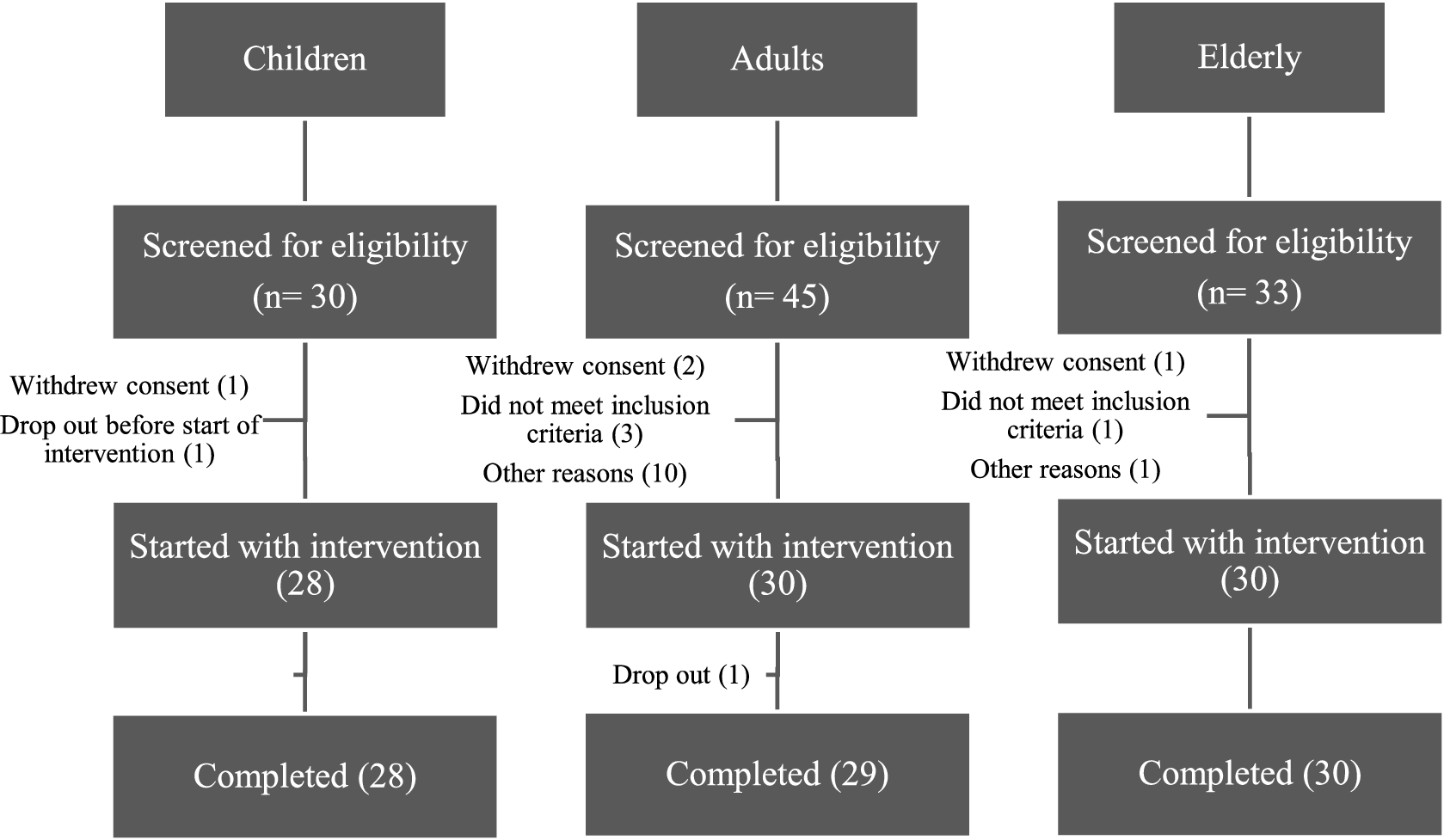
One hundred and eight (108) subjects were screened for study eligibility and a total of eighty-eight (88) participants were included in this study. The flowchart of subjects throughout the study is presented in Figure 1. A total of 28 children, 29 adults and 30 elderly completed the entire study. A total of 24 children, 24 adults and 27 elderly were included in per-protocol (PP) population at baseline. The baseline characteristics of the subjects are presented in Table 2. The main reasons for exclusion of subjects from the PP population were missed biological samples, occurrence of infectious diseases or use of medication that may have interfered with the outcome parameters of the study. Excellent mean compliance was observed (>99%).
Baseline characteristics of participants included in the per protocol group1
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
BSCU1 intake does not alter the composition and function of the gut microbiota in children, adults, and the elderly
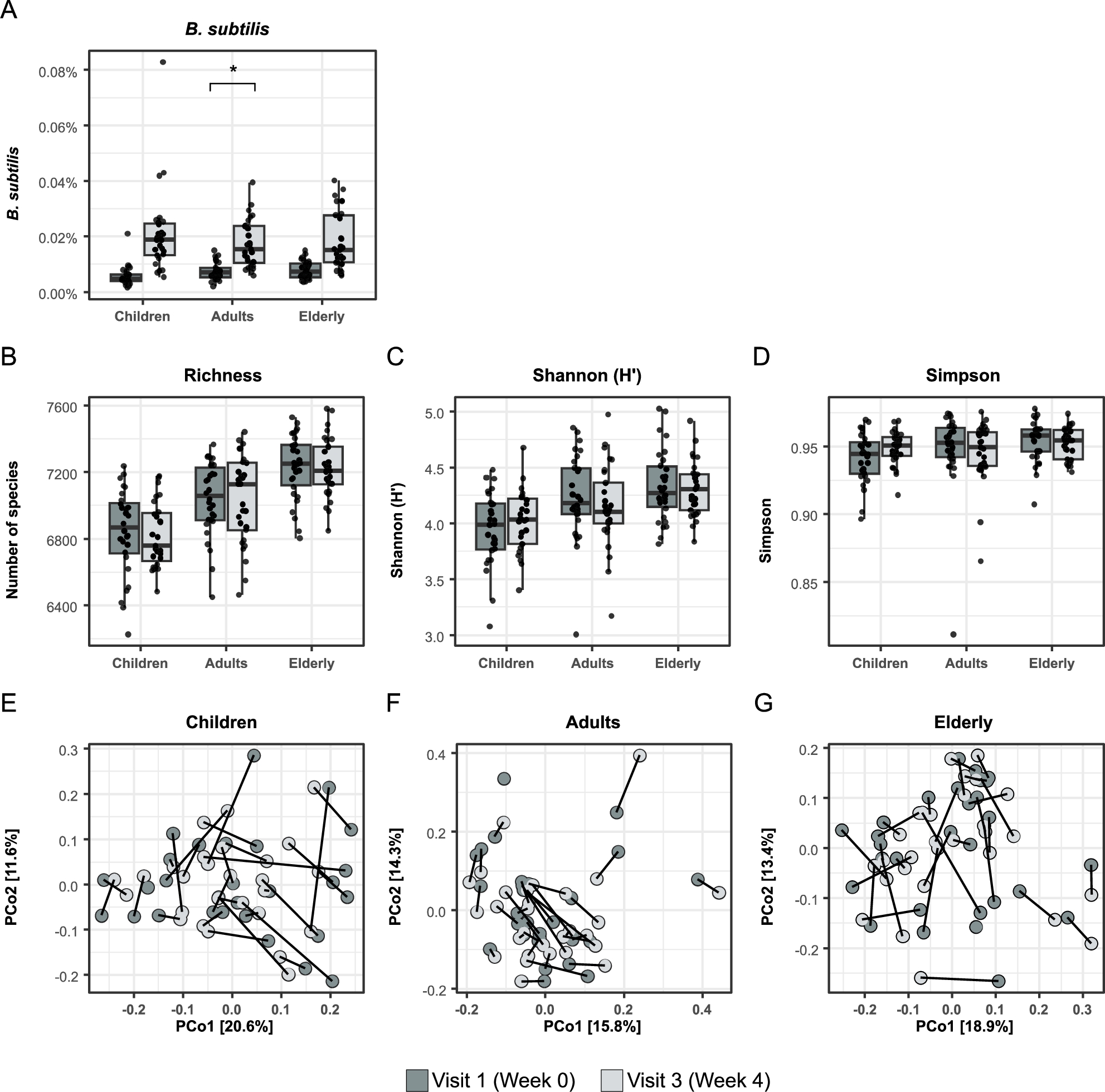
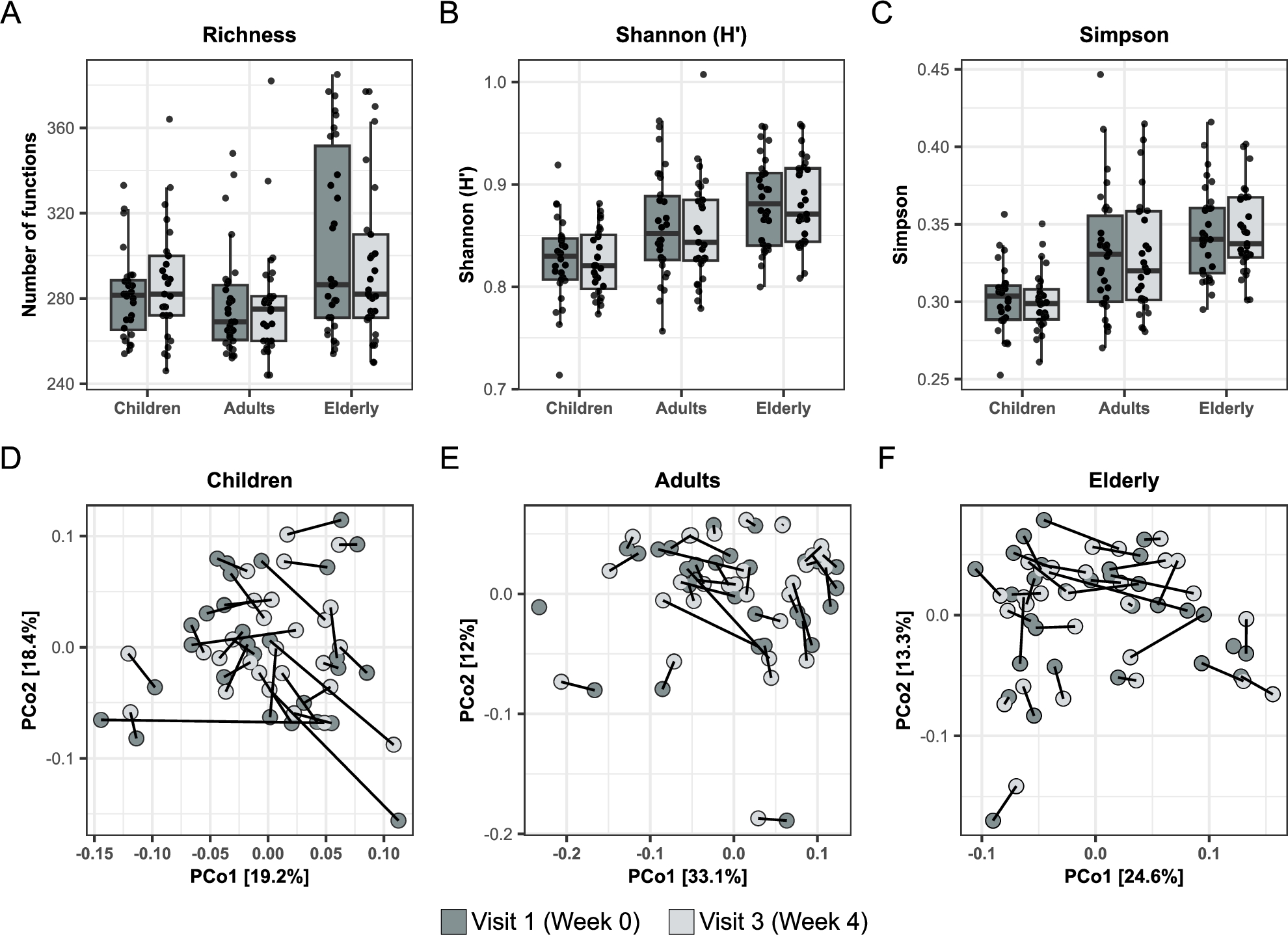
Effects of oral treatment with BSCU1 on the gut microbiota composition and functionality are shown in Figure 2 and Figure 3, respectively. As expected, the relative abundance of B. subtilis was significantly increased after treatment. However, when correcting for false discovery rate then only adults showed a significant increase in this bacterium (Figure 2A). No significant differences were observed in microbiota composition (Figure 2B-G) nor functionality (Figure 3A-F) in any groups and any time point comparison.
Effect of BSCU1 on microbiota composition. (A) Bacillus subtilis abundance in faecal samples before and after treatment, (B-D) Diversity indices and (E-G) PCoA analysis (*
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Effect of BSCU1 on microbiota functionality. (A-C) Diversity indices and (D-F) PCoA analysis before and after treatment.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Oral supplementation with BSCU1 tends to increase faecal sIgA content in children, adults, and the elderly
The effect of oral supplementation with BSCU1 on faecal sIgA levels is presented in Figure 4 and Table 3. There was a trend toward increased faecal sIgA production in children and the elderly 2 weeks after BSCU1 ingestion, with increases from baseline of 24% and 29%, respectively. This trend was also observed 4 weeks after the start of the probiotic supplementation in the three groups studied, with increases of 24%, 36%, and 20% in children, adults, and the elderly group, respectively, as compared to the baseline. BSCU1 ingestion thus seemed to increase faecal sIgA but these trends did not reach statistical significance.
Faecal sIgA levels in children, adults and the elderly. The sIgA content was determined at baseline and 2 and 4 weeks after the first intake of BSCU1 in each group. Data show means values ± standard error.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
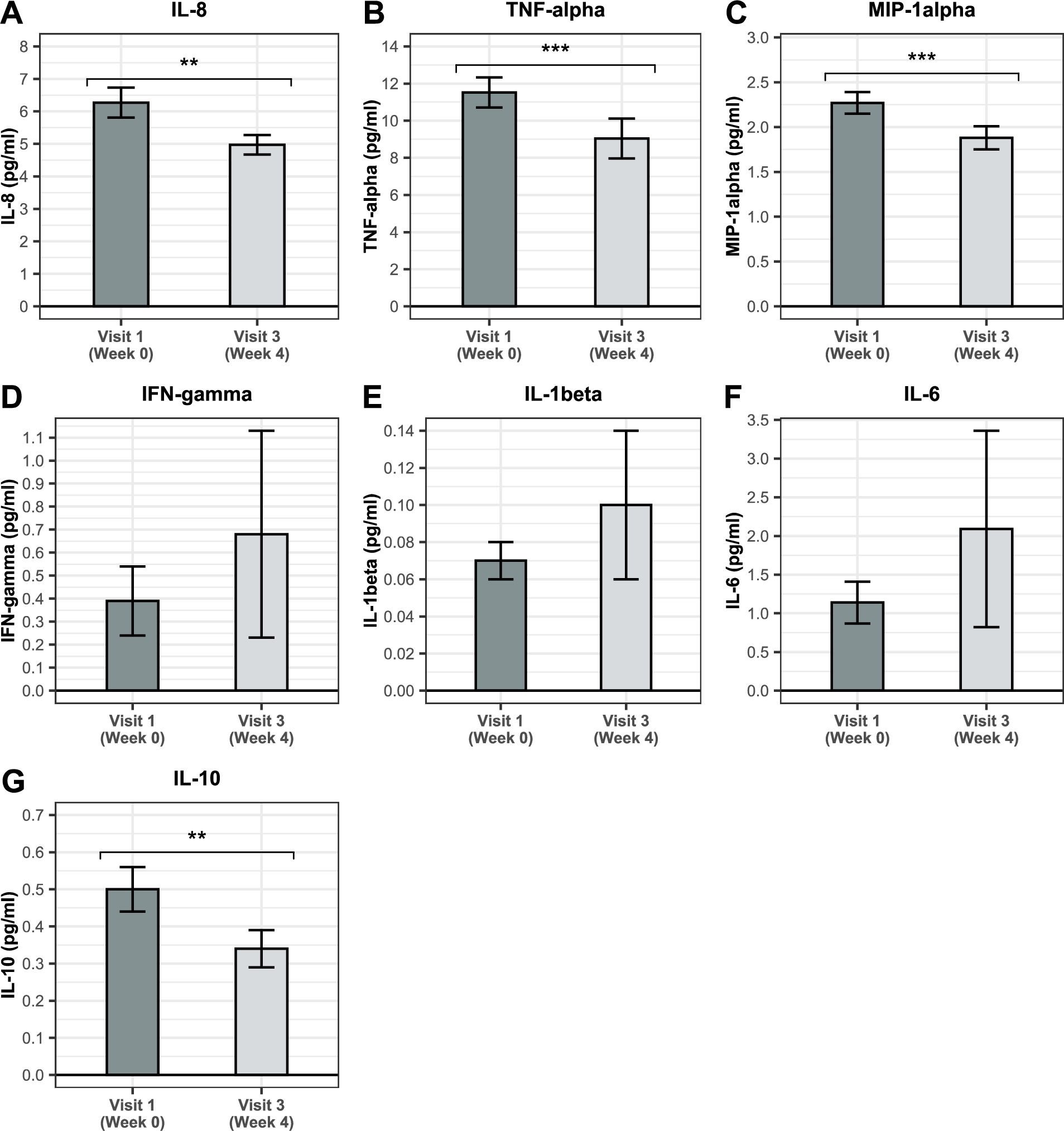
BSCU1 supplementation reduces low-grade inflammation in the elderly
A chronic systemic inflammatory state occurs to varying degrees during aging (Chung et al., 2019). Therefore, in the current study, the effect of BSCU1 supplementation on low-grade inflammation was assessed only in the elderly group (Figure 5). Levels of IL-8, TNF-alpha, and MIP-1alpha significantly decreased by 21%, 22%, and 17%, respectively, after supplementation with BSCU1. IFN-gamma, IL-1beta, and IL-6 were not affected. A statistically significant reduction in IL-10 level was also observed after intervention with BSCU1. IL-12 was under detection threshold.
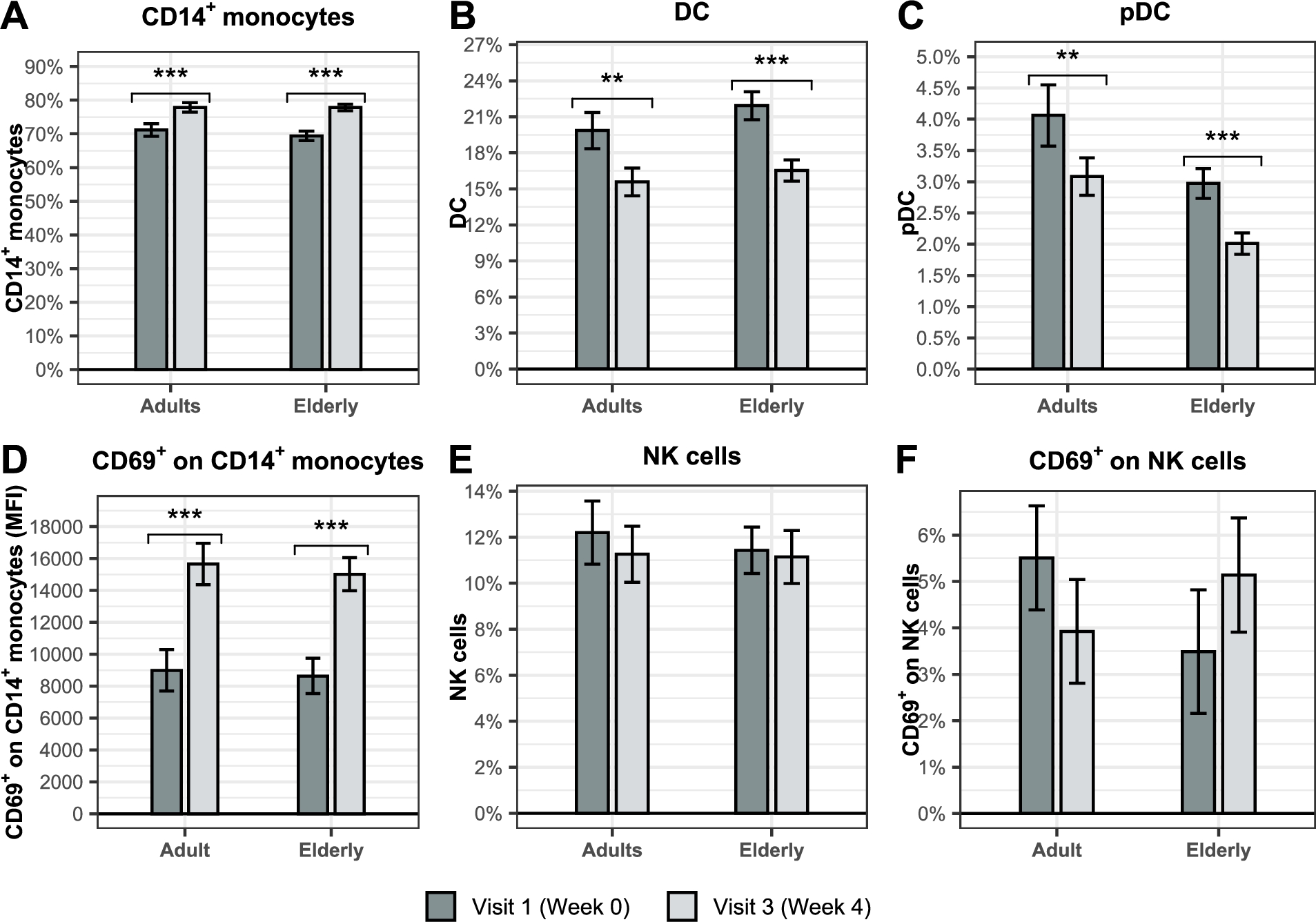
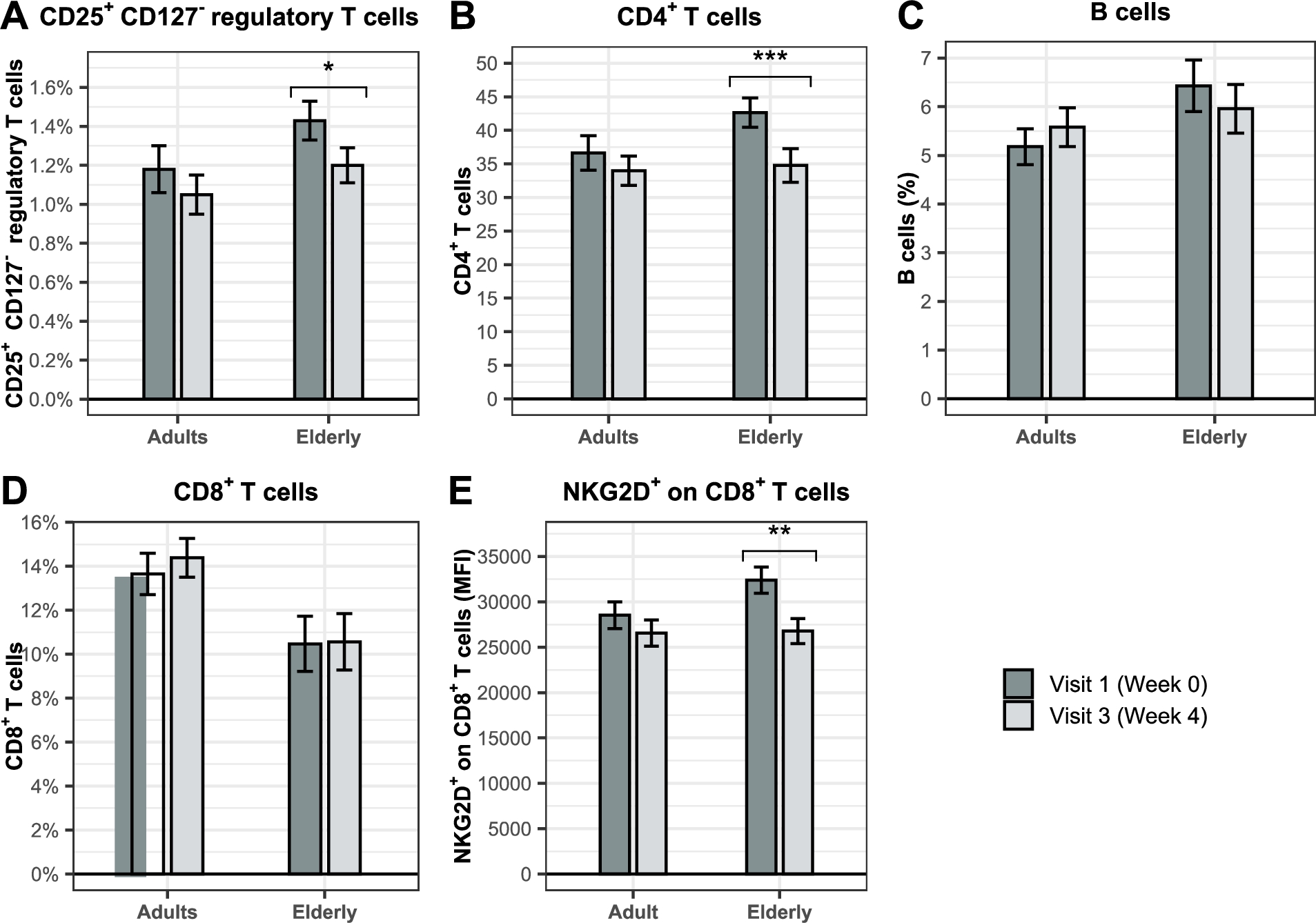
BSCU1 uptake modulates immune cell relative abundance in adults and the elderly
We further investigated by flow cytometry whether treatment with BSCU1 influenced the relative abundance of innate (Figure 6) and adaptive immune cells (Figure 7) in adult and elderly PBMCs. CD14+ monocytes were the predominantly affected cell types (Figure 6A). A significant increase of 6.6 and 8.2 percentage points were observed in the adult and elderly groups, respectively. The increase in the relative abundance of monocytes was accompanied by a significant decrease of 4 and 5.2 percentage points in conventional dendritic cells (DC) (Figure 6B) and 1 percentage point in plasmacytoid dendritic cells (pDC) in the adult and elderly groups, respectively (Figure 6C). Importantly, we observed an increase in the mean fluorescence intensity of the activation marker CD69 expression at the cell surface of CD14+ monocytes by 74.2% and 73.9% in adult and elderly groups, respectively (Figure 6D). The relative abundance of natural killer (NK) cells and their activation status were not affected by treatment with BSCU1 (Figure 6E-F). A significant decrease in CD25+ CD127â regulatory T cells (Tregs) and CD4+ T cells was observed in the elderly consuming BSCU1 (Figure 7A-B). There were no biologically significant changes in circulating B cells and CD8+ T cells (Figure 7C-D). Expression of NKG2D, an activating receptor mainly found on cytotoxic immune cells, was decreased at the cell surface of CD8+ T cells in elderly patients (Figure 7E). Taken together, these results suggest an immunomodulation by BSCU1 affecting mainly T cells and myeloid cells.
Effects of BSCU1 on faecal sIgA levels after 4 weeks of intervention
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Change in circulating cytokines in the elderly. Data show means ± standard error. P-values: ** [0.001 to 0.01[; *** <0.001.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Effect of BSCU1 on relative abundance and activation status of innate immune cells. Data show means ± standard error. P-values: ** [0.001 to 0.01[; *** <0.001.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Effect of BSCU1 on relative abundance and activation status of adaptive immune cells. Data show means ± standard error. P-values: * [0.01 to 0.05[; ** [0.001 to 0.01[; *** <0.001.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
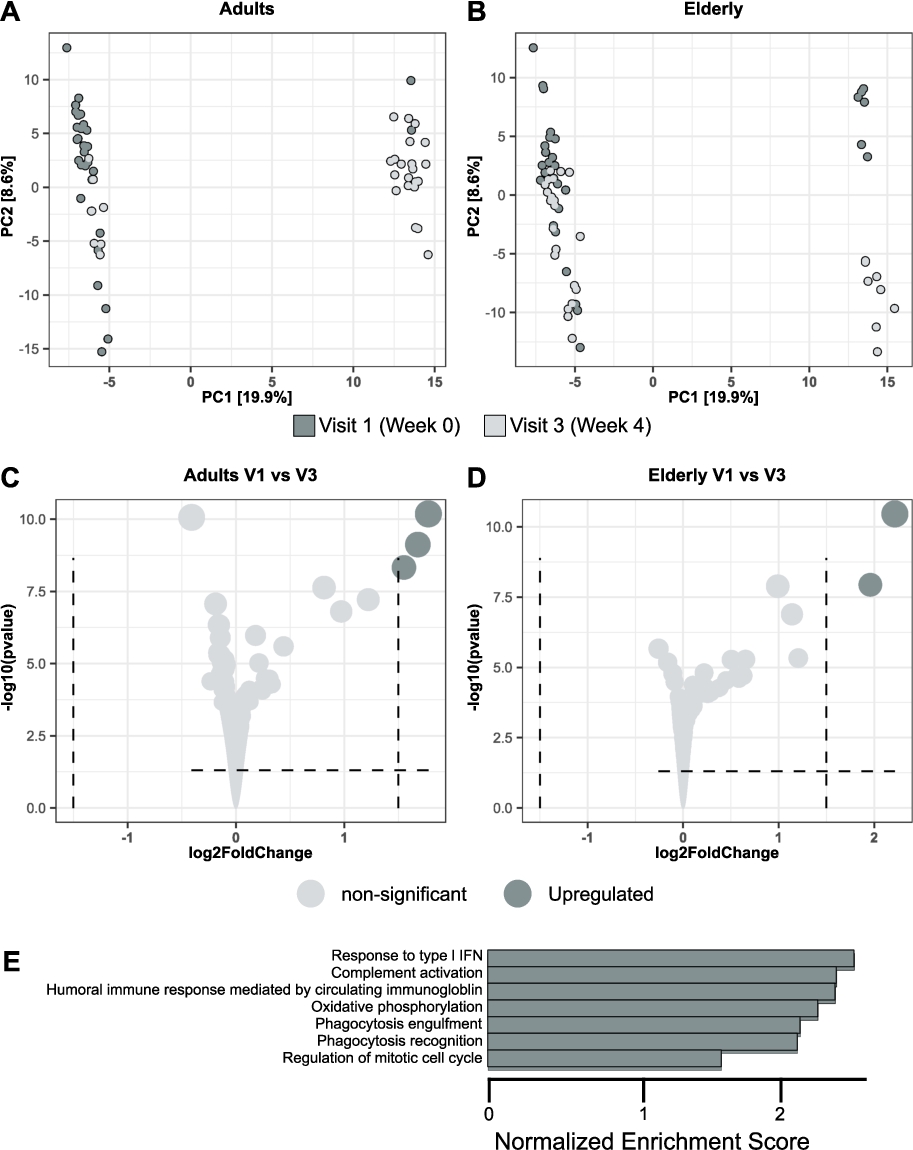
BSCU1 supplementation modulates gene expression program in immune cells
To further explore how supplementation with BSCU1 affects the gene expression program of immune cells, bulk RNA sequencing on PBMCs obtained at baseline and after 4 weeks of treatment has been performed in adult and elderly groups (Figure 8). Principal component analysis (PCA) revealed that 29% of the information was in PC1 and PC2, with more individuals segregating with PC2 in the adult group than in the elderly group, suggesting a more profound change in the PBMC gene expression program in adults compared to elderly (Figure 8A-B). The volcano plots for both the adult and elderly groups showed moderate modulation of gene expression, still a log2 fold change superior of 1.5 for upregulation of the inflammatory IL-8 cytokine, EGR1 (a zinc finger transcription factor essential for monopoiesis (Trizzino et al., 2021)) and G0S2 (a gene recently suggested to be involved in the inflammatory response of mononuclear cells (Kobayashi et al., 2008)) transcripts (Figure 8C-D). Gene Ontology (GO) term analysis in the adults showed enrichment in several pathways including âresponse to type I IFNâ, âcomplement activationâ, âoxidative phosphorylationâ, âphagocytosis recognitionâ and âphagocytosis engulfmentâ upon BSCU1 intake (Figure 8E). Thus, supplementation with BSCU1 resulted in changes in the gene expression program related to monocytes and their activation status.
BSCU1 supplementation modulates gene expression program in immune cells. Differential gene expression (DGE) analysis of the effect of BSCU1 treatment on peripheral PBMCs in adults and elderly. PCA analysis in adults (A) and elderly (B). Volcano plot in adults (C) and elderly (D). Selected GO terms enriched in adult PBMCs after 4 weeks of BSCU1 treatment compared with baseline (E).
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
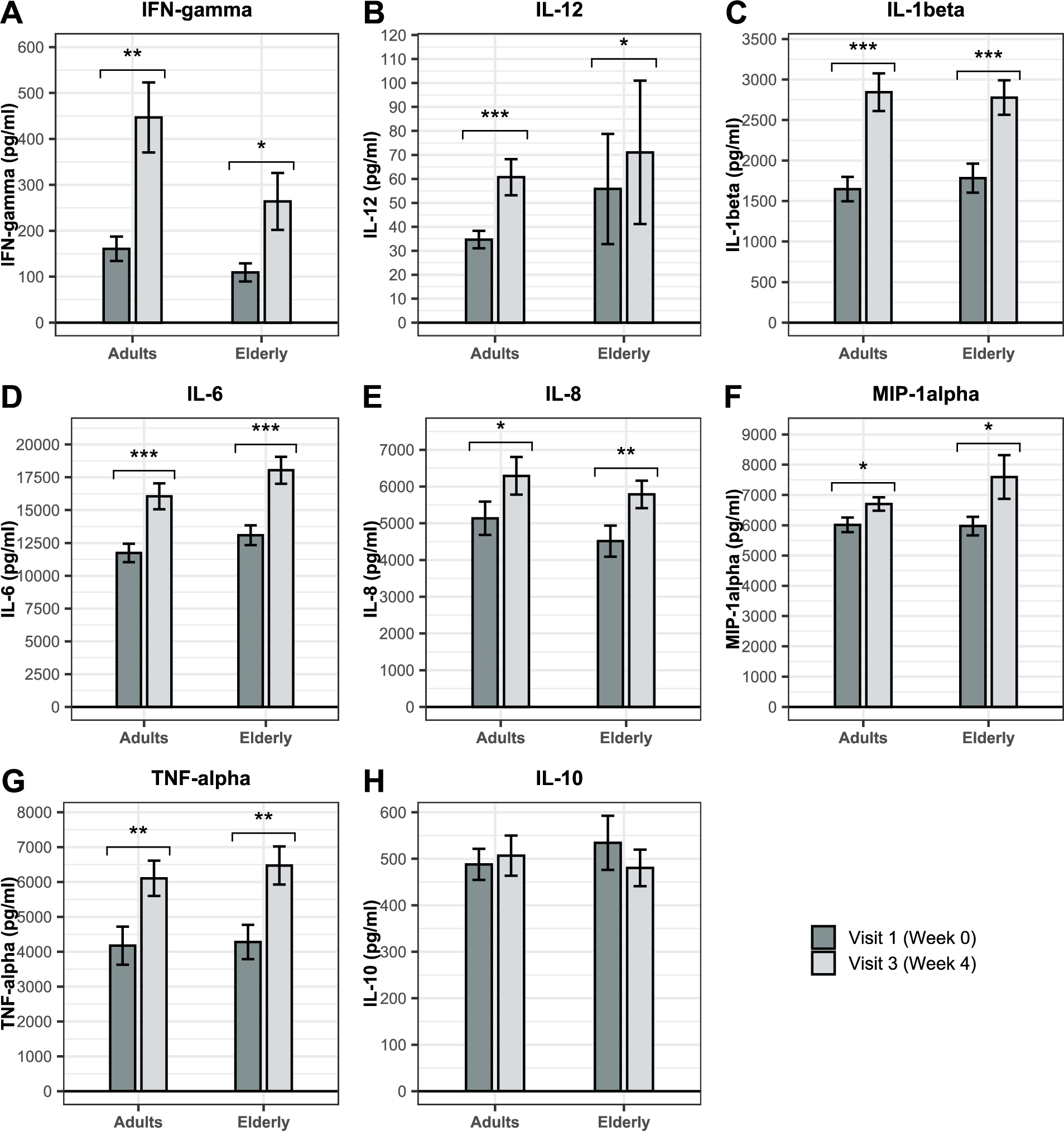
BSCU1 supplementation primes innate immune cells to better respond to an inflammatory challenge
GO term âresponse to type I IFNâ was enriched in the RNAseq dataset after supplementation with BSCU1. Ex vivo cytokine release was measured in whole blood after lipopolysaccharide (LPS) stimulation to indicate the potential or efficiency of immune cells to fight infections. Whole blood obtained at baseline (V1) and after 4 weeks of supplementation (V3) in adults and the elderly was stimulated with LPS for 24 hours. The change in ex vivo cytokines levels between V1 and V3 is presented in Figure 9. A significant increase in the production of IFN-gamma, IL-12, IL-1beta, IL-6, IL-8, MIP-1alpha, and TNF-alpha in both the adult and elderly groups were observed after LPS stimulation. The most affected cytokines were IFN-gamma, IL-1beta, and TNF-alpha, with respective increases of 178%, 72%, and 46% in the adult group and 143%, 56%, and 51% in the elderly. IL-12 was also strongly modulated with a 75% increase in adults and an increase of 27% in the elderly. The change in ex vivo cytokines levels between V1 and V3 was also assessed in the absence of LPS stimulation (Supplementary figure 3). In the absence of LPS stimulation, BSCU1 ingestion did not promote basal secretion of proinflammatory cytokines, although there was an increase in modest basal levels of IL-1beta in adults and MIP-1alpha in the elderly. Thus, supplementation with BSCU1 did not trigger basal inflammation but prepared cells to respond better to an inflammatory challenge.
Effect of BSCU1 on ex vivo cytokine production after stimulation with lipopolysaccharides (LPS). Data show means ± standard error. P-values: * [0.01 to 0.05[; ** [0.001 to 0.01[; *** <0.001.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
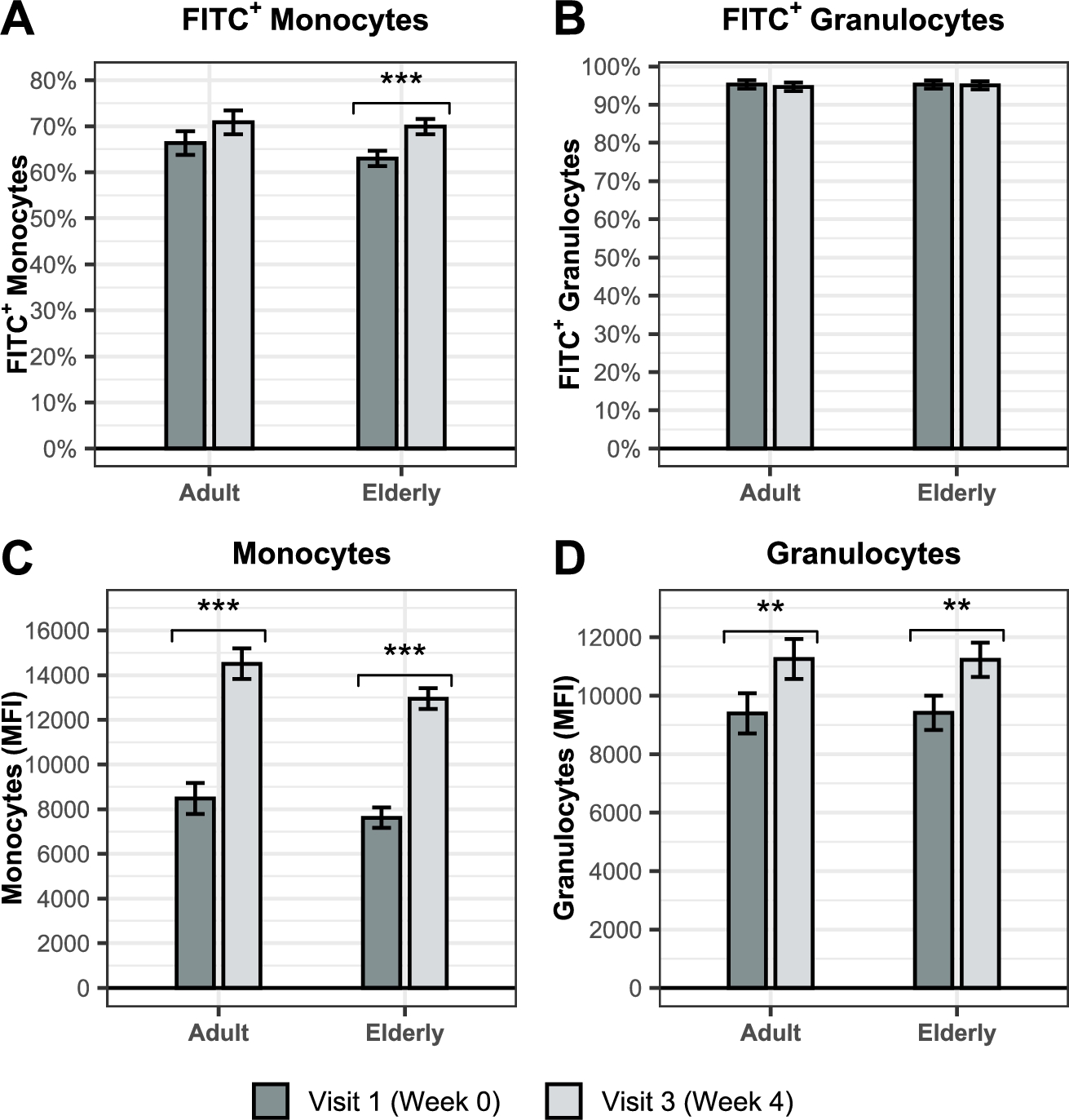
Ingestion of BSCU1 promotes phagocytosis capacity in adults and the elderly
GO term âphagocytosis recognitionâ and âphagocytosis engulfmentâ were enriched in the RNAseq dataset after supplementation with BSCU1. Phagocytosis capacity of granulocytes and monocytes was examined from whole blood collected at baseline and 4 weeks after the first ingestion in adults and elderly using pHrodo⢠Green Escherichia coli BioParticles⢠(Figure 10 and Table 4). The pHrodo Green dye is not fluorescent outside the cell, but fluorescence increases as pH decreases. The percentage of monocytes that phagocyted fluorescently labelled E. coli increased significantly in the elderly with similar trend in adults (Figure 10A). No significant change was observed in the percentage of granulocytes that phagocyted fluorescently labelled E. coli (Figure 10B). The mean fluorescent intensity (MFI) of monocytes, respectively, increased by 75.4% and 69.7% in adults and the elderly (Figure 10C). Increase in mean fluorescent intensity indicates higher pH acidification and thus a faster maturation of phagosomes. A significant difference was also observed for granulocytes with a 20% and 19.2% respective increase in the mean fluorescence intensity in adults and the elderly (Figure 10D). Thus, BSCU1 supplementation promoted the phagocytic capacity of myeloid cells, especially monocytes, in adults and the elderly.
BSCU1 promotes phagocytosis capacity in adults and the elderly. Percentage of monocytes (A) and granulocytes (B) that had engulfed bacteria in adult and elderly. Mean fluorescence intensity of engulfed bacteria in monocytes (C) and granulocytes (D) in adult and elderly. Data show means ± standard error. P-values: ** [0.001 to 0.01[; *** <0.001.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
4 Discussion
Oral supplementation of BSCU1 has been shown previously to increase mucosal sIgA and improved the resistance to common infectious diseases in elderly subjects (Lefevre et al., 2015). The current study has been designed to provide insights into the mechanisms of action through which BSCU1 could modulate the immune system and support protection against infections.
The microbiota was not globally altered by treatment, except for an increased abundance of B. subtilis following dietary supplementation with BSCU1. This is consistent with previous report on probiotics, including spore-forming probiotics (Freedman et al., 2021; Khalesi et al., 2019).
Despite the present study was not able to reproduce previous findings regarding the effect of BSCU1 on sIgA (Lefevre et al., 2015), a trend was observed toward an increase in faecal sIgA concentration in children, adults and the elderly. The interindividual variation of sIgA levels in the current study was higher than expected. As the sample size calculation was performed with a lower variation in sIgA levels, the current study may not have had sufficient power to detect differences in faecal sIgA. The absence of significant effect of BSCU1 on faecal sIgA in the present study could be explained by differences in the study duration, demographic characteristics of the subjects and analytical methods. Indeed, there are no standardised methods known for the measurement of faecal IgA. Differences in faecal sample collection, sIgA extraction, and quantification might partly account for the divergent findings between the studies. Also, the gut microbiota influences intestinal sIgA production and secretion, and in turn, influences faecal sIgA concentrations (Bunker and Bendelac, 2018). Importantly, the gut microbiota varies in different countries and ethnic groups (Deschasaux et al., 2018). It has been shown that individuals with high microbial diversity have relatively stable intestinal ecosystem and are therefore less susceptible to the introduction of new microbes (Xiao et al., 2020). Therefore, differences in the origin of the studied population may also account for the variability of the effect of BSCU1 on sIgA. Indeed, healthy Dutch individuals have been shown to display one of the highest microbial diversities, and are thereby less susceptible to changes in the gut microbiota.
Low-grade inflammation, often referred to as âinflammagingâ when associated with aging, is thought to contribute to many of the chronic diseases of the elderly (Baechle et al., 2023). A variety of stimuli induces inflammaging, including pathogens, endogenous cellular debris and misdirected molecules and nutrients, and the gut microbiota (Franceschi et al., 2018). Indeed, the gut microbiota plays a central role in the development of inflammation due to its ability to release inflammatory mediators. Probiotics can help restore a healthy balance of the gut microbiota and improve barrier function. While there is evidence that certain probiotic strains can reduce low-grade inflammation in the elderly, results are inconsistent and further research is needed (Jukic Peladic et al., 2021). The current study showed that BSCU1 intake lowered basal plasma levels of IL-8, TNF-alpha, and MIP-1alpha. In addition, a significant decrease in IL-10 was also observed in elderly following BSCU1 supplementation. Dysregulation of this cytokine has been suggested to play a role in the susceptibility of aging individuals toward infections (Almanan et al., 2020) and a reduction in IL-10 may therefore be beneficial in this particular age group. No statistically significant change in IFN-gamma level was observed. These results were inconsistent with our previous findings showing increased IFN-gamma levels and no differences in other proinflammatory cytokines (Lefevre et al., 2015). The administration protocol may be responsible for these discrepancies. Because we observed in the current study that BSCU1 probiotic intake primed immune cells to respond better to an inflammatory challenge, it is unlikely that this reduction in low-grade inflammation was related to overall immunosuppression. Rather, alteration of intestinal epithelial cells, e.g. improvement of barrier function, could help reduce the translocation of proinflammatory components of the gut microbiota to the periphery, thereby reducing the inflammatory cascade in the elderly (Untersmayr et al., 2022). It would be interesting to study the markers of gut barrier function in BSCU1-treated individuals. A placebo-controlled study with B. subtilis had previously shown that supplementation for four weeks decreased the number of total T cells, CD8Â + T cells, and Tregs (Freedman et al., 2021). We observed a significant decrease in the frequency of circulating CD4+ T cells and Tregs in the elderly who consumed BSCU1 but no differences in the relative frequency of CD8+ T cells. Previous trial on B. subtilis DE111 reported no effect of the intervention on myeloid cells (Freedman et al., 2021). In this trial, probiotic intervention with BSCU1 was associated with a significant increase in the frequency of monocytes in both the adult and elderly groups, confirming probiotic strain-dependent efficacy.
Effect of BSCU1 on ex vivo phagocytosis
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
We found that BSCU1 ingestion modulated the gene expression program in adult and elderly immune cells, indicating immune stimulation. We did not detect major changes by ingestion of BSCU1 on cytokine production of immune cells without stimulation, suggesting that BSCU1 intake did not cause systemic inflammation. However, probiotic treatment promoted upregulation of genes related to monocytes and their activation. Indeed, Gene Ontology (GO) term analysis showed enrichment in several pathways including âphagocytosis recognitionâ and âphagocytosis engulfmentâ upon BSCU1 intake. In addition, BSCU1 intake promoted pro-inflammatory cytokine secretion upon LPS stimulation, consistent with the GO term enrichment for âresponse to type I IFNâ. The fact that cytokine release was increased following LPS stimulation may indicate a potential beneficial effect on the ability to fight infection. We also found enrichment for the GO terms âphagocytosis recognitionâ and âphagocytosis engulfmentâ in adults who consumed BSCU1. Phagocytosis is an important function of the innate immune system that aims to fight and eliminate microbes (Uribe-Querol and Rosales, 2020). We measured increased phagocytotic activity of monocytes and granulocytes in both the adult and elderly groups that had taken the probiotic. This effect could be, at least partly, mediated by cell envelope molecules such as lipoteichoic acid (LTA). LTA is a major constituent of Gram-positive cell envelopes with immunomodulatory activity, such as macrophages activation (Mueller et al., 2006). VilleÌger et al. (2014) have investigated the strain-level structural diversity of LTA from different Bacillus strains and highlighted the immunostimulatory properties of the LTA from B. subtilis CU1 strain.
5 Conclusions
The spore-forming probiotic bacteria BSCU1 has been shown previously to stimulate immune response and improve the resistance to common infectious diseases in elderly subjects. This clinical study provided further insights into the mechanisms of action of BSCU1 (Figure 11). Ingestion of the probiotic BSCU1 reduced the low-grade chronic inflammation in the elderly and primed the immune system to better respond to microbial challenges. Immune profiling combined with transcriptomic analysis and ex vivo functional assay provided consistent evidence of increased phagocytic capacity following BSCU1 supplementation. The combination of improved responsiveness to microbial challenges and enhanced phagocytic activity are potential mechanisms of action by which BSCU1 may support defence mechanisms against pathogens.
Schematic representation of the effects of BSCU1 on the immune system.
Citation: Beneficial Microbes 15, 6 (2024) ; 10.1163/18762891-bja00028
Corresponding author; e-mail:Â f.mourey@gnosis.lesaffre.com
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.26324023
Materials and methods S1. Selection criteria.
Figure S1. The gating strategy to identify monocytes and DCs.
Figure S2. The gating strategy to identify T cells, B cells, and NK cells.
Figure S3. Effect of BSCU1 on ex vivo cytokine production without stimulation with LPS.
Acknowledgements
We thank study site staff for their cooperation. The participation of the volunteers in this study is greatly appreciated. We would like to thank NIZO laboratory personnel, Dr. Guus Kortman (NIZO) and Dr. Eric Hester (NIZO) for their contributions to the analysis and interpretation of gene expression and microbiota data; Prof. Joost van Neerven (Wageningen University) for the discussions on the immune cell phenotyping results; Joop Pfeil (Estimate Medical Statistics) for performing the statistical analyses and Dr. Anita Hartog as former NIZO colleague involved in the study design.
Authorsâ contribution
Design of the study, FM, JFJ, PS, AK; interpretation of the data, AK, TS, MP, ENM, AC; supervision of statistical analysis, FMa; writing â original draft preparation, ENM, FM; writing â review & editing, JFJ, BR, AD, AK, TS, AC; project administration, PS, FM. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
FM, JFJ, BR, AD, FMa and A.C. are employees of Lesaffre. FM, JFJ, BR, AD, FMa and AC were involved in the design of the study, the interpretation of the data and the writing of the manuscript. PS, AK and TD are employees of NIZO Ltd.
References
Almanan, M., Raynor, J., Ogunsulire, I., Malyshkina, A., Mukherjee, S., Hummel, S.A., Ingram, J.T., Saini, A., Xie, M.M., Alenghat, T., Way, S.S., Deepe, G.S., Divanovic, S., Singh, H., Miraldi, E., Zajac, A.J., Dent, A.L., Hölscher, C., Chougnet, C. and Hildeman, D.A., 2020. IL-10-producing Tfh cells accumulate with age and link inflammation with age-related immune suppression. Science Advances 6: eabb0806. https://doi.org/10.1126/sciadv.abb0806
Baechle, J.J., Chen, N., Makhijani, P., Winer, S., Furman, D. and Winer, D.A., 2023. Chronic inflammation and the hallmarks of aging. Molecular Metabolism 74: 101755. https://doi.org/10.1016/j.molmet.2023.101755
BeÌghin, L., Tims, S., Roelofs, M., RougeÌ, C., Oozeer, R., Rakza, T., Chirico, G., Roeselers, G., Knol, J., RozeÌ, J.C. and Turck, D., 2021. Fermented infant formula (with Bifidobacterium breve C50 and Streptococcus thermophilus O65) with prebiotic oligosaccharides is safe and modulates the gut microbiota towards a microbiota closer to that of breastfed infants. Clinical Nutrition 40: 778-787. https://doi.org/10.1016/j.clnu.2020.07.024
Belkaid, Y. and Harrison, O.J., 2017. Homeostatic immunity and the microbiota. Immunity 46: 562-576. https://doi.org/10.1016/j.immuni.2017.04.008
Budden, K.F., Gellatly, S.L., Wood, D.L.A., Cooper, M.A., Morrison, M., Hugenholtz, P. and Hansbro, P.M., 2017. Emerging pathogenic links between microbiota and the gut-lung axis. Nature Reviews Microbiology 15: 55-63. https://doi.org/10.1038/nrmicro.2016.142
Bunker, J.J. and Bendelac, A., 2018. IgA responses to microbiota. Immunity 49: 211-224. https://doi.org/10.1016/j.immuni.2018.08.011
Chung, H.Y., Kim, D.H., Lee, E.K., Chung, K.W., Chung, S., Lee, B., Seo, A.Y., Chung, J.H., Jung, Y.S., Im, E., Lee, J., Kim, N.D., Choi, Y.J., Im, D.S. and Yu, B.P., 2019. Redefining chronic inflammation in aging and age-related diseases: proposal of the senoinflammation concept. Aging and Disease 10: 367-382. https://doi.org/10.14336/AD.2018.0324
Cutting, S.M., 2011. Bacillus probiotics. Food Microbiology 28: 214-220. https://doi.org/10.1016/j.fm.2010.03.007
Deschasaux, M., Bouter, K.E., Prodan, A., Levin, E., Groen, A.K., Herrema, H., Tremaroli, V., Bakker, G.J., Attaye, I., Pinto-Sietsma, S.-J., van Raalte, D.H., Snijder, M.B., Nicolaou, M., Peters, R., Zwinderman, A.H., Bäckhed, F. and Nieuwdorp, M., 2018. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nature Medicine 24: 1526-1531. https://doi.org/10.1038/s41591-018-0160-1
DudonneÌ, S., Varin, T.V., Forato AnheÌ, F., DubeÌ, P., Roy, D., Pilon, G., Marette, A., Levy, EÌ., Jacquot, C., Urdaci, M. and Desjardins, Y., 2015. Modulatory effects of a cranberry extract co-supplementation with Bacillus subtilis CU1 probiotic on phenolic compounds bioavailability and gut microbiota composition in high-fat diet-fed mice. PharmaNutrition 3: 89-100. https://doi.org/10.1016/j.phanu.2015.04.002
Fan, Y. and Pedersen, O., 2021. Gut microbiota in human metabolic health and disease. Nature Reviews Microbiology 19: 55-71. https://doi.org/10.1038/s41579-020-0433-9
Flint, H.J., Scott, K.P., Louis, P. and Duncan, S.H., 2012. The role of the gut microbiota in nutrition and health. Nature reviews. Gastroenterology and Hepatology 9: 577-589. https://doi.org/10.1038/nrgastro.2012.156
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C. and Santoro, A., 2018. Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nature Reviews Endocrinology 14: 576-590. https://doi.org/10.1038/s41574-018-0059-4
Freedman, K.E., Hill, J.L., Wei, Y., Vazquez, A.R., Grubb, D.S., Trotter, R.E., Wrigley, S.D., Johnson, S.A., Foster, M.T. and Weir, T.L., 2021. Examining the gastrointestinal and immunomodulatory effects of the novel probiotic Bacillus subtilis DE111. International Journal of Molecular Sciences 22: 2453. https://doi.org/10.3390/ijms22052453
Fujiwara, D., Inoue, S., Wakabayashi, H. and Fujii, T., 2004. The anti-allergic effects of lactic acid bacteria are strain dependent and mediated by effects on both Th1/Th2 cytokine expression and balance. International Archives of Allergy and Immunology 135: 205-215. https://doi.org/10.1159/000081305
Gensollen, T., Iyer, S.S., Kasper, D.L. and Blumberg, R.S., 2016. How colonization by microbiota in early life shapes the immune system. Science 352: 539-544. https://doi.org/10.1126/science.aad9378
Hart, A.L., Lammers, K., Brigidi, P., Vitali, B., Rizzello, F., Gionchetti, P., Campieri, M., Kamm, M.A., Knight, S.C. and Stagg, A.J., 2004. Modulation of human dendritic cell phenotype and function by probiotic bacteria. Gut 53: 1602-1609. https://doi.org/10.1136/gut.2003.037325
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology and Hepatology 11: 506-514. https://doi.org/10.1038/nrgastro.2014.66
Jukic Peladic, N., DellâAquila, G., Carrieri, B., Maggio, M., Cherubini, A. and Orlandoni, P., 2021. Potential role of probiotics for inflammaging: a narrative review. Nutrients 13: 2919. https://doi.org/10.3390/nu13092919
Khalesi, S., Bellissimo, N., Vandelanotte, C., Williams, S., Stanley, D. and Irwin, C., 2019. A review of probiotic supplementation in healthy adults: helpful or hype? European Journal of Clinical Nutrition 73: 24-37. https://doi.org/10.1038/s41430-018-0135-9
Kobayashi, S., Ito, A., Okuzaki, D., Onda, H., Yabuta, N., Nagamori, I., Suzuki, K., Hashimoto, H. and Nojima, H., 2008. Expression profiling of PBMC-based diagnostic gene markers isolated from vasculitis patients. DNA Research 15: 253-265. https://doi.org/10.1093/dnares/dsn014
Lefevre, M., Racedo, S.M., Denayrolles, M., Ripert, G., DesfougeÌres, T., Lobach, A.R., Simon, R., PeÌlerin, F., Jüsten, P. and Urdaci, M.C., 2017. Safety assessment of Bacillus subtilis CU1 for use as a probiotic in humans. Regulatory Toxicology and Pharmacology 83: 54-65. https://doi.org/10.1016/j.yrtph.2016.11.010
Lefevre, M., Racedo, S.M., Ripert, G., Housez, B., Cazaubiel, M., Maudet, C., Jüsten, P., Marteau, P. and Urdaci, M.C., 2015. Probiotic strain Bacillus subtilis CU1 stimulates immune system of elderly during common infectious disease period: a randomized, double-blind placebo-controlled study. Immunity and Ageing 12: 24. https://doi.org/10.1186/s12979-015-0051-y
Lehtoranta, L., Latvala, S. and Lehtinen, M.J., 2020. Role of probiotics in stimulating the immune system in viral respiratory tract infections: a narrative review. Nutrients 12: 3163. https://doi.org/10.3390/nu12103163
Levy, M., Kolodziejczyk, A.A., Thaiss, C.A. and Elinav, E., 2017. Dysbiosis and the immune system. Nature Reviews Immunology 17: 219-232. https://doi.org/10.1038/nri.2017.7
Lopetuso, L.R., Scaldaferri, F., Franceschi, F. and Gasbarrini, A., 2016. Bacillus clausii and gut homeostasis: state of the art and future perspectives. Expert Review of Gastroenterology and Hepatology 10: 943-948. https://doi.org/10.1080/17474124.2016.1200465
McAleer, J.P. and Kolls, J.K., 2018. Contributions of the intestinal microbiome in lung immunity. European Journal of Immunology 48: 39-49. https://doi.org/10.1002/eji.201646721
Mohamadzadeh, M., Olson, S., Kalina, W.V., Ruthel, G., Demmin, G.L., Warfield, K.L., Bavari, S. and Klaenhammer, T.R., 2005. Lactobacilli activate human dendritic cells that skew T cells toward T helper 1 polarization. Proceedings of the National Academy of Sciences of the USA 102: 2880-2885. https://doi.org/10.1073/pnas.0500098102
Mueller, M., Stamme, C., Draing, C., Hartung, T., Seydel, U. and Schromm, A.B., 2006. Cell activation of human macrophages by lipoteichoic acid is strongly attenuated by lipopolysaccharide-binding protein. Journal of Biological Chemistry 281: 31448-31456. https://doi.org/10.1074/jbc.M605966200
Perez-Lopez, A., Behnsen, J., Nuccio, S.-P. and Raffatellu, M., 2016. Mucosal immunity to pathogenic intestinal bacteria. Nature Reviews Immunology 16: 135-148. https://doi.org/10.1038/nri.2015.17
Pickard, J.M., Zeng, M.Y., Caruso, R. and NuÌnÌez, G., 2017. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunological Reviews 279: 70-89. https://doi.org/10.1111/imr.12567
Pochard, P., Hammad, H., Ratajczak, C., Charbonnier-Hatzfeld, A.-S., Just, N., Tonnel, A.-B. and Pestel, J., 2005. Direct regulatory immune activity of lactic acid bacteria on Der p 1-pulsed dendritic cells from allergic patients. Journal of Allergy and Clinical Immunology 116: 198-204. https://doi.org/10.1016/j.jaci.2005.02.037
Sharon, G., Sampson, T.R., Geschwind, D.H. and Mazmanian, S.K., 2016. The central nervous system and the gut microbiome. Cell 167: 915-932. https://doi.org/10.1016/j.cell.2016.10.027
Trizzino, M., Zucco, A., Deliard, S., Wang, F., Barbieri, E., Veglia, F., Gabrilovich, D. and Gardini, A., 2021. EGR1 is a gatekeeper of inflammatory enhancers in human macrophages. Science Advances 7: aaz8836. https://doi.org/10.1126/sciadv.aaz8836
Untersmayr, E., Brandt, A., Koidl, L. and Bergheim, I., 2022. The intestinal barrier dysfunction as driving factor of inflammaging. Nutrients 14: 949. https://doi.org/10.3390/nu14050949
Urdaci, M.C., Bressollier, P. and Pinchuk, I., 2004. Bacillus clausii probiotic strains: antimicrobial and immunomodulatory activities. Journal of Clinical Gastroenterology 38: S86-S90. https://doi.org/10.1097/01.mcg.0000128925.06662.69.
Uribe-Querol, E. and Rosales, C., 2020. Phagocytosis: our current understanding of a universal biological process. Frontiers in Immunology 11: 1066. https://doi.org/10.3389/fimmu.2020.01066
VilleÌger, R., Saad, N., Grenier, K., Falourd, X., Foucat, L., Urdaci, M.C., Bressollier, P. and Ouk, T.-S., 2014. Characterization of lipoteichoic acid structures from three probiotic Bacillus strains: involvement of D-alanine in their biological activity. Antonie van Leeuwenhoek 106: 693-706. https://doi.org/10.1007/s10482-014-0239-8
Xiao, Y., Angulo, M.T., Lao, S., Weiss, S.T. and Liu, Y.-Y., 2020. An ecological framework to understand the efficacy of fecal microbiota transplantation. Nature Communications 11: 3329. https://doi.org/10.1038/s41467-020-17180-x
Zeuthen, L.H., Christensen, H.R. and Frøkiaer, H., 2006. Lactic acid bacteria inducing a weak interleukin-12 and tumor necrosis factor alpha response in human dendritic cells inhibit strongly stimulating lactic acid bacteria but act synergistically with gram-negative bacteria. Clinical and Vaccine Immunology 13: 365-375. https://doi.org/10.1128/CVI.13.3.365-375.2006
Zhang, H.-L., Li, W.-S., Xu, D.-N., Zheng, W.-W., Liu, Y., Chen, J., Qiu, Z.-B., Dorfman, R.G., Zhang, J. and Liu, J., 2016. Mucosa-reparing and microbiota-balancing therapeutic effect of Bacillus subtilis alleviates dextrate sulfate sodium-induced ulcerative colitis in mice. Experimental and Therapeutic Medicine 12: 2554-2562. https://doi.org/10.3892/etm.2016.3686





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}