Abstract
The gut microbiota has been proposed to grant the athlete a metabolic advantage that might be key when optimising performance. While a taxonomic core set of microorganisms characterising the athlete’s gut microbiota has not been delineated, some compositional features might be associated with improved metabolic efficiency, which appears to be driven by the production of bacterial metabolites, such as short-chain fatty acids. Not only long-term exercise but also dietary patterns associated with high-level sports practice contribute to this microbial environment, yet isolating the impact of individual dietary components is challenging. The present review synthetises the available evidence on the compositional aspects of the athlete’s gut microbiota, discusses mechanisms involved in the bidirectional association between exercise and the gut environment, and evaluates the role of athletes’ diet in this interplay. Additionally, a practical approach to indicators commonly reported in metagenomic and metabolomic analyses is provided to explore how these insights can translate to support dietary protocols.
1 Introduction
The human microbiota is the collection of microorganisms that inhabit the human body and mostly comprises bacteria (≈99%) (Qin et al., 2010), but also fungi, archaea, eukaryotic viruses, phages, and prophages (Scarpellini et al., 2015). In quantitative terms, microbiota cells have been estimated to equal human body cell number, accounting to 0.2 kg total mass for a 70 kg average person (Sender et al., 2016). Microbes distribute ubiquitously to almost all body surfaces and fluids, and are particularly important in mucosal membrane surfaces, such as the gastrointestinal tract. The gut harbours the most densely populated ecosystem studied to date, consisting of a diverse and dynamic microbial community whose composition is mainly defined during early life, and is subject to a process of maturation throughout adulthood to finally decline (at least in terms of diversity) as we age (Martino et al., 2022).
Driven by the advances in culture-independent molecular assays, a growing body of literature positions the gut microbiota as a crucial contributor to metabolic health (Lynch and Pedersen, 2016). In this sense, variations in microbial indices and differential abundance of certain bacteria phyla, or even specific species (explained in Textbox 1), have been consistently linked to metabolic disorders, such as obesity, type 2 diabetes, and cardiovascular disease (Jie et al., 2017; Turnbaugh et al., 2009; Wu et al., 2010). However, attempts to identify a taxonomic core set of microbes defining what can be considered a ‘healthy gut microbiota’ have been shown unfruitful due to the elevated intra- and inter-individual variability observed (Lozupone et al., 2012). Notwithstanding, we nowadays recognise the role that the gut microbiota plays in a wide repertoire of physiological processes including energy metabolism and nutrient uptake (Krajmalnik-Brown et al., 2012), gastrointestinal health (Aziz et al., 2013), and immune response (Yoo et al., 2020), which in turn can impact muscle functionality and physical performance.

Compositional indices used in faecal metagenomics
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
Optimising exercise performance is key for elite athletes, as subtle performance differences can be decisive in achieving competition goals. Consequently, authors have put a spotlight on the gut microbiota as a factor that might grant the athlete a metabolic or physiological advantage (Mohr et al., 2020). However, to date it remains unclear how these insights can translate to sports practice. Athletes, trainers, and sports physicians are deciding on dietary protocols aimed to optimise performance and recovery, without considering the impact that these practices may have on the gut microbiota. Similarly, sports professionals may turn to faecal analyses to explore gut microbial and metabolomic profiles to identify potential causes underlying gastrointestinal symptoms, but guidance on how to interpret or incorporate the findings to their protocols is lacking. Therefore, the present work aims to provide a complete picture of the scientific evidence pertaining to the characteristics of the gut microbiota of athletes, the mechanisms involved in the interplay between exercise and the gastrointestinal ecosystem, and the role that athletes’ dietary strategies might play in the gut homeostasis. Additionally, the present work also provides some guidance on microbial community dynamics and biomarkers commonly reported in metagenomic and metabolomic analyses to explore how these insights can translate to sports practice and dietary protocols.
Impact of physical activity, fitness, and exercise on the composition of the gut microbiota of non-athlete adults. Figure created using BioRender.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
2 The athlete’s gut microbiota profile
Once an adult-like profile of the gut microbiota is established, wide genetic, epigenetic, and environmental cues can influence both its composition and functionality, among which physical activity and exercise have gained considerable interest. Cross-sectional analyses of faecal samples from healthy non-athlete adults have consistently shown that physical activity patterns are insufficient to drive significant changes in faecal diversity indices in these populations, with only shifts in relative abundance of certain bacterial taxa being reported (Bressa et al., 2017; Castellanos et al., 2019; Galle et al., 2020; Whisner et al., 2018; Xu et al., 2022). Longitudinal studies with larger sample sizes have yielded mixed results, which were likely to be impacted by differences in body composition and dietary metrics as well as the tools used to assess physical activity (Holzhausen et al., 2022; Shah et al., 2023). In the same vein, short-term exercise interventions have failed to produce notable changes in diversity metrics in lean, and overweight individuals (Allen et al., 2018; Cronin et al., 2018; Moitinho-Silva et al., 2021; Resende et al., 2021), with some authors suggesting that aerobic exercise, contrary to resistance training, might produce subtle and reversible spatial shifts in the composition of the gut microbiota (Bycura et al., 2021). On the other hand, cardiorespiratory fitness measured by maximal oxygen consumption (VO2max) or peak oxygen consumption (VO2peak) has been shown to associate with higher α-diversity, increased Firmicutes:Bacteroidetes ratio (F:B) (recently renamed to Bacillota:Bacteroidota (Oren and Garrity, 2021). Hereinafter the older terminology will be used to facilitate the reading and understanding in the context of the reviewed literature), and expanded relative abundance of bacteria like Roseburia spp. and Eubacterium rectale, which are typically considered beneficial in several trials (Durk et al., 2019; Estaki et al., 2016; Resende et al., 2021; Yang et al., 2017). Albeit based on a limited number of studies, cardiorespiratory fitness, resulting from long-term consistent physical activity and exercise practice, appears to act as a sufficiently strong stimulus to exert changes in high-level compositional indices of the gut microbiota in non-athlete individuals. A schematic representation of these key points is presented in Figure 1.
On the other hand, athletes represent an ideal cohort for studying the effects of long-term exercise practice on human physiology and, particularly, on the gut microbiota. Specifically high-level athletes undergo severe metabolic and physiological adaptations aimed at improving muscle strength, enhancing aerobic endurance, optimising energy expenditure and heat mitigation, which, along with dietary strategies designed to support the needs of high-level sports practice, may contribute to shaping a characteristic gut community compositional profile.
Evidence on the role of the gut microbiota in athletic performance
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
It would then seem reasonable to expect notable differences in community composition arising from comparisons between athletes and sedentary individuals. In a pioneering study published in 2014, Clarke et al., aimed to explore differences in the gut microbiota profile between 40 elite rugby players and two groups of sedentary controls with high (n = 23) and low (n = 23) body mass index (BMI), respectively (Clarke et al., 2014). Data on dietary intake and physical activity levels were reported through validated questionnaires, and faecal samples were collected for 16S rRNA sequencing analyses (Clarke et al., 2014). As revealed by the analyses, several indices of α-diversity were higher in the elite athletes compared with both control groups, while no difference was observed between control groups (Clarke et al., 2014). Similarly, unweighted (qualitative) UniFrac distances were shown to effectively separate athlete and control microbiomes (Clarke et al., 2014). Further, several differences in bacterial taxa were reported, including less enriched Bacteroidetes and higher relative abundance of Akkermansiaceae (Akkermansia) in the elite athlete group (Clarke et al., 2014). Notably, lower abundance of Akkermansia was observed in the high-BMI group compared to the low-BMI control group (Clarke et al., 2014). While elite athletes had a mean BMI close to that of the high-BMI control group (29.1 ± 3.0 vs 31.2 ± 3.0), clearly the percentage of body fat was lower and lean body mass was higher in the elite athlete group (Clarke et al., 2014). This highlights the importance of measuring body composition outcomes (i.e. using bioimpedance), which may be more informative than merely reporting BMI when exploring sport-specific situations. In an ensuing study, Kulecka et al. (2020) conducted 16S rRNA sequencing analyses of the faecal microbiome of 14 marathon runners, 11 elite cross-country skiers, and 46 healthy controls. Study results showed higher richness and diversity (Chao1 and Simpson indices) along with elevated F:B in athletes compared with control individuals, which were more pronounced in cross-country skiers (Kulecka et al., 2020). Lower abundance of Bacteroidetes and higher abundance of Prevotella were observed in both athlete groups while increased abundance of Veilonella was shown in marathon runners (Kulecka et al., 2020). Lastly, in a recent report from the same group, the microbiome of professional athletes (marathon runners, cross-country skiers, and soccer players) was shown to be more diverse (Shannon) than that of e-sports players, characterised by poor physical activity habits, and display differences in the relative abundance of several beneficial bacteria (i.e. Akkermansia muciniphila and Roseburia hominis) (Kulecka et al., 2023).
In the light of these studies, it might be suggested that athletes have a more diverse gut microbiota that might contain a higher abundance of beneficial bacterial taxa (some of them with purported ergogenic properties (example presented in Textbox 2)), compared to that of sedentary individuals. Notwithstanding, there are inconsistent findings that have been reported in the literature. For example, in a study aimed to evaluate the impact of endurance and strength exercise on the microbiota of inactive individuals, there was an additional group consisting of 12 elite cyclists and triathletes that was recruited for comparison (Moitinho-Silva et al., 2021). No differences in α- or β-diversity indices were observed between physically inactive individuals and elite athletes (Moitinho-Silva et al., 2021). Only amplicon sequence variants (ASVs) belonging to Coprococcus, Parasutterella and the family Ruminococcaceae (Oscillospiraceae) were shown to be more abundant in elite athletes while those belonging to Dialister, Odoribacter and Phascolarctobacterium were reported to be increased in inactive individuals (Moitinho-Silva et al., 2021). Similarly, Jang et al. (2019) evaluated the faecal bacterial microbiome of 15 bodybuilders, 15 distance runners, and 15 sedentary controls observing no differences in the diversity of the microbiota of enrolled. Along with differences in the methods used to monitoring physical activity and dietary patterns, sample size limitations might explain these contradictory findings.
Elite athletes face high levels of stress derived from in-season competition demands and expectations (Pensgaard and Ursin, 1998). During exercise, a bidirectional association between the autonomous nervous system and the gastrointestinal tract exists, which takes part in the stress response and is proposed to be partially mediated by the release of gut hormones and gut microbiota-derived molecules (Clark and Mach, 2016). In addition, the adoption of extreme dietary patterns and adherence to exhaustive exercise protocols by elite athletes might lead to differences in gut microbial compositions compared to less trained individuals, or across different competition levels.
Only a few studies have reported comparisons between elite athletes and healthy physically active non-athlete individuals. In a study conducted by Hintikka et al. (2022) authors failed to identify substantial compositional differences in the gut microbiota of elite cross-country skiers and physically active control students (non-athlete group) (Hintikka et al., 2022). Nonetheless, athletes were observed to have lower phylogenetic diversity and relative abundance of mucin-degrading microbial taxa, including the Akkermansia genus, and these changes were suggested to be linked to the competitive season demands. Importantly, authors did not collect data on dietary intake and participants were not matched by body composition, which compromises reaching solid conclusions (Hintikka et al., 2022). In the abovementioned study conducted by Kulecka et al. (2023) a further comparison group consisting of active physical education students was explored. Aligned with comparisons with e-sports players, authors identified marked differences in bacterial abundance at the species level and β-diversity between professional athletes and students, yet no difference in α-diversity was observed (Kulecka et al., 2023). In another study, Xu et al. (2022) reported no differences in α- and β-diversity between track and field sports athletes and young individuals classified based on physical activity levels. While several differences in relative abundance of specific taxa were noted, these were rendered insignificant when adjusting for dietary intake (Xu et al., 2022). Results from these studies further support the need of controlling diet, body composition, and sampling time to enable comparisons between gut compositional profiles.
Regarding studies enrolling athletes competing at different levels, Han et al. (2020) evaluated the gut microbiome of 19 female rowing athletes stratified into elite rowers, young elite rowers, and young non-elite rowers. Authors reported that microbial α-diversity (Shannon and Simpson indices) was richer in elite athletes compared to non-elite (Han et al., 2020). Similarly, compositional spatial differences were observed between elite and non-elite groups as revealed by the analysis of unweighted UniFrac and Abund-Jaccard distance metrics (Han et al., 2020). Further, the relative abundance of Firmicutes and Proteobacteria (recently renamed to Pseudomonadota (Oren and Garrity, 2021)) in elite athletes was higher than that of the non-elite group whereas the opposite was true for Bacteroidetes (Han et al., 2020). The authors reported a phenotypic composition consisting of higher abundance of oxidative stress tolerant microbes in enrolled elite athletes, which was speculated to improve performance in competitive sports such as rowing (Han et al., 2020). When analysing dominant enterotypes, elite athletes displayed a clearly differentiating enterotype characterised by predominance of bacteria belonging to the Ruminococcaceae family, which was strongly associated with physical features, dietary intake, and sport-related aspects (Han et al., 2020). Overall, this study provided extensive evidence on existing taxonomical differences in the gut microbiota of elite and non-elite athletes. Similar results were observed in another cross-sectional study, where 28 martial arts athletes were analysed and classified according to qualifications obtained in national competitions (Liang et al., 2019). Data on frequency of food consumption over the past three months were reported and faecal samples were collected for 16S rRNA sequencing (Liang et al., 2019). Exercise load and microbial α-diversity indices (Shannon and Simpson) were significantly higher in the elite athletes (Liang et al., 2019). A trend towards different microbial structures across groups (β-diversity measurements) was observed (Liang et al., 2019). A total of 10 microbial taxa, including three families and seven genera were differentially abundant in higher-level vs lower-level athletes. Specifically, the Porphyromonadaceae and Acidaminococcaceae families, and the Parabacteroides, Phascolarctobacterium, Bilophila and Oscillibacter genera were enriched while Veillonellaceae (formerly Acidaminococcaceae), and the genera Allisonella, Citrobacter, and Megasphaera were shown to be lower in abundance in elite athletes compared to athletes classified to lower competitive levels (Liang et al., 2019). Notably, the abundance of Parabacteroides was strongly correlated with exercise load, particularly in elite athletes (Liang et al., 2019). Lastly, in a previous pilot study conducted by Petersen et al. (2017) authors analysed faecal samples of 33 cyclists using shotgun sequencing, and observed that professional cyclists were more likely to be included in a taxonomic cluster characterised by higher richness and diversity compared with amateur cyclists, although correlations were not statistically significant due to the pilot nature of the study. Of note, exercise load was associated with higher abundance of Prevotella, which often characterises diets rich in complex carbohydrates (Petersen et al., 2017).
Although based on a limited number of studies, it might be suggested that the gut microbiota of elite athletes has improved compositional features namely higher richness, diversity, relative abundance of beneficial bacteria taxa, and metabolic efficiency (discussed later on) compared to that of lower category athletes. However, there are several additional caveats to consider when evaluating comparisons between the gut microbial profiles of elite athletes and less trained individuals.
Firstly, not only training load but also type of exercise may differentially impact the gut microbiota in elite athletes, yet this has been insufficiently studied. O’Donovan et al. (2020) recruited 37 elite athletes who competed across 16 different sports disciplines, collected food frequency data, and faecal samples were analysed using Illumina shotgun sequencing. Sports were classified based on their dynamic or static components and, despite no observed significant variations in dietary intake across groups, there were discriminatory species found for most of the exercise categories evaluated (O’Donovan et al., 2020). In the same vein, a recent multi-cohort study involving 543 faecal samples from aerobics, wrestling, and rowing athletes reported substantial differences in the gut microbial profiles between the sports disciplines explored (Li et al., 2023). For instance, rowing female athletes displayed higher α-diversity (Shannon) than aerobics female athletes, and their overall profiles at the genus level were clearly differentiated based on Bray-Curtis dissimilarities (Li et al., 2023). The authors also observed network clusters and microbial subgroups enriched in athletes of different sports disciplines, and comprehensibly reported sex- and intensity-dependent interactions, among different associations with diet, performance, and immune cell counts (Li et al., 2023). Based on these findings, the authors argued that the athlete’s gut microbial composition and diversity is sport-specific (Li et al., 2023).
Characteristics of the elite athlete gut microbiota and potential confounders. Figure created using BioRender.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
Secondly, most evidence on a direct role of the gut microbiota in exercise performance relies on pre-clinical research (discussed in Textbox 2) and intervention trials (disclosed in following sections), with an absence of longitudinal observational research (Hintikka et al., 2022; Viciani et al., 2022). A scheme summarising the potential main features of the elite athlete gut microbiota versus physically active individuals, lower category athletes mentioned in the studies above, and factors that might confound these observations is presented in Figure 2.
3 Bidirectional interplay between the skeletal muscle and the gut microbiota
Compositional differences in the gut microbiota might consequently lead to metabolic repercussions denoted by differential expression of functional pathways, or changes in faecal (or circulating) levels of microbial metabolites. Several studies cited above have supplemented metagenomics with metabolomic or metatranscriptomic techniques to determine if compositional changes in the athlete’s gut microbiota reflect changes in functional or metabolic pathways (Barton et al., 2018, 2021; Cronin et al., 2018; O’Donovan et al., 2020; Petersen et al., 2017; Tabone et al., 2021). For instance, metabolomic changes in energy, amino acid, and carbohydrate metabolism have been repeatedly proposed to improve metabolic efficiency in athletes (Barton et al., 2018; O’Donovan et al., 2020; Petersen et al., 2017; Tabone et al., 2021). Similarly, different authors have opted for quantifying targeted microbial metabolites with potential effects on gut homeostasis and metabolic health to support the information provided in compositional assessments (Allen et al., 2018; Estaki et al., 2016). On the other hand, a growing body of literature supports an endocrine function of the skeletal muscle denoted by the production of myokines that are thought to reach distant locations to exert diverse effects (Delezie and Handschin, 2018; Pedersen and Febbraio, 2008). This molecular interplay between the skeletal muscle and the gut microbiota has been termed the ‘gut-muscle axis’ (Grosicki et al., 2018) and might be key when exploring associations between the gut microbiota and exercise.
By far, the most extensively studied products of the gut microbiota metabolism are short-chain fatty acids (SCFAs). SCFAs are a set of volatile fatty acids derived from the degradation of dietary fibre by the gut microbiota as well as from different routes involving succinate and lactate metabolism, amino acids, etc. (Martin-Gallausiaux et al., 2021; Rios-Covian et al., 2016) Acetate, propionate, and butyrate are the SCFAs produced in the largest quantities in the colon, and play important local roles in promoting nutrient absorption and maintaining the gut barrier (Rios-Covian et al., 2016). SCFAs are partially absorbed through the portal vein and incorporated into carbohydrate and lipid metabolic routes, thus serving as substrates for energy metabolism and lipid biosynthesis (Rios-Covian et al., 2016). After colonocyte and liver metabolism, some SCFAs remnants reach the systemic circulation, thus becoming bioavailable (Boets et al., 2017). Important signalling functions have been attributed to these bacterial products, which have been speculated to modulate diverse aspects related to gut peptide secretion, inflammatory and immune responses, body energy homeostasis, etc. (Kasubuchi et al., 2015) Signalling actions are thought to be mediated by the activation of the G-protein-coupled receptors GPR43 (FFAR2) and GPR41 (FFAR3), which are also expressed on skeletal muscle (Han et al., 2014; Nilsson et al., 2003).
As commented, differences in SCFA faecal content, pathways associated to SCFA synthesis, and populations of SCFA-producing bacteria between different physical activity behaviours and athletic levels have been reported in several abovementioned studies (Barton et al., 2018; Clarke et al., 2014; Estaki et al., 2016). However, direct evidence on the actions of SCFAs on muscle mass, function, and exercise performance has been largely obtained using in vitro and in vivo research models due to methodological challenges applicable to research in human subjects. Experiments conducted in microbiota-depleted mice have observed improvements in exercise capacity and muscle function in animals treated with SCFAs (Lahiri et al., 2019; Okamoto et al., 2019). Briefly, different SCFAs might improve short-term glycogen replenishment (Fushimi and Sato, 2005; Fushimi et al., 2001; Nakao et al., 2001; Sakakibara et al., 2006) through transient inhibition of glycolysis, enhanced GLUT4 expression, and improved insulin-mediated glucose uptake on skeletal muscle (Han et al., 2014; Maruta et al., 2016; Yamashita et al., 2009). In addition, SCFAs may play important roles in skeletal muscle lipid metabolism including enhanced fatty acid uptake and oxidation, increased energy expenditure, and reduced muscle lipid deposition (Den Besten et al., 2015; Gao et al., 2009; Liu et al., 2019; Maruta et al., 2016; Pan et al., 2015; Walsh et al., 2015), which, in combination with mentioned effects on carbohydrate metabolism, suggests a role of SCFAs in improving metabolic efficiency to support endurance capacity. Further, some preclinical evidence suggests that SCFAs might improve or preserve lean mass (Gao et al., 2009; Walsh et al., 2015). Studies conducted in elderly adults have observed associations between higher abundance of SCFA-producing bacteria, enhanced capacity for butyrate synthesis, increased faecal butyrate content, and higher skeletal muscle mass (Han et al., 2022; Lv et al., 2021).
Mechanisms involved in the potential role of short-chain fatty acids in exercise recovery and performance. AMPK = AMP-activated protein kinase; FFAR = free-fatty acid receptor; GLP-1 = glucagon-like peptide 1; HDAC = histone deacetylase, PPAR = peroxisome proliferator-activated receptor; SCFA = short-chain fatty acid. Figure created using BioRender.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
Exact mechanisms underlying effects on muscle mass have not been sufficiently explored. On the other hand, mechanisms by which SCFAs may mediate effects on improved energy utilisation are multiple and are proposed to include activation of AMP-activated protein kinase (AMPK) (Sakakibara et al., 2006; Yamashita et al., 2009), activation of peroxisome proliferator-activated receptors (PPARs) (Gao et al., 2009; Yamashita et al., 2009), and inhibition of histone deacetylases (HDACs) (Den Besten et al., 2015; Walsh et al., 2015). In particular, activation of the master regulator AMPK in muscle leads to the activation of catabolic pathways whose effects resemble those produced by SCFAs (Hu et al., 2010). Also, peroxisome proliferator-activated receptor gamma coactivator 1α (PGC1α) is a well-described downstream target of AMPK and is known to control mitochondrial biogenesis, a process crucial to cellular energy metabolism, and is thought to be involved in the activation of PPARs (He et al., 2020), which are closely related to lipid and carbohydrate metabolism (Walczak and Tontonoz, 2002). Regarding HDAC, effects on cell function are complex and require further understanding (He et al., 2020). SCFAs exert additional indirect effects that might contribute to muscle function, including enhanced glycogen synthesis resulting from hepatic metabolism to support fuel needs, increased blood flow to the muscle, improved gastrointestinal barrier status (thus improving nutrient availability), and induced release of gastrointestinal peptides, which overall can indirectly improve muscle metabolism, and potentially influence exercise performance and recovery (Frampton et al., 2020). A visual representation of the potential repercussions of SCFAs effects on exercise performance and recovery is presented in Figure 3.
Other products of gut bacteria metabolism have been proposed to modulate muscle function and impact exercise. Lipopolysaccharide (LPS) is an endotoxin produced by most gram-negative bacteria as part of their outer membrane structure, which, once present in the bloodstream, interacts with the pattern recognition toll-like receptor 4 (TLR4) and with CD14 to activate a pro-inflammatory signalling cascade (Ciesielska et al., 2021). In settings of infection, this process aims to eradicate invading bacteria (Ciesielska et al., 2021). However, taxonomic shifts towards decreased abundance of beneficial species purported to facilitate the maintenance of the gastrointestinal barrier, and increased abundance of pathogenic or ‘unhealthy’ gram-negative bacteria might lead to increased presence of LPS and leakage into systemic circulation (Kinashi and Hase, 2021). Low-grade levels of LPS have been linked to gastrointestinal symptoms during exercise, and higher circulating LPS levels are speculated to negatively impact recovery from exercise (Roberts et al., 2016). Sedentarism and low cardiorespiratory fitness have been shown to be associated with higher LPS levels and enhanced LPS biosynthesis pathways compared to exercise training and better cardiorespiratory fitness (Estaki et al., 2016; Lira et al., 2010). Besides data on physical activity behaviours, scarce information on the effect of LPS levels on exercise-related outcomes is available.
A promising set of compounds produced by gut microbial metabolism are secondary bile acids (Mancin et al., 2022b). Although classically considered as a product of cholesterol metabolism merely responsible for lipid absorption, endocrine functions have been attributed to these molecules (Mancin et al., 2022b). Briefly, primary bile acids are metabolised into secondary bile acids by the gut bacteria and a small proportion of the total bile acid pool reaches systemic circulation. These circulating secondary bile acids elicit signalling effects at distant locations, which are thought to be mediated by the activation of nuclear receptors such as the farnesoid X receptor (FXR) and the G-protein-couple receptor ‘TGR5’ (Mancin et al., 2022b). Only a few pre-clinical studies have investigated the role of secondary bile acids in muscle function and, therefore, any association with exercise can only be speculated upon. The action of bile acids on the FXR receptor (mainly expressed on ileum and liver (Panzitt and Wagner, 2021)) leads to the production of FGF19 in humans, which in turn appears to modulate skeletal muscle mass (Qiu et al., 2021). Also, TGR5 is demonstrated to be expressed on skeletal muscle where it is targeted by bile acids, and it is thought to promote hypertrophy and improve muscle function upon activation (Sasaki et al., 2018). The role of bile acids in muscle metabolism is a novel avenue to explore, and further research is needed to delineate their potential effects on exercise.
Besides secondary bile acids, emerging evidence suggests that other microbiota metabolites might mediate effects within the gut-muscle axis. For instance, bacterial metabolism of amino acids such as branched-chain amino acids (BCAAs) and aromatic amino acids (AAAs) leads to the production of fermentation products including branched-chain fatty acids (BCFAs), phenolic, indolic, and p-cresolic compounds among others (including SCFAs), which are speculated to exert diverse metabolic effects on the host (Hughes and Holscher, 2021; Zhai et al., 2021). On the other hand, some bacteria are known to participate in proteolytic processes thus modulating amino acid availability and subsequently muscle anabolism (Lin et al., 2017). Additionally, the stress response to exercise might be influenced by bacterial mediated amino acid metabolism through the production of compounds involved in the so-called ‘gut-brain axis’. The gut microbiota is known to modulate the synthesis of neurotransmitters and hormones such as catecholamines (i.e. serotonin, norepinephrine, noradrenaline, dopamine), γ-aminobutyric acid (GABA), etc. (Clark and Mach, 2016; Lin et al., 2017) Further, several bacteria belonging to the Bifidobacterium and the former Lactobacillus genera such as Limosilactobacillus reuteri strains are claimed to play a role in the de novo biosynthesis of vitamin K and most B-group vitamins with these being absorbed in the colon, thus adding to the pool of these micronutrients that contribute to the normal function of the muscle (LeBlanc et al., 2013). Interestingly, Magnúsdóttir et al. (2015) estimated that the gut microbiota might produce up to 86% of the dietary reference intake (DRI) of pyridoxine, 31% of cobalamin, 37% of folate, and 27% of niacin, among other B-group vitamins, yet these estimates did not account for potential cross-feeding processes. Some of these vitamins, such as folate and cobalamin, play important roles in red cell production (Peeling et al., 2023), which is particularly interesting for athletes to support oxygen delivery. Further studies are needed for a deeper understanding of the microbial contribution to vitamin pools in the host. Lastly, the gut microbiota might also contribute to elevations in antioxidant enzyme levels, produce bioactive compounds from dietary ingredients (i.e. urolithins derived from polyphenol metabolism), and interact with the endocannabinoid system (which is speculated to modulate exercise adaptation processes (Schonke et al., 2020)), among other less-explored activities (Przewlocka et al., 2020). The science around these mechanisms is still young and further research is needed to evaluate how these processes can impact exercise performance and recovery.
Since the association between the gut microbiota and the skeletal muscle is considered bidirectional, a reciprocal action of the skeletal muscle on the composition of the gut microbiota would be expected. However, although the skeletal muscle can act as an endocrine organ mediated through the production of myokines such as interleukins (i.e. interleukin (IL)-6, IL-8, IL-15), irisin, and β-aminoisobutyric acid (BAIBA), to name a few (Delezie and Handschin, 2018; Pedersen and Febbraio, 2008), the main effects of the skeletal muscle on the gut microbiota seem to be largely attributed to exercise practice. As concluded in the abovementioned studies, different types of exercise and intensity might differentially modulate populations and functionality of gut microbes (O’Donovan et al., 2020) and, therefore, the mechanisms involved can vary accordingly and are not fully understood. Mild exercise is known to enhance gut transit of intraluminal gas (Dainese et al., 2004) and reduce gut transit time (Oettlé, 1991). Increases in transit times have been shown to result in a decrease in biomass and diversity in distal compartments, being therefore considered a key aspect that modulates the composition of the gut microbiota (Tottey et al., 2017). Long-term moderate intensity exercise has been linked to improved intestinal permeability, increased expression of antimicrobial peptides (Luo et al., 2014), and reduced release of LPS into systemic circulation (Li et al., 2021). On the contrary, sustained vigorous or strenuous exercise redistributes blood flow to exercising muscles, lungs and heart leading to local splanchnic ischemia, which subsequently results in barrier injury and increased gut permeability (Van Wijck et al., 2011). In turn, these deleterious effects of strenuous exercise on gut health can lead to gastrointestinal discomfort and increased risk of endotoxemia, which are exacerbated in hot environments (Chantler et al., 2021). The flow of cytokines and LPS to the blood stream due to increased permeability is termed ‘leaky gut’, a phenomenon that athletes must avoid as it leads to impaired nutrient absorption, diarrhoea, flatulence, and pain (Ribeiro et al., 2021). Other mechanisms through which strenuous exercise is suggested to modulate the gut microbiota include activation of the hypothalamus-pituitary-adrenal (HPA) axis, weight loss, increased oxidative stress and inflammation, and changes in bile acid profiles (Cerda et al., 2016).
4 Role of nutrition and dietary interventions in the gut microbiota of athletes
Nutritional and dietary strategies aimed to optimise sports performance and accelerate recovery from exercise have been subject to extensive research (Thomas et al., 2016), albeit often without considering the impact that these strategies may have on the gut microbiota. Isolating the role of diet in the athlete’s gut microbiota is challenging since dietary choices depend on the demands of the exercise performed, including type, intensity, and competition level, and in turn exercise can modulate this ecosystem. Nevertheless, several clinical and preclinical research models described in the following sections have contributed to our understanding of how dietary strategies adopted by athletes can modulate the homeostasis of the gut microbiota. Data from available clinical studies evaluating dietary interventions aimed to modulate the gut microbiota of athletes with potential implications in performance or recovery are extracted and presented in Supplementary Table S1.
Protein
While protein population reference intakes for healthy adults are estimated to be 0.83 g/kg of body weight (EFSA Panel on Dietetic Products and Allergies, 2012), both resistance and endurance athletes are recommended to consume higher quantities (1.2-1.7 g/kg of body weight) (ADS/DC/ACSM, 2009) with the purpose of enhancing training adaptations and compensating muscle breakdown resulting from exercise. However, athletes are usually reported to meet or exceed these recommendations (Collins et al., 2021). As mentioned in previous sections, protein amino acids are subjected to fermentation processes that lead to the production of bacterial products and by-products (i.e. BCAAs, indole and phenolic compounds, sulphide (H2S), etc.), and some of these metabolites may contribute to undesirable processes in the host. For instance, while excess H2S has been proposed to inhibit mitochondrial cytochrome C oxidase activity, phenol has been reported to impair gut barrier function and integrity, and ammonia is thought to impair colonocyte energy metabolism (Portune et al., 2016). In settings of high protein intake, these processes might exacerbate leading to potential detrimental effects on athletes’ health. However, there is a dearth of evidence pertaining to how high-protein intake or the inadequate use of protein-based supplements can influence the composition of the gut microbiota. In a recent systematic review conducted by Wu et al. (2022) authors collected and critically analysed all interventional studies conducted in humans or animals, and in vitro research models aimed to delineate the impact of proteins, protein source and processing on the relative abundance of gut microbes. No single study conducted in humans was identified through the search strategy (Wu et al., 2022). Based on in vivo and in vitro records, authors concluded that both excess and insufficient protein intake as well as protein source are factors that can impact the composition of the gut microbiota (Wu et al., 2022). Specifically, high-protein diet was suggested to reduce the populations of propionate- and butyrate-producing bacteria and other carbohydrate utilising microbes, such as species from the Lachnospiraceae family, Akkermansia, Prevotella and Roseburia genera, which can subsequently foster the proliferation of proteolytic bacteria (Wu et al., 2022). Authors also suggested that animal proteins might elicit better effects on the gut microbiota as consequence of their higher digestibility and the lack of antinutrients (compounds that compromise the absorption of nutrients), which appears to lead to a more balanced composition of the gut microbiota (Wu et al., 2022).
The above-cited review excluded studies conducted in energy-restricted or unhealthy individuals such as overweight and obese adults (unhealthy participants) (Wu et al., 2022); however, some information can be drawn from these studies. One example consisted of a double-blinded randomised controlled trial conducted by Beaumont et al. (2017) in 38 overweight individuals receiving a 3-week isocaloric supplementation including either casein, soy protein, or maltodextrin (control). The microbiome of faecal and rectal biopsy samples was sequenced and faecal, urinary and plasma metabolomes were assessed (Beaumont et al., 2017). Authors reported no significant changes in the relative abundance of evaluated taxonomic groups, and α- and β-diversity indices remained unaffected by the different dietary interventions (Beaumont et al., 2017). However, targeted metabolic analyses of the faecal metabolome revealed notable changes denoted by higher BCAAs levels in the group allocated to casein, lower butyrate levels in both casein and soy protein groups, and higher levels of amino acid-derived bacterial compounds in the soy group (Beaumont et al., 2017). In an aforementioned study conducted by Cronin et al. (2018) where recruited subjects were predominantly overweight, authors observed that individuals allocated to a whey protein supplementation-only group displayed an altered gut viral genome compared with exercised individuals. A separate study looked at 50 healthy middle-aged men (mean BMI ≈ 27) who took part in a 10-week supervised resistance exercise training program consuming either 0.8-1.0 g/kg/day or 1.6-1,8 g/kg/day amounts of proteins (McKenna et al., 2021). Post-hoc metagenomic analyses of faecal samples revealed that subjects allocated to the high-protein group had lower within-group abundance of Veillonellaceae, Akkermansiaceae (Akkermansia), Eggerthellaceae, and Ruminococcaceae after one week of dietary habituation, while the exercise program led to increased abundance of both Eggerthellaceae and Veillonellaceae (McKenna et al., 2021). These changes were not observed in participants consuming moderate protein quantities (McKenna et al., 2021).
While the aforementioned studies were conducted in overweight or obese individuals, some of these compositional changes may be a reflection of lifestyle changes, therefore exploring associations between protein intake in healthy and lean individuals may reveal more direct effects on the gut microbiota. For instance, Estaki et al. (2016) reported that protein intake was a confounding variable for β-diversity at each tested taxonomic resolution in healthy young adults suggesting that protein intake explained the overall community compositional changes. Protein consumption also positively correlated with microbial diversity in elite rugby players in the study conducted by Clarke et al. (2014). On the other hand, while Jang et al. (2019) observed a negative correlation between daily protein intake and gut microbial α-diversity indices in distance runners, a microbial diversity similar to that of the control group was observed in bodybuilders consuming even higher amounts of proteins. Differences in other dietary components such as low fibre intake may be a contributing factor to the decrease in microbial diversity during high protein intake in endurance athletes, which could explain these results (Jang et al., 2019). One study attempted to illustrate the effect of protein supplementation in order to better understand how nutritional supplements such as whey protein would impact the gut microbiota of cross-country runners (Moreno-Perez et al., 2018). The athletes received either a protein supplement (10 g whey isolate and 10 g beef hydrolysate) or maltodextrin (control) once per day for 10 weeks (Moreno-Perez et al., 2018). Authors collected data on food intake and stool samples for 16S rRNA sequencing analyses, and measured SCFAs, pH, water, and faecal ammonia content at time zero and after the intervention (Moreno-Perez et al., 2018). A total of 18 participants were analysed at study end point, and, although no changes in α- and β-diversity indices or microbial metabolites were observed, authors reported a decrease in the relative abundance of the Lachnospiraceae family and the Roseburia, Blautia, Coprococcus genera along with lower F:B ratio in protein-supplemented athletes compared with control, which was considered by the authors as a decrease in health-promoting bacteria populations (Moreno-Perez et al., 2018).
In summary, the available research seems insufficient to draw solid conclusions on how excessive protein intake can impact the athletes’ microbiome. While evidence from preclinical research and studies conducted in obese individuals suggests that excessive protein intake might reduce populations of SCFA-producing bacteria (i.e. Roseburia and Akkermansia species), studies conducted in healthy individuals and athletes have yielded inconsistent results. Further, these effects are likely to depend on protein source and quality as well as on co-intake of different nutrients. For example, less digestible proteins, such as plant-sourced protein isolates may be more easily available for fermentation in the colon. Nonetheless, it can be argued that protein consumption exceeding recommended intake for athletes might exacerbate the production of proteolytic metabolites that are speculated to elicit deleterious effects on the athlete’s health, and therefore should be carefully managed.
Carbohydrates
Athletes often rely on the periodised consumption of carbohydrates before, during, and after exercise to maintain and refill fuel stores, a process crucial to prevent the onset of fatigue and facilitate recovery (Burke et al., 2001). To ensure energy availability and prevent gastrointestinal distress, athletes tend to avoid high-fibre diets in favour of those containing highly digestible carbohydrates (Rodriguez et al., 2009). Oligosaccharides, disaccharides, monosaccharides, and polyols (FODMAP) are also avoided in an attempt to reduce symptoms of gastrointestinal discomfort (Lis et al., 2019). Although the long-term use of these strategies might alter the composition of the athlete’s gut microbiota, no clinical studies have explored how replacing FODMAP and fibre with easily digestible carbohydrates modulates the gut microbiota in athletes. Of note, low quantities of non-absorbed digestible monosaccharides and disaccharides can also reach the colon and be available for colonic microbiota metabolism, thus the overconsumption of these carbohydrates can be conceived to have an impact on microbial populations (Chassard and Lacroix, 2013).
While the role of carbohydrate intake has not been explored in isolation, a couple of recent trials have evaluated how periodised dietary patterns, including shifts between carbohydrate and protein consumption, can impact the gut microbiota in athletes. In a cross-over study conducted by Furber et al. (2022) highly trained endurance athletes were allocated firstly to a high-protein diet (40% protein, 30% carbohydrate, and 30% fat) for one week and subsequently to a high-carbohydrate diet (10% protein, 60% carbohydrate, and 30% fat) for another week. Participants were not blinded to food items, and the diets were not thoroughly reported (Furber et al., 2022). The high-protein diet was associated with reduced time-to-exhaustion which was restored upon return to normal dietary intake, while the high carbohydrate diet led to improved performance, which also returned to base values after the intervention (Furber et al., 2022). Importantly, the dietary interventions had different impacts on the composition of the gut microbiota. The high protein diet was linked to reduced α-diversity of the viral microbiota, higher relative abundance of Sk1viruses, and enriched abundance of Leuconostoc bacterial genus (Furber et al., 2022). On the other hand, the high carbohydrate diet correlated with higher abundance of Collinsella and Ruminococcus species (Furber et al., 2022). Interestingly, improvements in performance were directly associated with greater ‘microbial stability’ among evaluated athletes, which implies that microbial communities resistant to diet-induced changes might be beneficial when performing exercise at a high level (Furber et al., 2022). An ensuing pilot trial aimed to evaluate changes in microbial dynamics following two different dietary regimens in bodybuilders, a protein-rich and low-carbohydrate diet, and a more balance post-competition diet (Luk et al., 2022). Although changes in food choices were aligned with differences in gut microbial composition and metabolomic responses, these changes were too dependent on inter-individual variability to draw conclusions (Luk et al., 2022).
While athletes may want to prioritise simple carbohydrate intake during training and competition periods to avoid short-term gastrointestinal distress and optimise performance, adequate fibre intake are warranted to avoid negative changes in gut microbial composition and function over time (Hughes and Holscher, 2021; Mohr et al., 2020). However, there is insufficient information on how long-term replacement of fibre with simple carbohydrates can impact the gut microbiota in athletes.
Lipids
Since carbohydrates are the main source of energy during exercise, and fatigue is closely associated with a depletion of glycogen stores, athletes typically resource to glycogen loading strategies that often imply reduced consumption of fat at levels even below those recommended for the general population (Pendergast et al., 2000). In sharp contrast, several authors have proposed the potential ergogenic effects that ketogenic diets, in other words, diets characterised by low-carbohydrate and high-fat intake (>75% energy), may have on athletes (McSwiney et al., 2019). The impact that these opposing strategies might have on the gut microbiota in regard to athletic performance has been largely overlooked in the literature.
High-fat diet (HFD) animal models have been extensively used to study the role that diet plays in metabolic disorders such as obesity, type 2 diabetes, hepatic steatosis, etc. (Buettner et al., 2007) Moreover, HFD is probably the most widely studied dietary intervention when assessing changes in the gut microbiota. In a large-scale meta-analysis conducted by Bisanz et al. (2019) data from 25 studies conducted in rodents and two additional trials recruiting humans were pooled to identify reproducible gut microbiome responses to HFD (Bisanz et al., 2019). Although studies differed considerably in terms of definition of HFD used (27.1%-65% fat), sequencing strategy, and variable regions explored, authors found a consistent increase in F:B with higher fat content, as well as modest significant decreases in richness (Chao1) and diversity (Shannon) indices (Bisanz et al., 2019). Further, 228 OTUs were shown to discriminate between high- and low-fat diets (Bisanz et al., 2019). While the amount of carbohydrates in evaluated diets was highly variable, this study provided solid evidence on the effects that high fat intake might exert on the composition of the gut microbiota.
Unlike HFD, ketogenic diets are characterised by low carbohydrate intake, which leads to the production of ketonic bodies as a result of low glucose availability. As suggested by a systematic review on the impact of ketogenic diets on the gut microbiota, findings are controversial and there is a need for deeper understanding (Paoli et al., 2019). Among the nine articles included in this review, eight studies explored animal models or enrolled non-healthy individuals (i.e. autism, multiple sclerosis patients), and only one reported data on healthy adults (Paoli et al., 2019). In particular, Murtaza et al. (2019) enrolled elite race walkers receiving either a high-carbohydrate diet (60% carbohydrates, 16% protein, 20% fat), a periodised high-carbohydrate diet with the same composition, or a ketogenic diet (3.5% carbohydrates, 17% protein, 78% fat) for three weeks. Athletes provided stool samples for metagenomic analyses and took part in an intensified training program (Murtaza et al., 2019). Notably, athletes receiving the ketogenic diet reported poorer performance in a 10 km race, and impaired exercise economy compared with athletes consuming the high-carbohydrate diets (Murtaza et al., 2019). Moreover, the most profound effects on the gut microbiota were observed in athletes consuming the ketogenic diet, which were denoted by reduced abundance of Faecalibacterium spp., and increased abundance of Dorea spp. and several OTUs belonging to the Bacteroides genus (Murtaza et al., 2019). Importantly, these changes negatively correlated with performance outcomes in these athletes (Murtaza et al., 2019). In a more recent study, Mancin et al. (2022a) reported no effects on gut microbial diversity in semi-professional soccer players allocated to a 30-day ‘ketogenic Mediterranean diet with phytoextracts’, yet a decrease in Actinobacteria (recently renamed to Actinomycetota (Oren and Garrity, 2021)) and an increase in bacteria from the Clostridia, Butyricimonas, Odoribacter, and Ruminococcus taxa was reported in these subjects. Accordingly, the authors suggested that different types of ketogenic diets might elicit different types of effects (Mancin et al., 2022a).
A factor contributing to the different effects observed with the abovementioned ketogenic diets might be the type of lipids provided. In fact, the ketogenic Mediterranean diet explored by Mancin et al. (2022a) was rich in PUFAs, which were proposed to improve gut epithelial integrity thus indirectly contributing to the maintenance of the gut microbiota. On this subject, an interesting systematic review conducted by Wolters et al. (2019) aimed to evaluate how not only the quantity but also the quality of dietary fat intake can modulate the gut microbiota in humans. Notwithstanding limitations inherent to the methods used in the included studies, authors concluded that diets rich in saturated fatty acids were likely to exert unfavourable effects on the gut microbiota denoted by a reduction in richness and diversity (Wolters et al., 2019). On the other hand, polyunsaturated fatty acids (PUFAs) did not appear to modulate these indices nor negatively impact metabolic health outcomes in studied individuals (Wolters et al., 2019). Interestingly, in female elite soccer players competing in an official tournament, the relative abundance of Faecalibacterium genus was shown to indirectly correlate with daily total fat, saturated fat, and red meat intakes (Oliveira et al., 2022). All these data are consistent with the fact that saturated fatty acids are typically linked to pro-inflammatory processes which mimics the action of LPS (Fritsche, 2015), but PUFAs (particularly n3-PUFAs) are associated with improved inflammatory status (Calder, 2006). Further, n3-PUFAs have been associated with enhanced SCFA production (Robertson et al., 2017), decreased intestinal barrier permeability (Lam et al., 2015), and might regulate immune processes (Radzikowska et al., 2019). Although available evidence is weak, athletes might opt for avoiding both high-fat and low-fat diets in favour of more balance dietary alternatives where polyunsaturated fatty acids should play an important role. These might follow the principles of the Mediterranean diet but adapted to contemporary sports nutrition practices (Griffiths et al., 2022).
Lastly, the influence on the gut microbiota of dietary strategies consisting of reduced fat intake has not been specifically explored. Since lipids are clearly involved in the production of bile acids, and as mentioned, secondary bile acids are speculated to play important roles in muscle function, adequate lipid intake might positively contribute to the interplay between gut microbiota and skeletal muscle.
Biotics
The term ‘Biotics’ encompasses a family of dietary strategies conceived to directly target the gut microbiota by modulating the composition and functional output (Kocot et al., 2022). Among them, prebiotics, probiotics, and synbiotics are the most widely studied and have gained great interest over the years due to their potential role in the gut-muscle axis.
The International Society for Probiotics and Prebiotics (ISAPP) defines prebiotics as ‘substrates that are selectively utilised by host microorganisms conferring a health benefit’ (Gibson et al., 2017). Carbohydrates such as fermentable fibres, milk oligosaccharides (HMO), and non-digestible oligosaccharides, such as fructooligosaccharides (FOS) and galactooligosaccharides (GOS), fall under the definition of prebiotics (Gibson et al., 2017). The fermentation of these ingredients leads to the selective proliferation of bacterial species that, in turn, generate functional metabolites such as SCFAs, thus conferring a beneficial effect on the host (Gibson et al., 2017). Although athletes might be reluctant to consume fermentable carbohydrates in order to prevent gastrointestinal distress during competition, some authors have considered prebiotics as a viable dietary strategy to optimise the gut microbiota in athletes (O’Brien et al., 2022).
A far more explored approach consists of providing live microorganisms to selectively manipulate the composition of the gut microbiota. In 2014, the ISPP retained the WHO/FAO definition of probiotics as ‘live microorganisms that, when administered in adequate amounts, confer a health benefit on the host’ (Hill et al., 2014). Some of these beneficial effects such as improving gastrointestinal health, modulating immune status, and regulating inflammatory and oxidative responses may positively impact performance and recovery from exercise. Naturally, probiotic interventions must engraft and establish themselves as community members of the gut microbiota to mediate these effects, all of which may depend on strain selection and dose provided.
As presented in Supplementary Table S1, clinical studies aimed to modulate the gut microbiota of athletes to impact outcomes of performance and/or recovery largely consist of probiotic interventions. Overall, mixed results on the effect of probiotic administration on VO2max in athletes have been reported (Lamprecht et al., 2012; Marshall et al., 2017; Mazur-Kurach et al., 2022). While some interventional studies have reported improvements in performance or recovery outcomes in endurance (i.e. improved endurance performance and recovery of muscle function from exercise-induced muscle damage) (Huang et al., 2020; Lin et al., 2020) and resistance athletes (Jager et al., 2016), others have not observed such effects (Carbuhn et al., 2018; Strasser et al., 2016; Townsend et al., 2018). Thus, a direct link between probiotic consumption and improved performance or accelerated recovery cannot be concluded from the available literature.
While evidence remains inconclusive, indirect effects on maintaining performance might be mediated by the potential role of probiotics in improving gastrointestinal health and immune function. In this sense, athletes are frequently exposed to stress and intense exercise efforts that can result in gastrointestinal discomfort, and might compromise the gut barrier integrity thus facilitating the uptake of microbial toxins and limiting nutrients absorption (Miles, 2020). Some clinical studies evaluating probiotic interventions have reported reduced severity and incidence of gastrointestinal symptoms in cyclists and marathon runners (Lamprecht et al., 2012; Pugh et al., 2019; West et al., 2011) as well as shorter duration of these symptoms during training and competition (Kekkonen et al., 2007). Further, some evidence suggests that probiotics might address exercise-induced enhanced gastrointestinal permeability. Specifically, long-term administration of multi-strain probiotic products to endurance athletes has been shown to notably reduce zonulin levels indicating improved intestinal barrier status (Lamprecht et al., 2012; Mazur-Kurach et al., 2022), yet mixed results have been reported in team-sports players (Townsend et al., 2018). Similarly, while multi-strain probiotic supplementation has been shown to decrease post-exercise plasmatic LPS levels in trained runners (Shing et al., 2014), and triathletes (Roberts et al., 2016), a single-strain intervention led to no change in LPS levels during offseason training in swimmers (Carbuhn et al., 2018). It seems therefore that multi-strain probiotic products are more likely to exert beneficial effects on gastrointestinal health than single-strain products, but further research is needed to understand the role of probiotics in addressing gastrointestinal symptoms in athletes.
Prolonged and high-intensity exercises are known to induce shifts in white blood cell (WBC) counts denoted by decreases in lymphocyte populations and increases in granulocyte and monocyte counts during the recovery period (Simpson et al., 2015). If maintained, this blunted immune response has been proposed to represent an ‘open window’, where an increase in the athlete’s susceptibility to upper respiratory tract infections (URTI) is hypothesised (Pedersen et al., 1996). Since probiotics are proposed to modulate the immune response, several authors have explored probiotic interventions with the purpose of preventing URTI and improving immune-related outcomes in athletes; however, results have been shown to be inconsistent. The long-term administration of either multi-strain or single-strain probiotic products has been reported to reduce the incidence and duration of symptoms of URTI in athletes of different sports disciplines (Cox et al., 2010; Michalickova et al., 2016; Strasser et al., 2016). On the other hand, several studies have reported null or unclear associations (Carbuhn et al., 2018; West et al., 2011). Similarly, concentrations of immune or stress-related markers such as different WBC counts and salivary IgA, human cathelicidin, β-defensin, and α-amylase after exhaustive exercise have been shown to be modulated by probiotic interventions in some studies (Batatinha et al., 2020; Pumpa et al., 2019; Vaisberg et al., 2019), while others have reported mixed or null results (Carbuhn et al., 2018; West et al., 2011). Although a consensus statement from the International Society of Sports Nutrition (ISSN) concluded positively on the consumption of probiotics to prevent or ameliorate infections (‘Regular consumption of specific probiotic strains may assist with immune function and may reduce the number of sick days an athlete experiences when training or during competition’) (Jäger et al., 2019), uncertainties surrounding strains explored, doses, sex-based differences, type of exercise performed, validity of assessed markers, and the low sample size of available studies may hinder drawing solid conclusions on the impact of probiotic supplementation on athletes’ immune status.
Another interesting aspect of probiotic supplementation in athletes consists of their potential role in ameliorating inflammatory and oxidative responses to exercise. Through improving the intestinal barrier, probiotics might prevent or ameliorate the LPS-mediated low-grade inflammatory response (Lamprecht et al., 2012). Further, the immunomodulatory effects of probiotics might lead to reduced inflammatory response to exhaustive or muscle-damaging exercise, which is linked to improved recovery. Regarding inflammation, interleukins (i.e. IL-6), tumour necrosis factor-α (TNF-α), and c-reactive protein (CRP) are typically assessed as pro-inflammatory markers to evaluate the effect of probiotics. Probiotic supplementation has been reported to reduce cytokine levels (i.e. TNF-α, IL-6, IL-10) after acute (Huang et al., 2019; Lamprecht et al., 2012) and long-term intense exercise (Townsend et al., 2018; West et al., 2011). On the other hand, different studies have observed mixed results when evaluating probiotic supplements to improve cytokine profiles after exercise in athletes (Jager et al., 2016; Pugh et al., 2019). Circulating levels of pro-inflammatory markers following exercise can vary depending on sampling time and on the nature of the exercise performed. For example, increases in CRP derived from exercise-induced muscle damage occur at 24-48 h after muscle-damaging exercise (Silva et al., 2018), while exercise-induced increases in IL-6 concentrations are mainly due to local production in the skeletal muscle, and rapidly decrease during the first hours within the recovery period (Silva et al., 2018). In general, improvements in TNF-α might be derived from improved gut barrier structure, which in turn might improve nutrient assimilation thus leading to potential beneficial effects on recovery from exercise in athletes. Nonetheless, further research is needed to elucidate these effects.
Oxidative stress as consequence of exhaustive exercise is thought to be a potential contributor to skeletal muscle fatigue and overtraining syndrome in athletes (Finaud et al., 2006). Probiotics such as those belonging to the Bifidobacterium and former Lactobacillus genera (i.e. Lacticaseibacillus rhamnosus, Limosilactobacillus fermentum, etc.), have been shown to display free radical scavenging activities and upregulate the expression of antioxidant enzymes (Mishra et al., 2015). Other already-mentioned properties might further contribute to these anti-oxidant effects, such as decreased cytokine-mediated oxidative response, enhanced uptake of dietary antioxidants, etc. (Martarelli et al., 2011). However, evaluating the impact of probiotic supplementation on exercise-induce oxidative stress is challenging due to limitations inherent to available biomarkers (Gomez-Cabrera et al., 2021; Mishra et al., 2015). Although some authors have reported beneficial effects of probiotic supplementation on oxidative status in athletes under physical stress, denoted by improvements in several markers of oxidative status (i.e. total antioxidant status, myeloperoxidase levels) (Huang et al., 2020; Martarelli et al., 2011), others have reported negative or inconsistent results (Lamprecht et al., 2012; Valimaki et al., 2012). Harmonisation of methods employed is warranted to shed light on the effect of probiotics on oxidative status in athletes.
Of note, only a few studies have explored combinations of probiotics and prebiotics, such as FOS, in athletes (Fritz et al., 2024; Marshall et al., 2017; Quero et al., 2021; Roberts et al., 2016). Among these, Fritz et al. (2024) reported findings from a recent study evaluating the impact of combined 31-day prebiotic and multi-strain probiotic supplementation on body composition and gut microbiome outcomes in 10 male elite water polo players consuming a plant-based vegan protein supplement compared to a matched control group consuming only the protein supplement. Interestingly, a decrease in fat body mass, and an increase in skeletal muscle mass were observed only in the prebiotic- and probiotic-supplemented group (Fritz et al., 2024). Further, changes in muscle mass were correlated with activation of some SCFA-related functional pathways, which authors speculated to denote improved efficiency of the gut microbiota of the intervention-group athletes (Fritz et al., 2024). Notably, rather than a simple mixture of probiotics and prebiotics, the ISAPP defines synbiotics as ‘a mixture comprising live microorganisms and substrate(s) selectively utilised by host microorganisms that confers a health benefit on the host’ (Swanson et al., 2020). Thus, it is unclear if cited combinations of probiotics and prebiotics fall under this category, or if additional benefits can be perceived from these combinations. Lastly, no single study to date has explored postbiotic interventions (‘preparation of inanimate microorganisms and/or their components that confers a health benefit on the host’ (Salminen et al., 2021)) in athletes, representing a potential dietary strategy in future research.
Other dietary strategies
Athletes rely on different dietary strategies to achieve training and competition goals often based on personal preferences. These include the use of ergogenic supplements, recovery products, as well as changes in nutrient timing and energy availability. Further, high-level sports practice leads to changes in energy balance and sweat-induced dehydration. All these factors are speculated to contribute to the characteristics of the athlete gut microbiota, yet a further understanding of the processes involved is needed.
For instance, polyphenols are a set of compounds that are candidate to fall under the classification of prebiotics (Gibson et al., 2017). Polyphenols and polyphenol-rich ingredients (i.e. green tea, tart cherry, pomegranate, curcumin) are often used to improve recovery from muscle damage due to their anti-inflammatory and antioxidative properties (D’Angelo, 2020). Mainly based on preclinical evidence, ingredients such as tea and berries, contain phenolic compounds that might modulate the gut microbiota leading to increases in Bifidobacterium, former Lactobacillus, Roseburia and Akkermansia species, and produce bioactive bacterial intermediates that might modulate skeletal muscle function (Ghosh et al., 2020; Lavefve et al., 2020; Liu et al., 2020). Nonetheless, recent clinical research suggested a minimal impact of 10-week consumption of polyphenol-rich cocoa on the gut microbiota, and plasma and faecal metabolome of male athletes (Tabone et al., 2022). In the same line, athletes frequently consume micronutrients, such as vitamin D, iron, and calcium, as supplements to avoid suboptimal status that might impair performance and recovery (Maughan et al., 2018). Altogether, reduced micronutrients availability seems to negatively impact commensal gut microbiota leading to colonisation and proliferation of pathogens (Mach and Clark, 2017), yet no single study to date has explored the effects that micronutrient intakes have on the microbiota of non-deficient athletes.
Due to the needs of high-level competitive sports, athletes are encouraged to achieve high energy intake in order not to reach a negative energy balance, which can lead to the syndrome termed ‘relative energy deficiency in sport’ (RED-S) (Holtzman and Ackerman, 2019). However, different sports disciplines may have specific energy requirements. For example, American football, sumo, and wrestling are sports where positive energy balance is sometimes favoured. On the contrary, transitory energy restriction can be pursued by wrestlers to participate in a certain wrestling weight category or by team sports athletes returning from offseason. Changes in energy balance might exert changes in the gut microbiota, as observed from comparisons between diets with similar macronutrient profile but different caloric content (2,400 kcal/day vs 3,400 kcal/day) showing increased F:B ratio in individuals consuming the high-calorie diet (Jumpertz et al., 2011). Further, the gut microbiota can also contribute to energy availability, which is even more important when high quantities of nutrients are available for bacterial metabolism in the colon. All these aspects make it difficult to directly evaluate associations between energy balance and the gut microbiota in athletes, and, therefore, most data on changes in energy balance are drawn from pathological populations characterised by extreme dietary patterns, such as anorexia nervosa patients and undernourished children (Iddrisu et al., 2021; Mörkl et al., 2017).
Lastly, the role of hydration status and its impact on the gut microbiota of athletes remains largely unexplored. A growing body of literature supports a bidirectional association between constipation and the gut microbiota (Zhang et al., 2021). Mild dehydration impacts water balance and promotes constipation (Arnaud, 2003), and athletes are frequently exposed to large fluid losses in the form of sweat, which can lead to dehydration and is linked to poor performance (Judelson et al., 2007). However, exercise has been consistently shown to associate with improved symptoms of constipation, which makes it difficult to draw conclusions (Gao et al., 2019). In a recent study, 146 healthy adults were assessed for correlations between known hydration markers and faecal microbiota (Willis et al., 2021). Species in the Bacteroides, Desulfovibrio, Roseburia, Peptococcus and Akkermansia genera were shown to be in higher relative abundance in individuals considered to be in a poorer hydration state, yet no rationale nor mechanism underlying these changes was suggested (Willis et al., 2021). Further work needs to be done to further understand hydration and its impact on the gut microbiota and gut barrier integrity.
5 Practical approach to the athlete gut microbiota
Higher accessibility to metagenomic sequencing platforms and the rise of companies offering increasingly affordable omics services have provided athletes, coaches, and sports physicians the opportunity of profiling faecal samples to identify potential causes underlying gastrointestinal symptoms and guide dietary protocols. While there is a lack of consensus on the features that characterise the athlete’s gut microbiota and the role that the athlete’s diet plays in this ecosystem, some available faecal indicators frequently reported in these analyses might be useful to guide simple dietary approaches aimed to improve the gut microbiota of athletes.
Diversity metrics are the most widely explored indices in gut microbiome studies. Low α-diversity is a sign of dependence instead of coexistence between microbial populations (Karkman et al., 2017). On the contrary, high α-diversity denotes a more stable microbiota, which might be more resilient to environmental aggressions such as pathogens and antibiotic usage (Karkman et al., 2017). Further, a growing body of evidence discussed in previous sections suggests that elevated gut α-diversity is a hallmark of the changes produced by long-term exercise and fitness. Overall, the evidence strongly suggests that a decrement in α-diversity is a negative feature of the gut microbial environment, and a potential sign of dysbiosis in athletes. Nevertheless, although diversity indices are powerful tools to provide insights into the overall composition of the gut microbiota, inconsistencies are also present in the literature, which remarks that a single metric is insufficient when assessing microbiome profiles, and different indices might be needed to assess α-diversity. Of note, fermented food intake has been reported as a valid dietary strategy to increase gut bacteria diversity, yet caution should be warranted when generalising these findings (Wastyk et al., 2021).
Changes in phyla ratios, such as F:B, have been subject to extensive research in clinical populations (Magne et al., 2020), but how deviations in F:B can translate to the gut microbiota of healthy athletes is less explored. Since Firmicutes and Bacteroidetes (renamed to Bacillota and Bacteroidota) (Oren and Garrity, 2021) represent over 90% of all gut bacteria, extreme changes in these ratios are expected to contribute to dysbiotic states, yet how diet impacts these indices is not clear. Proteobacteria and Actinobacteria (renamed to Pseudomonadota and Actinomycetota) (Oren and Garrity, 2021) are the third and fourth most abundant bacteria phyla in the human gut, respectively. While expansions in Proteobacteria are consistently linked to inflammatory and metabolic conditions (Shin et al., 2015), families from the Actinobacteria phylum, such as Bifidobacteriaceae, are pivotal for the maintenance of the gastrointestinal barrier, among other functions involved in gut homeostasis (Binda et al., 2018). Species belonging to the Actinobacteria and Firmicutes phyla are among the main responders to dietary fibre (Makki et al., 2018), while increased levels of Proteobacteria have been associated to low-fibre diets (Shin et al., 2015). Even though changes in phyla ratios might be considered as an oversimplification of the complex interactions existing in the gut microbiota and might not necessarily indicate dysbiosis, a depletion in Actinobacteria and a drastic increase in the relative abundance of Proteobacteria might be linked to inadequate fibre intake, and dietary interventions aimed to restore this balance should be considered.
At a different taxonomic resolution, bacteria species such as A. muciniphila and Faecalibacterium prausnitzii are among the most promising emerging next-generation probiotics (Kumari et al., 2021). While F. prausnitzii is one of the major butyrate-producers present in the healthy gut microbiota (Louis and Flint, 2017), A. muciniphila has been considered an exercise-responsive taxon (Munukka et al., 2018) that seems to play key roles in the remodelling of the colonic mucus (Chen et al., 2021). As shown in previous sections, these two species are among the most widely researched when exploring associations between physical activity, exercise, and the gut microbiota (i.e. ref. Bressa et al., 2017; O’Donovan et al., 2020). Other genera, such as Prevotella and Parabacteroides, have been considered as exercise-responsive taxa (Jang et al., 2019; Petersen et al., 2017), but further research might be needed to confirm these findings. Additionally, high relative abundance of SCFA-producing bacteria from genera belonging to the Firmicutes phylum, such as Eubacterium and Roseburia (i.e. E. rectale and R. hominis), has been proposed as a sign of a healthy gut microbial environment. In line with the aforementioned changes in phyla ratios, metagenomic analyses might place a special focus on the low relative abundance of these beneficial bacteria, particularly when combined with low faecal SCFA levels, which might suggest inadequate fibre intake, and potentially impact metabolic efficiency in athletes.
Faecal analyses might provide additional insights into how dietary strategies adopted by athletes influence their health outcomes. Protein intake exceeding recommendations for athletes and the abuse of protein supplements when recommended intakes are met might lead to exacerbated faecal levels of protein fermentation products, such as p-cresol, ammonia and indoles, which in turn are proposed to negatively impact renal and intestinal barrier function (Felizardo et al., 2016; Portune et al., 2016). In addition, blood samples might supplement information reported in faecal analyses. Elevated consumption of animal food proteins (especially red meat) and supplementation with L-carnitine have been associated with microbial production of TMA, which is transformed into TMAO in liver and represents a generally considered deleterious molecule consistently associated with cardiovascular disease (Yang et al., 2019). Similarly, analyses of gut barrier status markers (i.e. zonulin) in combination with circulating levels of LPS and pro-inflammatory markers, can be informative when assessing increased permeability of the gastrointestinal barrier, particularly in the presence of gastrointestinal symptoms and after strenuous exercise in the heat, where splanchnic blood flow can be compromised (Yang et al., 2019). Supplementing faecal analyses with blood monitoring of these molecules might provide additional information when exploring underlying causes of gastrointestinal symptoms in athletes, and help take dietary measures to prevent the excessive microbial production of detrimental metabolites.
To date, it is unclear if probiotic interventions can modulate microbial structural indices of diversity or dissimilarity, or restore dysbiosis following disruptive events (Kristensen et al., 2016; Lynne, 2014). As concluded by Sniffen et al. (2018) in an analysis of 249 probiotic trials, no single probiotic formulation is supported by ‘strong’ or ‘moderate’ evidence for preventing respiratory tract infections. On the other hand, some formulations might be well-supported for the treatment or prevention of specific gastrointestinal symptoms such as constipation and acute diarrhoea (Sniffen et al., 2018). Thus, choosing an appropriate probiotic should be based on strain- and disease-specific effectiveness of the product, and not as a general tool to restore dysbiotic states. While several multi-strain probiotic formulations have been reported to improve the structure of the gastrointestinal barrier in studies discussed throughout the manuscript, only formulations designed for the treatment of gastrointestinal disorders (i.e. intestinal bowel disease) have demonstrated efficacy (Sniffen et al., 2018), and there is insufficient evidence on optimal compositions for improving gastrointestinal barrier function in healthy populations.
Lastly, it is important to remark that a single analysis is only a snapshot of the complex dynamic picture that represents the human gut microbiota, and longitudinal repeated testing using both metagenomic and metabolomic approaches should be considered when evaluating the need of any nutritional intervention. Further, what may represent dysbiosis in some athletes, may not in others, and the use of exploratory analyses should be based on the presence of gastrointestinal symptoms, which have been clearly associated with dysbiotic states (Wei et al., 2021). A scheme summarising discussed practical approach is presented in Figure 4.
Evidence on factors contributing to gut dysbiosis and dietary recommendations to address them. F:B. = Firmicutes-Bacteroidetes ratio; FOS = fructooligosaccharides; GI = gastrointestinal; GOS = galactooligosaccharides; LPS = lipopolysaccharide; SCFA = short-chain fatty acids; TMAO = trimethylamine n-oxide; ↑/↓ = moderate increases/decreases;
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00007
6 Conclusions
Athletes appear to display a metabolically favourable gut microbial environment characterised by higher diversity and relative abundance of beneficial bacteria, a consequence of years of high-level sports practice and adherence to specific dietary patterns. Improved metabolic efficiency seems to be mediated by the production of bacterial metabolites, mainly SCFAs, which in turn can impact performance and recovery from exercise. Thus, sports professionals might resort to faecal analyses to explore strategies aimed to optimise this interplay. To this end, indicators commonly reported in microbiome analyses, such as diversity indices and relative abundance of generally considered beneficial bacteria, might be useful to evaluate dysbiotic states in the athlete. Longitudinal repeated testing might provide valuable insights to guide simple dietary approaches supported by strong evidence to improve the gut microbiota. The science in this field evolves rapidly, and new evidence on different bacterial metabolites, such as secondary bile acids, and dietary strategies, including the use of probiotic formulations and fermented foods, might further support and expand on these practical applications in the future.
Corresponding author; e-mail: inigomaria.perez@abbott.com
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.25534273
Table S1. Dietary interventions targeting the gut microbiota to improve performance or recovery outcomes in athletes.
Acknowledgements
The present joint research initiative originates from the Abbott-Real Madrid Innovation Lab, a collaborative framework between Abbott and Real Madrid endeavoured to advance scientific knowledge in sports physiology and human nutrition.
Authors’ contribution
IPC, RR, JLC, ASD, HB, and NM contributed to study conceptualization and design. IPC and ASD performed the literature search. IPC draft the manuscript. RR, ASD, JLC, HB, and NM substantially reviewed and edited the manuscript. All authors approved the final submitted version of the manuscript.
Conflict of interest
ÍPC, RR, ASD, and HB were employed by Abbott Laboratories. JLC and NM were employed by Real Madrid.
Funding
This research received no external funding.
References
American Dietetic Association, Dietitians of Canada, and the American College of Sports Medicine (ADS/DC/ACSM), 2009. Nutrition and athletic performance. Medicine and Science in Sports and Exercise 41: 709-731. https://doi.org/10.1249/MSS.0b013e31890eb86
Allen, J.M., Mailing, L.J., Niemiro, G.M., Moore, R., Cook, M.D., White, B.A., Holscher, H.D. and Woods, J.A., 2018. Exercise alters gut microbiota composition and function in lean and obese humans. Medicine and Science in Sports and Exercise 50: 747-757. https://doi.org/10.1249/MSS.0000000000001495
Arnaud, M.J., 2003. Mild dehydration: a risk factor of constipation? European Journal of Clinical Nutrition 57: S88-S95. https://doi.org/10.1038/sj.ejcn.1601907
Aziz, Q., Doré, J., Emmanuel, A., Guarner, F. and Quigley, E.M.M., 2013. Gut microbiota and gastrointestinal health: current concepts and future directions. Neurogastroenterology and Motility 25: 4-15. https://doi.org/10.1111/nmo.12046
Barton, W., Cronin, O., Garcia-Perez, I., Whiston, R., Holmes, E., Woods, T., Molloy, C.B., Molloy, M.G., Shanahan, F., Cotter, P.D. and O’Sullivan, O., 2021. The effects of sustained fitness improvement on the gut microbiome: a longitudinal, repeated measures case-study approach. Translational Sports Medicine 4: 174-192. https://doi.org/10.1002/tsm2.215
Barton, W., Penney, N.C., Cronin, O., Garcia-Perez, I., Molloy, M.G., Holmes, E., Shanahan, F., Cotter, P.D. and Sullivan, O., 2018. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 67: 625. https://doi.org/10.1136/gutjnl-2016-313627
Batatinha, H., Tavares-Silva, E., Leite, G.S.F., Resende, A.S., Albuquerque, J.A.T., Arslanian, C., Fock, R.A., Lancha, A.H., Jr., Lira, F.S., Kruger, K., Thomatieli-Santos, R. and Rosa-Neto, J.C., 2020. Probiotic supplementation in marathonists and its impact on lymphocyte population and function after a marathon: a randomized placebo-controlled double-blind study. Science Reports 10: 18777. https://doi.org/10.1038/s41598-020-75464-0
Beaumont, M., Portune, K.J., Steuer, N., Lan, A., Cerrudo, V., Audebert, M., Dumont, F., Mancano, G., Khodorova, N., Andriamihaja, M., Airinei, G., Tomé, D., Benamouzig, R., Davila, A.M., Claus, S.P., Sanz, Y. and Blachier, F., 2017. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: a randomized, parallel, double-blind trial in overweight humans. American Journal of Clinical Nutrition 106: 1005-1019. https://doi.org/10.3945/ajcn.117.158816
Binda, C., Lopetuso, L.R., Rizzatti, G., Gibiino, G., Cennamo, V. and Gasbarrini, A., 2018. Actinobacteria: a relevant minority for the maintenance of gut homeostasis. Digestive and Liver Disease 50: 421-428.
Bisanz, J.E., Upadhyay, V., Turnbaugh, J.A., Ly, K. and Turnbaugh, P.J., 2019. Meta-analysis reveals reproducible gut microbiome alterations in response to a high-fat diet. Cell Host Microbe 26: 265-272. e264. https://doi.org/10.1016/j.chom.2019.06.013
Boets, E., Gomand, S.V., Deroover, L., Preston, T., Vermeulen, K., De Preter, V., Hamer, H.M., Van den Mooter, G., De Vuyst, L., Courtin, C.M., Annaert, P., Delcour, J.A. and Verbeke, K.A., 2017. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: a stable isotope study. Journal of Physiology 595: 541-555. https://doi.org/10.1113/JP272613
Bressa, C., Bailen-Andrino, M., Perez-Santiago, J., Gonzalez-Soltero, R., Perez, M., Montalvo-Lominchar, M.G., Mate-Munoz, J.L., Dominguez, R., Moreno, D. and Larrosa, M., 2017. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 12: e0171352. https://doi.org/10.1371/journal.pone.0171352
Buettner, R., Schölmerich, J. and Bollheimer, L.C., 2007. High-fat diets: modeling the metabolic disorders of human obesity in rodents. Obesity 15: 798-808. https://doi.org/10.1038/oby.2007.608
Burke, L.M., Cox, G.R., Culmmings, N.K. and Desbrow, B., 2001. Guidelines for daily carbohydrate intake: do athletes achieve them? Sports Medicine 31: 267-299. https://doi.org/10.2165/00007256-200131040-00003
Bycura, D., Santos, A.C., Shiffer, A., Kyman, S., Winfree, K., Sutliffe, J., Pearson, T., Sonderegger, D., Cope, E. and Caporaso, J.G., 2021. Impact of different exercise modalities on the human gut microbiome. Sports 9: 14. https://doi.org/10.3390/sports9020014
Calder, P.C., 2006. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. American Journal of Clinical Nutrition 83: 1505S-1519S. https://doi.org/10.1093/ajcn/83.6.1505S
Carbuhn, A.F., Reynolds, S.M., Campbell, C.W., Bradford, L.A., Deckert, J.A., Kreutzer, A. and Fry, A.C., 2018. Effects of probiotic (Bifidobacterium longum 35624) supplementation on exercise performance, immune modulation, and cognitive outlook in division I female swimmers. Sports 6: 116. https://doi.org/10.3390/sports6040116
Castellanos, N., Diez, G.G., Antunez-Almagro, C., Bailen, M., Bressa, C., Gonzalez Soltero, R., Perez, M. and Larrosa, M., 2019. A critical mutualism – competition interplay underlies the loss of microbial diversity in sedentary lifestyle. Frontiers in Microbiology 10: 3142. https://doi.org/10.3389/fmicb.2019.03142
Cerda, B., Perez, M., Perez-Santiago, J.D., Tornero-Aguilera, J.F., Gonzalez-Soltero, R. and Larrosa, M., 2016. Gut microbiota modification: another piece in the puzzle of the benefits of physical exercise in health? Frontiers in Physiology 7: 51. https://doi.org/10.3389/fphys.2016.00051
Chantler, S., Griffiths, A., Matu, J., Davison, G., Jones, B. and Deighton, K., 2021. The effects of exercise on indirect markers of gut damage and permeability: a systematic review and meta-analysis. Sports Medicine 51: 113-124. https://doi.org/10.1007/s40279-020-01348-y
Chassard, C. and Lacroix, C., 2013. Carbohydrates and the human gut microbiota. Current Opinion in Clinical Nutrition and Metabolic Care 16: 453-460. https://doi.org/10.1097/MCO.0b013e3283619e63
Chen, T., Wang, R., Duan, Z., Yuan, X., Ding, Y., Feng, Z., Bu, F., Liu, L., Wang, Q., Zhou, J., Zhu, L., Ni, Q., Shi, G. and Chen, Y., 2021. Akkermansia muciniphila protects against psychological disorder-induced gut microbiota-mediated colonic mucosal barrier damage and aggravation of colitis. Frontiers in Cellular and Infection Microbiology 11: 723856. https://doi.org/10.3389/fcimb.2021.723856
Ciesielska, A., Matyjek, M. and Kwiatkowska, K., 2021. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cellular and Molecular Life Sciences 78: 1233-1261. https://doi.org/10.1007/s00018-020-03656-y
Clark, A. and Mach, N., 2016. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: a systematic review for athletes. Journal of the International Society of Sports Nutrition 13: 43. https://doi.org/10.1186/s12970-016-0155-6
Clarke, S.F., Murphy, E.F., O’Sullivan, O., Lucey, A.J., Humphreys, M., Hogan, A., Hayes, P., O’Reilly, M., Jeffery, I.B., Wood-Martin, R., Kerins, D.M., Quigley, E., Ross, R.P., O’Toole, P.W., Molloy, M.G., Falvey, E., Shanahan, F. and Cotter, P.D., 2014. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 63: 1913-1920. https://doi.org/10.1136/gutjnl-2013-306541
Collins, J., Maughan, R.J., Gleeson, M., Bilsborough, J., Jeukendrup, A., Morton, J.P., Phillips, S.M., Armstrong, L., Burke, L.M., Close, G.L., Duffield, R., Larson-Meyer, E., Louis, J., Medina, D., Meyer, F., Rollo, I., Sundgot-Borgen, J., Wall, B.T., Boullosa, B., Dupont, G., Lizarraga, A., Res, P., Bizzini, M., Castagna, C., Cowie, C.M., D’Hooghe, M., Geyer, H., Meyer, T., Papadimitriou, N., Vouillamoz, M. and McCall, A., 2021. UEFA expert group statement on nutrition in elite football. Current evidence to inform practical recommendations and guide future research. British Journal of Sports Medicine 55: 416-416. https://doi.org/10.1136/bjsports-2019-101961
Cox, A.J., Pyne, D.B., Saunders, P.U. and Fricker, P.A., 2010. Oral administration of the probiotic Lactobacillus fermentum VRI-003 and mucosal immunity in endurance athletes. British Journal of Sports Medicine 44: 222-226. https://doi.org/10.1136/bjsm.2007.044628
Cronin, O., Barton, W., Skuse, P., Penney, N.C., Garcia-Perez, I., Murphy, E.F., Woods, T., Nugent, H., Fanning, A., Melgar, S., Falvey, E.C., Holmes, E., Cotter, P.D., O’Sullivan, O., Molloy, M.G. and Shanahan, F., 2018. A prospective metagenomic and metabolomic analysis of the impact of exercise and/or whey protein supplementation on the gut microbiome of sedentary adults. mSystems 3: e00044-18. https://doi.org/10.1128/mSystems.00044-18
Dainese, R., Serra, J., Azpiroz, F. and Malagelada, J.R., 2004. Effects of physical activity on intestinal gas transit and evacuation in healthy subjects. Amercian Journal of Medicine 116: 536-539. https://doi.org/10.1016/j.amjmed.2003.12.018
D’Angelo, S., 2020. Polyphenols: potential beneficial effects of these phytochemicals in athletes. Current Sports Medicine Reports 19: 260-265. https://doi.org/10.1249/jsr.0000000000000729
Delezie, J. and Handschin, C., 2018. Endocrine crosstalk between skeletal muscle and the brain. Frontiers in Neurology 9: 698. https://doi.org/10.3389/fneur.2018.00698
Den Besten, G., Bleeker, A., Gerding, A., van Eunen, K., Havinga, R., van Dijk, T.H., Oosterveer, M.H., Jonker, J.W., Groen, A.K., Reijngoud, D.J. and Bakker, B.M., 2015. Short-chain fatty acids protect against high-fat diet-induced obesity via a PPARγ-dependent switch from lipogenesis to fat oxidation. Diabetes 64: 2398-2408. https://doi.org/10.2337/db14-1213
Durk, R.P., Castillo, E., Marquez-Magana, L., Grosicki, G.J., Bolter, N.D., Lee, C.M. and Bagley, J.R., 2019. Gut microbiota composition is related to cardiorespiratory fitness in healthy young adults. International Journal of Sport Nutrition and Exercise Metabolism 29: 249-253. https://doi.org/10.1123/ijsnem.2018-0024
EFSA Panel on Dietetic Products, Nutrition and Allergies, 2012. Scientific opinion on dietary reference values for protein. EFSA Journal 10: 2557. https://doi.org/10.2903/j.efsa.2012.2557
Estaki, M., Pither, J., Baumeister, P., Little, J.P., Gill, S.K., Ghosh, S., Ahmadi-Vand, Z., Marsden, K.R. and Gibson, D.L., 2016. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 4: 42. https://doi.org/10.1186/s40168-016-0189-7
Felizardo, R.J.F., Castoldi, A., Andrade-Oliveira, V. and Câmara, N.O.S., 2016. The microbiota and chronic kidney diseases: a double-edged sword. Clinical and Translational Immunology 5: e86. https://doi.org/10.1038/cti.2016.36
Finaud, J., Lac, G. and Filaire, E., 2006. Oxidative stress. Sports Medicine 36: 327-358. https://doi.org/10.2165/00007256-200636040-00004
Finotello, F., Mastrorilli, E. and Di Camillo, B., 2018. Measuring the diversity of the human microbiota with targeted next-generation sequencing. Briefings in Bioinformatics 19: 679-692. https://doi.org/10.1093/bib/bbw119
Frampton, J., Murphy, K.G., Frost, G. and Chambers, E.S., 2020. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nature Metabolism 2: 840-848. https://doi.org/10.1038/s42255-020-0188-7
Fritsche, K.L., 2015. The science of fatty acids and inflammation. Advances in Nutrition 6: 293S-301S.
Fritz, P., Fritz, R., Bóday, P., Bóday, Á., Bató, E., Kesserű, P. and Oláh, C., 2024. Gut microbiome composition: link between sports performance and protein absorption? Journal of the International Society of Sports Nutrition 21: 2297992. https://doi.org/10.1080/15502783.2023.2297992
Furber, M.J.W., Young, G.R., Holt, G.S., Pyle, S., Davison, G., Roberts, M.G., Roberts, J.D., Howatson, G. and Smith, D.L., 2022. Gut microbial stability is associated with greater endurance performance in athletes undertaking dietary periodization. mSystems 7: e00129-00122. https://doi.org/10.1128/msystems.00129-22
Fushimi, T. and Sato, Y., 2005. Effect of acetic acid feeding on the circadian changes in glycogen and metabolites of glucose and lipid in liver and skeletal muscle of rats. British Journal of Nutrition 94: 714-719. https://doi.org/10.1079/bjn20051545
Fushimi, T., Tayama, K., Fukaya, M., Kitakoshi, K., Nakai, N., Tsukamoto, Y. and Sato, Y., 2001. Acetic acid feeding enhances glycogen repletion in liver and skeletal muscle of rats. Journal of Nutrition 131: 1973-1977. https://doi.org/10.1093/jn/131.7.1973
Galle, F., Valeriani, F., Cattaruzza, M.S., Gianfranceschi, G., Liguori, R., Antinozzi, M., Mederer, B., Liguori, G. and Romano Spica, V., 2020. Mediterranean diet, physical activity and gut microbiome composition: a cross-sectional study among healthy young Italian adults. Nutrients 12: 2164. https://doi.org/10.3390/nu12072164
Gao, R., Tao, Y., Zhou, C., Li, J., Wang, X., Chen, L., Li, F. and Guo, L., 2019. Exercise therapy in patients with constipation: a systematic review and meta-analysis of randomized controlled trials. Scandinavian Journal of Gastroenterology 54: 169-177. https://doi.org/10.1080/00365521.2019.1568544
Gao, Z., Yin, J., Zhang, J., Ward, R.E., Martin, R.J., Lefevre, M., Cefalu, W.T. and Ye, J., 2009. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 58: 1509-1517. https://doi.org/10.2337/db08-1637
Ghosh, N., Das, A., Biswas, N., Gnyawali, S., Singh, K., Gorain, M., Polcyn, C., Khanna, S., Roy, S. and Sen, C.K., 2020. Urolithin A augments angiogenic pathways in skeletal muscle by bolstering NAD+ and SIRT1. Scientific Reports 10: 20184. https://doi.org/10.1038/s41598-020-76564-7
Gibson, G.R., Hutkins, R., Sanders, M.E., Prescott, S.L., Reimer, R.A., Salminen, S.J., Scott, K., Stanton, C., Swanson, K.S., Cani, P.D., Verbeke, K. and Reid, G., 2017. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology and Hepatology 14: 491-502. https://doi.org/10.1038/nrgastro.2017.75
Gomez-Cabrera, M.C., Carretero, A., Millan-Domingo, F., Garcia-Dominguez, E., Correas, A.G., Olaso-Gonzalez, G. and Vina, J., 2021. Redox-related biomarkers in physical exercise. Redox Biology 42: 101956. https://doi.org/10.1016/j.redox.2021.101956
Griffiths, A., Matu, J., Whyte, E., Akin-Nibosun, P., Clifford, T., Stevenson, E. and Shannon, O.M., 2022. The Mediterranean dietary pattern for optimising health and performance in competitive athletes: a narrative review. British Journal of Nutrition 128: 1285-1298. https://doi.org/10.1017/s0007114521003202
Grosicki, G.J., Fielding, R.A. and Lustgarten, M.S., 2018. Gut microbiota contribute to age-related changes in skeletal muscle size, composition, and function: biological basis for a gut-muscle axis. Calcified Tissue International 102: 433-442. https://doi.org/10.1007/s00223-017-0345-5
Han, D.-S., Wu, W.-K., Liu, P.-Y., Yang, Y.-T., Hsu, H.-C., Kuo, C.-H., Wu, M.-S. and Wang, T.-G., 2022. Differences in the gut microbiome and reduced fecal butyrate in elders with low skeletal muscle mass. Clinical Nutrition 41: 1491-1500. https://doi.org/10.1016/j.clnu.2022.05.008
Han, J.H., Kim, I.S., Jung, S.H., Lee, S.G., Son, H.Y. and Myung, C.S., 2014. The effects of propionate and valerate on insulin responsiveness for glucose uptake in 3T3-L1 adipocytes and C2C12 myotubes via G protein-coupled receptor 41. PLoS ONE 9: e95268. https://doi.org/10.1371/journal.pone.0095268
Han, M., Yang, K., Yang, P., Zhong, C., Chen, C., Wang, S., Lu, Q. and Ning, K., 2020. Stratification of athletes’ gut microbiota: the multifaceted hubs associated with dietary factors, physical characteristics and performance. Gut Microbes 12: 1-18. https://doi.org/10.1080/19490976.2020.1842991
He, J., Zhang, P., Shen, L., Niu, L., Tan, Y., Chen, L., Zhao, Y., Bai, L., Hao, X., Li, X., Zhang, S. and Zhu, L., 2020. Short-chain fatty acids and their association with signalling pathways in inflammation, glucose and lipid metabolism. International Journal of Molecular Sciences 21: 6356.
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology and Hepatology 11: 506-514. https://doi.org/10.1038/nrgastro.2014.66
Hintikka, J.E., Munukka, E., Valtonen, M., Luoto, R., Ihalainen, J.K., Kallonen, T., Waris, M., Heinonen, O.J., Ruuskanen, O. and Pekkala, S., 2022. Gut microbiota and serum metabolome in elite cross-country skiers: a controlled study. Metabolites 12: 335. https://doi.org/10.3390/metabo12040335
Holtzman, B. and Ackerman, K.E., 2019. Measurement, determinants, and implications of energy intake in athletes. Nutrients 11: 665. https://doi.org/10.3390/nu11030665
Holzhausen, E.A., Malecki, K.C., Sethi, A.K., Gangnon, R., Cadmus-Bertram, L., Deblois, C.L., Suen, G., Safdar, N. and Peppard, P.E., 2022. Assessing the relationship between physical activity and the gut microbiome in a large, population-based sample of Wisconsin adults. PLoS ONE 17: e0276684. https://doi.org/10.1371/journal.pone.0276684
Hsu, Y.J., Chiu, C.C., Li, Y.P., Huang, W.C., Huang, Y.T., Huang, C.C. and Chuang, H.L., 2015. Effect of intestinal microbiota on exercise performance in mice. Journal of Strength and Conditioning Research 29: 552-558. https://doi.org/10.1519/jsc.0000000000000644
Hu, G.-X., Chen, G.-R., Xu, H., Ge, R.-S. and Lin, J., 2010. Activation of the AMP activated protein kinase by short-chain fatty acids is the main mechanism underlying the beneficial effect of a high fiber diet on the metabolic syndrome. Medical Hypotheses 74: 123-126. https://doi.org/10.1016/j.mehy.2009.07.022
Huang, W.C., Chen, Y.H., Chuang, H.L., Chiu, C.C. and Huang, C.C., 2019. Investigation of the effects of microbiota on exercise physiological adaption, performance, and energy utilization using a gnotobiotic animal model. Front Microbiol 10: 1906. https://doi.org/10.3389/fmicb.2019.01906
Huang, W.C., Pan, C.H., Wei, C.C. and Huang, H.Y., 2020. Lactobacillus plantarum PS128 improves physiological adaptation and performance in triathletes through gut microbiota modulation. Nutrients 12. https://doi.org/10.3390/nu12082315
Hughes, R.L. and Holscher, H.D., 2021. Fueling gut microbes: a review of the interaction between diet, exercise, and the gut microbiota in athletes. Advances in Nutrition 12: 2190-2215. https://doi.org/10.1093/advances/nmab077
Iddrisu, I., Monteagudo-Mera, A., Poveda, C., Pyle, S., Shahzad, M., Andrews, S. and Walton, G.E., 2021. Malnutrition and gut microbiota in children. Nutrients 13: 2727.
Jäger, R., Mohr, A.E., Carpenter, K.C., Kerksick, C.M., Purpura, M., Moussa, A., Townsend, J.R., Lamprecht, M., West, N.P., Black, K., Gleeson, M., Pyne, D.B., Wells, S.D., Arent, S.M., Smith-Ryan, A.E., Kreider, R.B., Campbell, B.I., Bannock, L., Scheiman, J., Wissent, C.J., Pane, M., Kalman, D.S., Pugh, J.N., ter Haar, J.A. and Antonio, J., 2019. International society of sports nutrition position stand: probiotics. Journal of the International Society of Sports Nutrition 16: 62. https://doi.org/10.1186/s12970-019-0329-0
Jager, R., Purpura, M., Stone, J.D., Turner, S.M., Anzalone, A.J., Eimerbrink, M.J., Pane, M., Amoruso, A., Rowlands, D.S. and Oliver, J.M., 2016. Probiotic Streptococcus thermophilus FP4 and Bifidobacterium breve BR03 supplementation attenuates performance and range-of-motion decrements following muscle damaging exercise. Nutrients 8: 642. https://doi.org/10.3390/nu8100642
Jang, L.G., Choi, G., Kim, S.W., Kim, B.Y., Lee, S. and Park, H., 2019. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: an observational study. Journal of the International Society of Sports Nutrition 16: 21. https://doi.org/10.1186/s12970-019-0290-y
Jie, Z., Xia, H., Zhong, S.L., Feng, Q., Li, S., Liang, S., Zhong, H., Liu, Z., Gao, Y., Zhao, H., Zhang, D., Su, Z., Fang, Z., Lan, Z., Li, J., Xiao, L., Li, J., Li, R., Li, X., Li, F., Ren, H., Huang, Y., Peng, Y., Li, G., Wen, B., Dong, B., Chen, J.Y., Geng, Q.S., Zhang, Z.W., Yang, H., Wang, J., Wang, J., Zhang, X., Madsen, L., Brix, S., Ning, G., Xu, X., Liu, X., Hou, Y., Jia, H., He, K. and Kristiansen, K., 2017. The gut microbiome in atherosclerotic cardiovascular disease. Nature Communications 8: 845. https://doi.org/10.1038/s41467-017-00900-1
Judelson, D.A., Maresh, C.M., Anderson, J.M., Armstrong, L.E., Casa, D.J., Kraemer, W.J. and Volek, J.S., 2007. Hydration and muscular performance. Sports Medicine 37: 907-921. https://doi.org/10.2165/00007256-200737100-00006
Jumpertz, R., Le, D.S., Turnbaugh, P.J., Trinidad, C., Bogardus, C., Gordon, J.I. and Krakoff, J., 2011. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. American Journal of Clinical Nutrition 94: 58-65. https://doi.org/10.3945/ajcn.110.010132
Karkman, A., Lehtimaki, J. and Ruokolainen, L., 2017. The ecology of human microbiota: dynamics and diversity in health and disease. Annals of the New York Academy of Science 1399: 78-92. https://doi.org/10.1111/nyas.13326
Kasubuchi, M., Hasegawa, S., Hiramatsu, T., Ichimura, A. and Kimura, I., 2015. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 7: 2839. https://doi.org/10.3390/nu7042839
Kekkonen, R.A., Vasankari, T.J., Vuorimaa, T., Haahtela, T., Julkunen, I. and Korpela, R., 2007. The effect of probiotics on respiratory infections and gastrointestinal symptoms during training in marathon runners. International Journal of Sport Nutrition and Exercise Metabolism 17: 352-363. https://doi.org/10.1123/ijsnem.17.4.352
Kinashi, Y. and Hase, K., 2021. Partners in leaky gut syndrome: intestinal dysbiosis and autoimmunity. Frontiers in Immunology 12: 673708. https://doi.org/10.3389/fimmu.2021.673708
Kocot, A.M., Jarocka-Cyrta, E. and Drabinska, N., 2022. Overview of the importance of biotics in gut barrier integrity. International Journal of Molecular Sciences 23: 2896. https://doi.org/10.3390/ijms23052896
Krajmalnik-Brown, R., Ilhan, Z.-E., Kang, D.-W. and DiBaise, J.K., 2012. Effects of gut microbes on nutrient absorption and energy regulation. Nutrition in Clinical Practice 27: 201-214. https://doi.org/10.1177/0884533611436116
Kristensen, N.B., Bryrup, T., Allin, K.H., Nielsen, T., Hansen, T.H. and Pedersen, O., 2016. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: a systematic review of randomized controlled trials. Genome Medicine 8: 52. https://doi.org/10.1186/s13073-016-0300-5
Kulecka, M., Fraczek, B., Balabas, A., Czarnowski, P., Zeber-Lubecka, N., Zapala, B., Baginska, K., Glowienka, M., Szot, M., Skorko, M., Kluska, A., Piatkowska, M., Mikula, M. and Ostrowski, J., 2023. Characteristics of the gut microbiome in esports players compared with those in physical education students and professional athletes. Frontiers in Nutrition 9: 1092846. https://doi.org/10.3389/fnut.2022.1092846
Kulecka, M., Fraczek, B., Mikula, M., Zeber-Lubecka, N., Karczmarski, J., Paziewska, A., Ambrozkiewicz, F., Jagusztyn-Krynicka, K., Cieszczyk, P. and Ostrowski, J., 2020. The composition and richness of the gut microbiota differentiate the top Polish endurance athletes from sedentary controls. Gut Microbes 11: 1374-1384. https://doi.org/10.1080/19490976.2020.1758009
Kumari, M., Singh, P., Nataraj, B.H., Kokkiligadda, A., Naithani, H., Azmal Ali, S., Behare, P.V. and Nagpal, R., 2021. Fostering next-generation probiotics in human gut by targeted dietary modulation: an emerging perspective. Food Research International 150: 110716. https://doi.org/10.1016/j.foodres.2021.110716
Lahiri, S., Kim, H., Garcia-Perez, I., Reza, M.M., Martin, K.A., Kundu, P., Cox, L.M., Selkrig, J., Posma, J.M., Zhang, H., Padmanabhan, P., Moret, C., Gulyás, B., Blaser, M.J., Auwerx, J., Holmes, E., Nicholson, J., Wahli, W. and Pettersson, S., 2019. The gut microbiota influences skeletal muscle mass and function in mice. Science Translational Medicine 11: 5662. https://doi.org/10.1126/scitranslmed.aan5662
Lam, Y.Y., Ha, C.W.Y., Hoffmann, J.M.A., Oscarsson, J., Dinudom, A., Mather, T.J., Cook, D.I., Hunt, N.H., Caterson, I.D., Holmes, A.J. and Storlien, L.H., 2015. Effects of dietary fat profile on gut permeability and microbiota and their relationships with metabolic changes in mice. Obesity 23: 1429-1439. https://doi.org/10.1002/oby.21122
Lamprecht, M., Bogner, S., Schippinger, G., Steinbauer, K., Fankhauser, F., Hallstroem, S., Schuetz, B. and Greilberger, J.F., 2012. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. Journal of the International Society of Sports Nutrition 9: 45. https://doi.org/10.1186/1550-2783-9-45
Lavefve, L., Howard, L.R. and Carbonero, F., 2020. Berry polyphenols metabolism and impact on human gut microbiota and health. Food and Function 11: 45-65. https://doi.org/10.1039/C9FO01634A
LeBlanc, J.G., Milani, C., de Giori, G.S., Sesma, F., van Sinderen, D. and Ventura, M., 2013. Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Current Opinion in Biotechnology 24: 160-168. https://doi.org/10.1016/j.copbio.2012.08.005
Li, K., Liu, A., Zong, W., Dai, L., Liu, Y., Luo, R., Ge, S. and Dong, G., 2021. Moderate exercise ameliorates osteoarthritis by reducing lipopolysaccharides from gut microbiota in mice. Saudi Journal of Biological Sciences 28: 40-49. https://doi.org/10.1016/j.sjbs.2020.08.027
Li, Y., Cheng, M., Zha, Y., Yang, K., Tong, Y., Wang, S., Lu, Q. and Ning, K., 2023. Gut microbiota and inflammation patterns for specialized athletes: a multi-cohort study across different types of sports. mSystems 8: e0025923. https://doi.org/10.1128/msystems.00259-23
Liang, R., Zhang, S., Peng, X., Yang, W., Xu, Y., Wu, P., Chen, J., Cai, Y. and Zhou, J., 2019. Characteristics of the gut microbiota in professional martial arts athletes: a comparison between different competition levels. PLoS ONE 14: e0226240. https://doi.org/10.1371/journal.pone.0226240
Lin, C.L., Hsu, Y.J., Ho, H.H., Chang, Y.C., Kuo, Y.W., Yeh, Y.T., Tsai, S.Y., Chen, C.W., Chen, J.F., Huang, C.C. and Lee, M.C., 2020. Bifidobacterium longum subsp. longum OLP-01 supplementation during endurance running training improves exercise performance in middle- and long-distance runners: a double-blind controlled trial. Nutrients 12: 1972. https://doi.org/10.3390/nu12071972
Lin, R., Liu, W., Piao, M. and Zhu, H., 2017. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 49: 2083-2090. https://doi.org/10.1007/s00726-017-2493-3
Lira, F.S., Rosa, J.C., Pimentel, G.D., Souza, H.A., Caperuto, E.C., Carnevali Jr., L.C., Seelaender, M., Damaso, A.R., Oyama, L.M., de Mello, M.T. and Santos, R.V., 2010. Endotoxin levels correlate positively with a sedentary lifestyle and negatively with highly trained subjects. Lipids in Health and Disease 9: 82. https://doi.org/10.1186/1476-511x-9-82
Lis, D.M., Kings, D. and Larson-Meyer, D.E., 2019. Dietary practices adopted by track-and-field athletes: gluten-free, low FODMAP, vegetarian, and fasting. International Journal of Sport Nutrition and Exercise Metabolism 29: 236-245. https://doi.org/10.1123/ijsnem.2018-0309
Liu, L., Fu, C. and Li, F., 2019. Acetate affects the process of lipid metabolism in rabbit liver, skeletal muscle and adipose tissue. Animals 9: 799. https://doi.org/10.3390/ani9100799
Liu, Y.C., Li, X.Y. and Shen, L., 2020. Modulation effect of tea consumption on gut microbiota. Appl Microbiol Biotechnol 104: 981-987. https://doi.org/10.1007/s00253-019-10306-2
Louis, P. and Flint, H.J., 2017. Formation of propionate and butyrate by the human colonic microbiota. Environmental Microbiology 19: 29-41. https://doi.org/10.1111/1462-2920.13589
Lozupone, C.A., Stombaugh, J.I., Gordon, J.I., Jansson, J.K. and Knight, R., 2012. Diversity, stability and resilience of the human gut microbiota. Nature 489: 220-230. https://doi.org/10.1038/nature11550
Luk, A., Mitchell, L., Hackett, D., O’Connor, H., Koay, Y., O’Sullivan, J. and Holmes, A., 2022. Intersection of diet and exercise with the gut microbiome and circulating metabolites in male bodybuilders. Metabolites 12: 911. https://doi.org/10.3390/metabo12100911
Luo, B., Xiang, D., Nieman, D.C. and Chen, P., 2014. The effects of moderate exercise on chronic stress-induced intestinal barrier dysfunction and antimicrobial defense. Brain, Behavior, and Immunity 39: 99-106. https://doi.org/10.1016/j.bbi.2013.11.013
Lv, W.Q., Lin, X., Shen, H., Liu, H.M., Qiu, X., Li, B.Y., Shen, W.D., Ge, C.L., Lv, F.Y., Shen, J., Xiao, H.M. and Deng, H.W., 2021. Human gut microbiome impacts skeletal muscle mass via gut microbial synthesis of the short-chain fatty acid butyrate among healthy menopausal women. Journal of Cachexia, Sarcopenia and Muscle 12: 1860-1870. https://doi.org/10.1002/jcsm.12788
Lynch, S.V. and Pedersen, O., 2016. The human intestinal microbiome in health and disease. New England Journal of Medicine 375: 2369-2379. https://doi.org/10.1056/NEJMra1600266
Lynne, V.M., 2014. Use of probiotics to correct dysbiosis of normal microbiota following disease or disruptive events: a systematic review. BMJ Open 4: e005047. https://doi.org/10.1136/bmjopen-2014-005047
Mach, N. and Clark, A., 2017. Micronutrient deficiencies and the human gut microbiota. Trends in Microbiology 25: 607-610.
Magne, F., Gotteland, M., Gauthier, L., Zazueta, A., Pesoa, S., Navarrete, P. and Balamurugan, R., 2020. The Firmicutes/Bacteroidetes ratio: a relevant marker of gut dysbiosis in obese patients? Nutrients 12: 1474. https://doi.org/10.3390/nu12051474
Magnúsdóttir, S., Ravcheev, D., de Crécy-Lagard, V. and Thiele, I., 2015. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Frontiers in Genetics 6: 00148. https://doi.org/10.3389/fgene.2015.00148
Makki, K., Deehan, E.C., Walter, J. and Bäckhed, F., 2018. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 23: 705-715. https://doi.org/10.1016/j.chom.2018.05.012
Mancin, L., Amatori, S., Caprio, M., Sattin, E., Bertoldi, L., Cenci, L., Sisti, D., Bianco, A. and Paoli, A., 2022a. Effect of 30 days of ketogenic Mediterranean diet with phytoextracts on athletes’ gut microbiome composition. Frontiers in Nutrition 9: 979651. https://doi.org/10.3389/fnut.2022.979651
Mancin, L., Wu, G.D. and Paoli, A., 2022b. Gut microbiota-bile acid-skeletal muscle axis. Trends in Microbiology 31: 254-269. https://doi.org/10.1016/j.tim.2022.10.003
Marshall, H., Chrismas, B.C.R., Suckling, C.A., Roberts, J.D., Foster, J. and Taylor, L., 2017. Chronic probiotic supplementation with or without glutamine does not influence the eHsp72 response to a multi-day ultra-endurance exercise event. Applied Physiology, Nutrition, and Metabolism 42: 876-883. https://doi.org/10.1139/apnm-2017-0131
Martarelli, D., Verdenelli, M.C., Scuri, S., Cocchioni, M., Silvi, S., Cecchini, C. and Pompei, P., 2011. Effect of a probiotic intake on oxidant and antioxidant parameters in plasma of athletes during intense exercise training. Current Microbiology 62: 1689-1696. https://doi.org/10.1007/s00284-011-9915-3
Martin-Gallausiaux, C., Marinelli, L., Blottière, H.M., Larraufie, P. and Lapaque, N., 2021. SCFA: mechanisms and functional importance in the gut. Proceedings of the Nutrition Society 80: 37-49. https://doi.org/10.1017/s0029665120006916
Martino, C., Dilmore, A.H., Burcham, Z.M., Metcalf, J.L., Jeste, D. and Knight, R., 2022. Microbiota succession throughout life from the cradle to the grave. Nature Reviews Microbiology 20: 707-720. https://doi.org/10.1038/s41579-022-00768-z
Maruta, H., Yoshimura, Y., Araki, A., Kimoto, M., Takahashi, Y. and Yamashita, H., 2016. Activation of AMP-activated protein kinase and stimulation of energy metabolism by acetic acid in L6 myotube cells. PLoS ONE 11: e0158055. https://doi.org/10.1371/journal.pone.0158055
Maughan, R.J., Burke, L.M., Dvorak, J., Larson-Meyer, D.E., Peeling, P., Phillips, S.M., Rawson, E.S., Walsh, N.P., Garthe, I., Geyer, H., Meeusen, R., van Loon, L.J.C., Shirreffs, S.M., Spriet, L.L., Stuart, M., Vernec, A., Currell, K., Ali, V.M., Budgett, R.G., Ljungqvist, A., Mountjoy, M., Pitsiladis, Y.P., Soligard, T., Erdener, U. and Engebretsen, L., 2018. IOC consensus statement: dietary supplements and the high-performance athlete. British Journal of Sports Medicine 52: 439-455. https://doi.org/10.1136/bjsports-2018-099027
Mazur-Kurach, P., Fraczek, B. and Klimek, A.T., 2022. Does multi-strain probiotic supplementation impact the effort capacity of competitive road cyclists? International Journal of Environmental Research and Public Health 19: 12205. https://doi.org/10.3390/ijerph191912205
McKenna, C.F., Salvador, A.F., Hughes, R.L., Scaroni, S.E., Alamilla, R.A., Askow, A.T., Paluska, S.A., Dilger, A.C., Holscher, H.D., De Lisio, M., Khan, N.A. and Burd, N.A., 2021. Higher protein intake during resistance training does not potentiate strength, but modulates gut microbiota, in middle-aged adults: a randomized control trial. American Journal of Physiology – Endocrinology and Metabolism 320: E900-E913. https://doi.org/10.1152/ajpendo.00574.2020
McSwiney, F.T., Doyle, L., Plews, D.J. and Zinn, C., 2019. Impact of ketogenic diet on athletes: current insights. Open Access Journal of Sports Medicine 10: 171-183. https://doi.org/10.2147/oajsm.s180409
Michalickova, D., Minic, R., Dikic, N., Andjelkovic, M., Kostic-Vucicevic, M., Stojmenovic, T., Nikolic, I. and Djordjevic, B., 2016. Lactobacillus helveticus Lafti L10 supplementation reduces respiratory infection duration in a cohort of elite athletes: a randomized, double-blind, placebo-controlled trial. Applied Physiology, Nutrition, and Metabolism 41: 782-789. https://doi.org/10.1139/apnm-2015-0541%M27363733
Miles, M.P., 2020. Probiotics and gut health in athletes. Current Nutrition Reports 9: 129-136. https://doi.org/10.1007/s13668-020-00316-2
Mishra, V., Shah, C., Mokashe, N., Chavan, R., Yadav, H. and Prajapati, J., 2015. Probiotics as potential antioxidants: a systematic review. Journal of Agricultural and Food Chemistry 63: 3615-3626. https://doi.org/10.1021/jf506326t
Mohr, A.E., Jager, R., Carpenter, K.C., Kerksick, C.M., Purpura, M., Townsend, J.R., West, N.P., Black, K., Gleeson, M., Pyne, D.B., Wells, S.D., Arent, S.M., Kreider, R.B., Campbell, B.I., Bannock, L., Scheiman, J., Wissent, C.J., Pane, M., Kalman, D.S., Pugh, J.N., Ortega-Santos, C.P., Ter Haar, J.A., Arciero, P.J. and Antonio, J., 2020. The athletic gut microbiota. Journal of the International Society of Sports Nutrition 17: 24. https://doi.org/10.1186/s12970-020-00353-w
Moitinho-Silva, L., Wegener, M., May, S., Schrinner, F., Akhtar, A., Boysen, T.J., Schaeffer, E., Hansen, C., Schmidt, T., Ruhlemann, M.C., Hubenthal, M., Rausch, P., Kondakci, M.T., Maetzler, W., Weidinger, S., Laudes, M., Suss, P., Schulte, D., Junker, R., Sommer, F., Weisser, B., Bang, C. and Franke, A., 2021. Short-term physical exercise impacts on the human holobiont obtained by a randomised intervention study. BMC Microbiology 21: 162. https://doi.org/10.1186/s12866-021-02214-1
Moreno-Perez, D., Bressa, C., Bailen, M., Hamed-Bousdar, S., Naclerio, F., Carmona, M., Perez, M., Gonzalez-Soltero, R., Montalvo-Lominchar, M.G., Carabana, C. and Larrosa, M., 2018. Effect of a protein supplement on the gut microbiota of endurance athletes: a randomized, controlled, double-blind pilot study. Nutrients 10: 337. https://doi.org/10.3390/nu10030337
Mörkl, S., Lackner, S., Müller, W., Gorkiewicz, G., Kashofer, K., Oberascher, A., Painold, A., Holl, A., Holzer, P., Meinitzer, A., Mangge, H. and Holasek, S., 2017. Gut microbiota and body composition in anorexia nervosa inpatients in comparison to athletes, overweight, obese, and normal weight controls. International Journal of Eating Disorders 50: 1421-1431. https://doi.org/10.1002/eat.22801
Munukka, E., Ahtiainen, J.P., Puigbo, P., Jalkanen, S., Pahkala, K., Keskitalo, A., Kujala, U.M., Pietila, S., Hollmen, M., Elo, L., Huovinen, P., D’Auria, G. and Pekkala, S., 2018. Six-week endurance exercise alters gut metagenome that is not reflected in systemic metabolism in over-weight women. Frontiers in Microbiology 9: 2323. https://doi.org/10.3389/fmicb.2018.02323
Murtaza, N., Burke, L., Vlahovich, N., Charlesson, B., O’ Neill, H., Ross, M., Campbell, K., Krause, L. and Morrison, M., 2019. The effects of dietary pattern during intensified training on stool microbiota of elite race walkers. Nutrients 11: 261. https://doi.org/10.3390/nu11020261
Nakao, C., Yamada, E., Fukaya, M., Tayama, K., Tsukamoto, Y. and Sato, Y., 2001. Effect of acetate on glycogen replenishment in liver and skeletal muscles after exhaustive swimming in rats. Scandinavian Journal of Medicine and Science in Sports 11: 33-37. https://doi.org/10.1034/j.1600-0838.2001.011001033.x
Nay, K., Jollet, M., Goustard, B., Baati, N., Vernus, B., Pontones, M., Lefeuvre-Orfila, L., Bendavid, C., Rué, O., Mariadassou, M., Bonnieu, A., Ollendorff, V., Lepage, P., Derbré, F. and Koechlin-Ramonatxo, C., 2019. Gut bacteria are critical for optimal muscle function: a potential link with glucose homeostasis. American Journal of Physiology – Endocrinology and Metabolism 317: E158-e171. https://doi.org/10.1152/ajpendo.00521.2018
Nilsson, N.E., Kotarsky, K., Owman, C. and Olde, B., 2003. Identification of a free fatty acid receptor, FFA2R, expressed on leukocytes and activated by short-chain fatty acids. Biochemical and Biophysical Research Communications 303: 1047-1052. https://doi.org/10.1016/S0006-291X(03)00488-1
O’Brien, M.T., O’Sullivan, O., Claesson, M.J. and Cotter, P.D., 2022. The athlete gut microbiome and its relevance to health and performance: a review. Sports Medicine 52: 119-128. https://doi.org/10.1007/s40279-022-01785-x
O’Donovan, C.M., Madigan, S.M., Garcia-Perez, I., Rankin, A., O’Sullivan, O. and Cotter, P.D., 2020. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. Journal of Science and Medicine in Sport 23: 63-68. https://doi.org/10.1016/j.jsams.2019.08.290
Oettlé, G.J., 1991. Effect of moderate exercise on bowel habit. Gut 32: 941-944. https://doi.org/10.1136/gut.32.8.941
Okamoto, T., Morino, K., Ugi, S., Nakagawa, F., Lemecha, M., Ida, S., Ohashi, N., Sato, D., Fujita, Y. and Maegawa, H., 2019. Microbiome potentiates endurance exercise through intestinal acetate production. American Journal of Physiology – Endocrinology and Metabolism 316: E956-e966. https://doi.org/10.1152/ajpendo.00510.2018
Oliveira, C.B., Marques, C., Abreu, R., Figueiredo, P., Calhau, C., Brito, J. and Sousa, M., 2022. Gut microbiota of elite female football players is not altered during an official international tournament. Scandinavian Journal of Medicine and Science in Sports 32 Suppl 1: 62-72. https://doi.org/10.1111/sms.14096
Oren, A. and Garrity, G.M., 2021. Valid publication of the names of forty-two phyla of prokaryotes. International Journal of Systematic and Evolutionary Microbiology 71: 56. https://doi.org/10.1099/ijsem.0.005056
Pan, J.H., Kim, J.H., Kim, H.M., Lee, E.S., Shin, D.-H., Kim, S., Shin, M., Kim, S.H., Lee, J.H. and Kim, Y.J., 2015. Acetic acid enhances endurance capacity of exercise-trained mice by increasing skeletal muscle oxidative properties. Bioscience, Biotechnology, and Biochemistry 79: 1535-1541. https://doi.org/10.1080/09168451.2015.1034652
Panzitt, K. and Wagner, M., 2021. FXR in liver physiology: multiple faces to regulate liver metabolism. Biochimica et Biophysica Acta (BBA) – Molecular Basis of Disease 1867: 166133. https://doi.org/10.1016/j.bbadis.2021.166133
Paoli, A., Mancin, L., Bianco, A., Thomas, E., Mota, J.F. and Piccini, F., 2019. Ketogenic diet and microbiota: friends or enemies? Genes 10: 534. https://doi.org/10.3390/genes10070534
Pedersen, B.K. and Febbraio, M.A., 2008. Muscle as an endocrine organ: focus on muscle-derived interleukin-6. Physiological Reviews 88: 1379-1406. https://doi.org/10.1152/physrev.90100.2007
Pedersen, B.K., Rohde, T. and Zacho, M., 1996. Immunity in athletes. Journal of Sports Medicine and Physical Fitness 36: 236-245.
Peeling, P., Sim, M. and McKay, A.K.A., 2023. Considerations for the consumption of vitamin and mineral supplements in athlete populations. Sports Medicine 53: 15-24. https://doi.org/10.1007/s40279-023-01875-4
Pendergast, D.R., Leddy, J.J. and Venkatraman, J.T., 2000. A perspective on fat intake in athletes. Journal of the American College of Nutrition 19: 345-350. https://doi.org/10.1080/07315724.2000.10718930
Pensgaard, A.M. and Ursin, H., 1998. Stress, control, and coping in elite athletes. Scandinavian Journal of Medicine and Science in Sports 8: 183-189. https://doi.org/10.1111/j.1600-0838.1998.tb00190.x
Petersen, L.M., Bautista, E.J., Nguyen, H., Hanson, B.M., Chen, L., Lek, S.H., Sodergren, E. and Weinstock, G.M., 2017. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 5: 98. https://doi.org/10.1186/s40168-017-0320-4
Portune, K.J., Beaumont, M., Davila, A.-M., Tomé, D., Blachier, F. and Sanz, Y., 2016. Gut microbiota role in dietary protein metabolism and health-related outcomes: the two sides of the coin. Trends in Food Science and Technology 57: 213-232. https://doi.org/10.1016/j.tifs.2016.08.011
Przewlocka, K., Folwarski, M., Kazmierczak-Siedlecka, K., Skonieczna-Zydecka, K. and Kaczor, J.J., 2020. Gut-muscle axisexists and may affect skeletal muscle adaptation to training. Nutrients 12: 1451. https://doi.org/10.3390/nu12051451
Pugh, J.N., Sparks, A.S., Doran, D.A., Fleming, S.C., Langan-Evans, C., Kirk, B., Fearn, R., Morton, J.P. and Close, G.L., 2019. Four weeks of probiotic supplementation reduces GI symptoms during a marathon race. European Journal of Applied Physiology 119: 1491-1501. https://doi.org/10.1007/s00421-019-04136-3
Pumpa, K.L., McKune, A.J. and Harnett, J., 2019. A novel role of probiotics in improving host defence of elite rugby union athlete: a double blind randomised controlled trial. Journal of Science and Medicine in Sport 22: 876-881. https://doi.org/10.1016/j.jsams.2019.03.013
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K.S., Manichanh, C., Nielsen, T., Pons, N., Levenez, F., Yamada, T., Mende, D.R., Li, J., Xu, J., Li, S., Li, D., Cao, J., Wang, B., Liang, H., Zheng, H., Xie, Y., Tap, J., Lepage, P., Bertalan, M., Batto, J.M., Hansen, T., Le Paslier, D., Linneberg, A., Nielsen, H.B., Pelletier, E., Renault, P., Sicheritz-Ponten, T., Turner, K., Zhu, H., Yu, C., Li, S., Jian, M., Zhou, Y., Li, Y., Zhang, X., Li, S., Qin, N., Yang, H., Wang, J., Brunak, S., Dore, J., Guarner, F., Kristiansen, K., Pedersen, O., Parkhill, J., Weissenbach, J., Meta, H.I.T.C., Bork, P., Ehrlich, S.D. and Wang, J., 2010. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464: 59-65. https://doi.org/10.1038/nature08821
Qiu, Y., Yu, J., Li, Y., Yang, F., Yu, H., Xue, M., Zhang, F., Jiang, X., Ji, X. and Bao, Z., 2021. Depletion of gut microbiota induces skeletal muscle atrophy by FXR-FGF15/19 signalling. Annals of Medicine 53: 508-522. https://doi.org/10.1080/07853890.2021.1900593
Quero, C.D., Manonelles, P., Fernandez, M., Abellan-Aynes, O., Lopez-Plaza, D., Andreu-Caravaca, L., Hinchado, M.D., Galvez, I. and Ortega, E., 2021. Differential health effects on inflammatory, immunological and stress parameters in professional soccer players and sedentary individuals after consuming a synbiotic. A triple-blinded, randomized, placebo-controlled pilot study. Nutrients 13: 1321. https://doi.org/10.3390/nu13041321
Radzikowska, U., Rinaldi, A., Sözener, Z., Karaguzel, D., Wojcik, M., Cypryk, K., Akdis, M., Akdis, C. and Sokolowska, M., 2019. The influence of dietary fatty acids on immune responses. Nutrients 11: 2990. https://doi.org/10.3390/nu11122990
Resende, A.S., Leite, G.S.F. and Lancha Junior, A.H., 2021. Changes in the gut bacteria composition of healthy men with the same nutritional profile undergoing 10-week aerobic exercise training: a randomized controlled trial. Nutrients 13: 2839. https://doi.org/10.3390/nu13082839
Ribeiro, F.M., Petriz, B., Marques, G., Kamilla, L.H. and Franco, O.L., 2021. Is there an exercise-intensity threshold capable of avoiding the leaky gut? Frontiers in Nutrition 8: 627289. https://doi.org/10.3389/fnut.2021.627289
Rios-Covian, D., Ruas-Madiedo, P., Margolles, A., Gueimonde, M., de Los Reyes-Gavilan, C.G. and Salazar, N., 2016. Intestinal short chain fatty acids and their link with diet and human health. Frontiers in Microbiology 7: 185. https://doi.org/10.3389/fmicb.2016.00185
Roberts, J.D., Suckling, C.A., Peedle, G.Y., Murphy, J.A., Dawkins, T.G. and Roberts, M.G., 2016. An exploratory investigation of endotoxin levels in novice long distance triathletes, and the effects of a multi-strain probiotic/prebiotic, antioxidant intervention. Nutrients 8: 733. https://doi.org/10.3390/nu8110733
Robertson, R.C., Seira Oriach, C., Murphy, K., Moloney, G.M., Cryan, J.F., Dinan, T.G., Ross, R.P. and Stanton, C., 2017. Deficiency of essential dietary n-3 PUFA disrupts the caecal microbiome and metabolome in mice. British Journal of Nutrition 118: 959-970. https://doi.org/10.1017/S0007114517002999
Rodriguez, N.R., DiMarco, N.M. and Langley, S., 2009. Position of the American Dietetic Association, Dietitians of Canada, and the American College of Sports Medicine: nutrition and athletic performance. Journal of the American Dietetic Association 109: 509-527.
Sakakibara, S., Yamauchi, T., Oshima, Y., Tsukamoto, Y. and Kadowaki, T., 2006. Acetic acid activates hepatic AMPK and reduces hyperglycemia in diabetic KK-A(y) mice. Biochemical and Biophysical Research Communications 344: 597-604. https://doi.org/10.1016/j.bbrc.2006.03.176
Salminen, S., Collado, M.C., Endo, A., Hill, C., Lebeer, S., Quigley, E.M.M., Sanders, M.E., Shamir, R., Swann, J.R., Szajewska, H. and Vinderola, G., 2021. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nature Reviews Gastroenterology and Hepatology 18: 649-667. https://doi.org/10.1038/s41575-021-00440-6
Sasaki, T., Kuboyama, A., Mita, M., Murata, S., Shimizu, M., Inoue, J., Mori, K. and Sato, R., 2018. The exercise-inducible bile acid receptor Tgr5 improves skeletal muscle function in mice. Journal of Biological Chemistry 293: 10322-10332. https://doi.org/10.1074/jbc.RA118.002733
Scarpellini, E., Ianiro, G., Attili, F., Bassanelli, C., De Santis, A. and Gasbarrini, A., 2015. The human gut microbiota and virome: potential therapeutic implications. Digestive and Liver Disease 47: 1007-1012. https://doi.org/10.1016/j.dld.2015.07.008
Scheiman, J., Luber, J.M., Chavkin, T.A., MacDonald, T., Tung, A., Pham, L.D., Wibowo, M.C., Wurth, R.C., Punthambaker, S., Tierney, B.T., Yang, Z., Hattab, M.W., Avila-Pacheco, J., Clish, C.B., Lessard, S., Church, G.M. and Kostic, A.D., 2019. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nature Medicine 25: 1104-1109. https://doi.org/10.1038/s41591-019-0485-4
Schonke, M., Martinez-Tellez, B. and Rensen, P.C., 2020. Role of the endocannabinoid system in the regulation of the skeletal muscle response to exercise. Current Opinion in Pharmacology 52: 52-60. https://doi.org/10.1016/j.coph.2020.05.003
Sender, R., Fuchs, S. and Milo, R., 2016. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell 164: 337-340. https://doi.org/10.1016/j.cell.2016.01.013
Shah, S., Mu, C., Moossavi, S., Shen-Tu, G., Schlicht, K., Rohmann, N., Geisler, C., Laudes, M., Franke, A., Züllig, T., Köfeler, H. and Shearer, J., 2023. Physical activity-induced alterations of the gut microbiota are BMI dependent. FASEB Journal 37: e22882. https://doi.org/10.1096/fj.202201571R
Shin, N.R., Whon, T.W. and Bae, J.W., 2015. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends in Biotechnology 33: 496-503. https://doi.org/10.1016/j.tibtech.2015.06.011
Shing, C.M., Peake, J.M., Lim, C.L., Briskey, D., Walsh, N.P., Fortes, M.B., Ahuja, K.D.K. and Vitetta, L., 2014. Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. European Journal of Applied Physiology 114: 93-103. https://doi.org/10.1007/s00421-013-2748-y
Silva, J.R., Rumpf, M.C., Hertzog, M., Castagna, C., Farooq, A., Girard, O. and Hader, K., 2018. Acute and residual soccer match-related fatigue: a systematic review and meta-analysis. Sports Medicine 48: 539-583. https://doi.org/10.1007/s40279-017-0798-8
Simpson, R.J., Kunz, H., Agha, N. and Graff, R., 2015. Exercise and the regulation of immune functions. Progress in Molecular Biology and Translational Science 135: 355-380. https://doi.org/10.1016/bs.pmbts.2015.08.001
Sniffen, J.C., McFarland, L.V., Evans, C.T. and Goldstein, E.J.C., 2018. Choosing an appropriate probiotic product for your patient: an evidence-based practical guide. PLoS ONE 13: e0209205. https://doi.org/10.1371/journal.pone.0209205
Stojanov, S., Berlec, A. and Štrukelj, B., 2020. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 8: 1715.
Strasser, B., Geiger, D., Schauer, M., Gostner, J.M., Gatterer, H., Burtscher, M. and Fuchs, D., 2016. Probiotic supplements beneficially affect tryptophan-kynurenine metabolism and reduce the incidence of upper respiratory tract infections in trained athletes: a randomized, double-blinded, placebo-controlled trial. Nutrients 8: 752. https://doi.org/10.3390/nu8110752
Su, X., 2021. Elucidating the beta-diversity of the microbiome: from global alignment to local alignment. mSystems 6: 00363-00321. https://doi.org/10.1128/msystems.00363-21
Swanson, K.S., Gibson, G.R., Hutkins, R., Reimer, R.A., Reid, G., Verbeke, K., Scott, K.P., Holscher, H.D., Azad, M.B., Delzenne, N.M. and Sanders, M.E., 2020. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nature Reviews Gastroenterology and Hepatology 17: 687-701. https://doi.org/10.1038/s41575-020-0344-2
Tabone, M., Bressa, C., Garcia-Merino, J.A., Moreno-Perez, D., Van, E.C., Castelli, F.A., Fenaille, F. and Larrosa, M., 2021. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Science Reports 11: 3558. https://doi.org/10.1038/s41598-021-82947-1
Tabone, M., Garcia-Merino, J.A., Bressa, C., Rocha Guzman, N.E., Herrera Rocha, K., Chu Van, E., Castelli, F.A., Fenaille, F. and Larrosa, M., 2022. Chronic consumption of cocoa rich in procyanidins has a marginal impact on gut microbiota and on serum and fecal metabolomes in male endurance athletes. Journal of Agricultural and Food Chemistry 70: 1878-1889. https://doi.org/10.1021/acs.jafc.1c07547
Thomas, D.T., Erdman, K.A. and Burke, L.M., 2016. Nutrition and athletic performance. Medicine and Science in Sports and Exercise 48: 543-568.
Tottey, W., Feria-Gervasio, D., Gaci, N., Laillet, B., Pujos, E., Martin, J.F., Sebedio, J.L., Sion, B., Jarrige, J.F., Alric, M. and Brugère, J.F., 2017. Colonic transit time is a driven force of the gut microbiota composition and metabolism: in vitro evidence. Journal of Neurogastroenterology and Motility 23: 124-134. https://doi.org/10.5056/jnm16042
Townsend, J.R., Bender, D., Vantrease, W.C., Sapp, P.A., Toy, A.M., Woods, C.A. and Johnson, K.D., 2018. Effects of probiotic (Bacillus subtilis DE111) supplementation on immune function, hormonal status, and physical performance in division I baseball players. Sports 6: 70. https://doi.org/10.3390/sports6030070
Turnbaugh, P.J., Hamady, M., Yatsunenko, T., Cantarel, B.L., Duncan, A., Ley, R.E., Sogin, M.L., Jones, W.J., Roe, B.A., Affourtit, J.P., Egholm, M., Henrissat, B., Heath, A.C., Knight, R. and Gordon, J.I., 2009. A core gut microbiome in obese and lean twins. Nature 457: 480-484. https://doi.org/10.1038/nature07540
Vaisberg, M., Paixao, V., Almeida, E.B., Santos, J.M.B., Foster, R., Rossi, M., Pithon-Curi, T.C., Gorjao, R., Momesso, C.M., Andrade, M.S., Araujo, J.R., Garcia, M.C., Cohen, M., Perez, E.C., Santos-Dias, A., Vieira, R.P. and Bachi, A.L.L., 2019. Daily intake of fermented milk containing Lactobacillus casei Shirota (Lcs) modulates systemic and upper airways immune/inflammatory responses in marathon runners. Nutrients 11: 1678. https://doi.org/10.3390/nu11071678
Valimaki, I.A., Vuorimaa, T., Ahotupa, M., Kekkonen, R., Korpela, R. and Vasankari, T., 2012. Decreased training volume and increased carbohydrate intake increases oxidized LDL levels. International Journal of Sports Medicine 33: 291-296. https://doi.org/10.1055/s-0031-1291223
Van Wijck, K., Lenaerts, K., van Loon, L.J., Peters, W.H., Buurman, W.A. and Dejong, C.H., 2011. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 6: e22366. https://doi.org/10.1371/journal.pone.0022366
Viciani, E., Barone, M., Bongiovanni, T., Quercia, S., Di Gesu, R., Pasta, G., Manetti, P., Iaia, F.M., Trecroci, A., Rampelli, S., Candela, M., Biagi, E. and Castagnetti, A., 2022. Fecal microbiota monitoring in elite soccer players along the 2019-2020 competitive season. International Journal of Sports Medicine 43: 1137-1147. https://doi.org/10.1055/a-1858-1810
Walczak, R. and Tontonoz, P., 2002. PPARadigms and PPARadoxes: expanding roles for PPARγ in the control of lipid metabolism. Journal of Lipid Research 43: 177-186.
Walsh, M.E., Bhattacharya, A., Sataranatarajan, K., Qaisar, R., Sloane, L., Rahman, M.M., Kinter, M. and Van Remmen, H., 2015. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 14: 957-970. https://doi.org/10.1111/acel.12387
Wastyk, H.C., Fragiadakis, G.K., Perelman, D., Dahan, D., Merrill, B.D., Yu, F.B., Topf, M., Gonzalez, C.G., Van Treuren, W., Han, S., Robinson, J.L., Elias, J.E., Sonnenburg, E.D., Gardner, C.D. and Sonnenburg, J.L., 2021. Gut-microbiota-targeted diets modulate human immune status. Cell 184: 4137-4153 .e4114. https://doi.org/10.1016/j.cell.2021.06.019
Wei, L., Singh, R., Ro, S. and Ghoshal, U.C., 2021. Gut microbiota dysbiosis in functional gastrointestinal disorders: underpinning the symptoms and pathophysiology. JGH Open 5: 976-987. https://doi.org/10.1002/jgh3.12528
West, N.P., Pyne, D.B., Cripps, A.W., Hopkins, W.G., Eskesen, D.C., Jairath, A., Christophersen, C.T., Conlon, M.A. and Fricker, P.A., 2011. Lactobacillus fermentum (PCC®) supplementation and gastrointestinal and respiratory-tract illness symptoms: a randomised control trial in athletes. Nutrition Journal 10: 30. https://doi.org/10.1186/1475-2891-10-30
Whisner, C.M., Maldonado, J., Dente, B., Krajmalnik-Brown, R. and Bruening, M., 2018. Diet, physical activity and screen time but not body mass index are associated with the gut microbiome of a diverse cohort of college students living in university housing: a cross-sectional study. BMC Microbiology 18: 210. https://doi.org/10.1186/s12866-018-1362-x
Willis, N.B., Munoz, C.X., Mysonhimer, A.R., Edwards, C.G., Wolf, P.G., Hillman, C.H., Burd, N.A., Holscher, H.D. and Khan, N.A., 2021. Hydration biomarkers are related to the differential abundance of fecal microbiota and plasma lipopolysaccharide-binding protein in adults. Annals of Nutrition and Metabolism 77 Suppl 4: 37-45. https://doi.org/10.1159/000520478
Wolters, M., Ahrens, J., Romani-Perez, M., Watkins, C., Sanz, Y., Benitez-Paez, A., Stanton, C. and Gunther, K., 2019. Dietary fat, the gut microbiota, and metabolic health – a systematic review conducted within the MyNewGut project. Clinical Nutrition 38: 2504-2520. https://doi.org/10.1016/j.clnu.2018.12.024
Wu, S., Bhat, Z.F., Gounder, R.S., Mohamed Ahmed, I.A., Al-Juhaimi, F.Y., Ding, Y. and Bekhit, A.E.-D.A., 2022. Effect of dietary protein and processing on gut microbiota – a systematic review. Nutrients 14: 453.
Wu, X., Ma, C., Han, L., Nawaz, M., Gao, F., Zhang, X., Yu, P., Zhao, C., Li, L., Zhou, A., Wang, J., Moore, J.E., Millar, B.C. and Xu, J., 2010. Molecular characterisation of the faecal microbiota in patients with type II diabetes. Current Microbiology 61: 69-78. https://doi.org/10.1007/s00284-010-9582-9
Xu, Y., Zhong, F., Zheng, X., Lai, H.Y., Wu, C. and Huang, C., 2022. Disparity of gut microbiota composition among elite athletes and young adults with different physical activity independent of dietary status: a matching study. Frontiers in Nutrition 9: 843076. https://doi.org/10.3389/fnut.2022.843076
Yamashita, H., Maruta, H., Jozuka, M., Kimura, R., Iwabuchi, H., Yamato, M., Saito, T., Fujisawa, K., Takahashi, Y., Kimoto, M., Hiemori, M. and Tsuji, H., 2009. Effects of acetate on lipid metabolism in muscles and adipose tissues of type 2 diabetic Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Bioscience, Biotechnology, and Biochemistry 73: 570-576. https://doi.org/10.1271/bbb.80634
Yang, S., Li, X., Yang, F., Zhao, R., Pan, X., Liang, J., Tian, L., Li, X., Liu, L., Xing, Y. and Wu, M., 2019. Gut microbiota-dependent marker TMAO in promoting cardiovascular disease: inflammation mechanism, clinical prognostic, and potential as a therapeutic target. Frontiers in Pharmacology 10: 1360. https://doi.org/10.3389/fphar.2019.01360
Yang, Y., Shi, Y., Wiklund, P., Tan, X., Wu, N., Zhang, X., Tikkanen, O., Zhang, C., Munukka, E. and Cheng, S., 2017. The association between cardiorespiratory fitness and gut microbiota composition in premenopausal women. Nutrients 9: 792. https://doi.org/10.3390/nu9080792
Yoo, J.Y., Groer, M., Dutra, S.V.O., Sarkar, A. and McSkimming, D.I., 2020. Gut microbiota and immune system interactions. Microorganisms 8: 1587.
Zhai, L., Wu, J., Lam, Y.Y., Kwan, H.Y., Bian, Z.-X. and Wong, H.L.X., 2021. Gut-microbial metabolites, probiotics and their roles in type 2 diabetes. International Journal of Molecular Sciences 22: 12846.
Zhang, S., Wang, R., Li, D., Zhao, L. and Zhu, L., 2021. Role of gut microbiota in functional constipation. Gastroenterology Report 9: 392-401. https://doi.org/10.1093/gastro/goab035



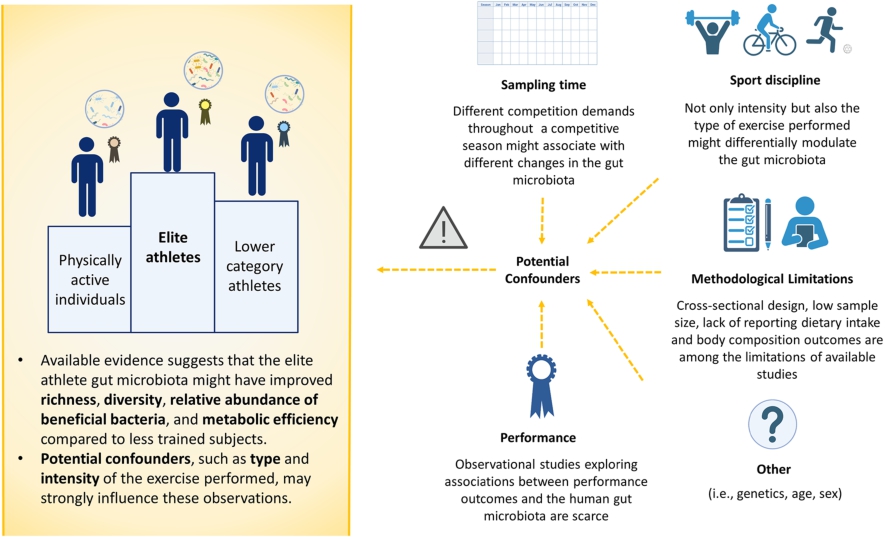



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}