Abstract
Enterococcus faecium SF68 (SF68) is a well-known probiotic with a long history of safe use. Recent changes in the taxonomy of enterococci have shown that a novel species, Enterococcus lactis, is closely related with E. faecium and occurs together with other enterococci in a phylogenetically well-defined E. faecium species group. The close phylogenetic relationship between the species E. faecium and E. lactis prompted a closer investigation into the taxonomic status of E. faecium SF68. Using phylogenomics and ANI, the taxonomic analysis in this study showed that probiotic E. faecium SF68, when compared to other E. faecium and E. lactis type and reference strains, could be re-classified as belonging to the species E. lactis. Further investigations into the functional properties of SF68 showed that it is potentially capable of bacteriocin production, as a bacteriocin gene cluster encoding the leaderless bacteriocin EntK1 together with putative Lactococcus lactis bacteriocins LsbA, and LsbB-like putative immunity peptide (LmrB) were found located in an operon on plasmid pF9. However, bacteriocin expression was not studied. Competitive exclusion experiments in co-culture over 7 days at 37 °C showed that the probiotic SF68 could inhibit the growth of specific E. faecium and Listeria monocytogenes strains, while showing little or no inhibitory activity towards an entero-invasive Escherichia coli and a Salmonella Typhimurium strain, respectively. In cell culture experiments with colon carcinoma HT29 cells, the probiotic SF68 was also able to strain-specifically inhibit adhesion and/or invasion of enterococcal and L. monocytogenes strains, while such adhesion and invasion inhibition effects were less pronounced for E. coli and Salmonella strains. This study therefore provides novel data on the taxonomy and functional properties of SF68, which can be reclassified as Enterococcus lactis SF68, thereby enhancing the understanding of its probiotic nature.
1 Introduction
Enterococcus faecium is a lactic acid bacterium (LAB) occurring in a wide variety of environments, probably because of its ability to tolerate a wide range of pH and temperatures, in addition to its capacity to metabolise a wide spectrum of carbohydrates (Franz et al., 1999; Wessels et al., 1990). Some strains of E. faecium are important as starter cultures for fermented foods and feeds, while others have been used for many years as human and animal probiotics (Franz et al., 2011; Hanchi et al., 2018; Im et al., 2023). E. faecium occurs in the gut of humans and animals (Devriese and Pot, 1995; Devriese et al., 1994; Leclercq et al., 1996) and some strains were shown to cause nosocomial infections such as bacteraemia and endocarditis (Chenoweth and Schaberg, 1990; Im et al., 2023; Morrison et al., 1997; Murray, 1990; Selleck et al., 2019).
Currently there are 61 known Enterococcus species organised in so-called ‘species groups’ of genetically closely related species. The E. faecium species group contains the related species Enterococcus faecium, Enterococcus canis, Enterococcus durans, Enterococcus hirae, Enterococcus mundtii, Enterococcus phoeniculicola, Enterococcus ratti, Enterococcus villorum and Enterococcus thailandicus (Svec and Franz, 2014). Morandi et al. (2012) described a novel species named Enterococcus lactis showing a high degree of relatedness to E. faecium, E. durans and E. hirae and thus also considered to belong to the E. faecium group.
Since many years, a certain degree of genetic heterogeneity has been noted for the species E. faecium. Already in 2002, a study by Vancanneyt et al. (2002) showed the presence of two genomic subgroups within the species. Interestingly, all human clinical isolates or strains showing vancomycin resistance or haemolytic activity were located in genomic subgroup I which also contained the E. faecium type strain LMG 11423T (= DSM 20477T). Subsequently, with the emergence of more powerful, genome-level investigations a more detailed picture of the intra-species population structure of E. faecium emerged. Accordingly, Lebreton et al. (2013) showed that E. faecium strains can be grouped into two distinct clades, of which clade A contains hospital-associated (HA) while clade B contains community-associated strains (CA) strains. A further split in clade A gave rise to the subclades A1 and A2 and subclade A1 now contains mostly animal-associated strains, while subclade A2 contains human clinical isolates (Belloso Daza et al., 2021; Lebreton et al., 2013). The E. faecium sequence type 17 (ST17) was identified to be ancestral for the hospital-associated subclade A1 and forms the clonal complex 17 (CC17). Subsequently, most HA strains were found to be members of CC17 (Lee et al., 2019). Distinguishing characteristics between class A and class B strains are that clade A strains frequently show resistance to antibiotics (vancomycin, ampicillin and high levels of aminoglycosides) and contains virulence factors and mobile genetic elements (Belloso Daza et al., 2021; Gorrie et al., 2019; Lee et al., 2019). Also, clade A strains were found to be ampicillin resistant, while clade B strains are generally ampicillin sensitive (Belloso Daza et al., 2021; Montealegre et al., 2017).
In 2021, Belloso Daza and co-workers investigated the whole genomes of E. lactis, its heterotypic synonym E. xinjiangensis (Li and Gu, 2021; Ren et al., 2016) and E. faecium. Besides the E. faecium type strain NCTC 7171T (= DSM 20477T), they also considered strains from diverse lineages of E. faecium, including HA (clade A) and CA (clade B) strains. In addition, the authors used 16S rRNA gene sequence analysis, phylogenomics based on the core pangenome, multi-locus sequence typing, average nucleotide identity and digital DNA-DNA hybridization to confirm the genetic and evolutionary difference between clade A and the clade B and E. lactis group, the latter which contains isolates at that time identified as E. faecium clade B and E. lactis intertwined in the same cluster. This supported the reassignment of the strains of E. faecium clade B to the species E. lactis. They also confirmed the absence of virulence gene markers IS16, hylEfm and
Some enterococcal strains are utilised as probiotics, like the extensively studied SF68, which is used in pharmaceutical preparations as well as in animal nutrition. These probiotics have been used mainly for the treatment of diarrhoea, antibiotic-associated diarrhoea, irritable bowel syndrome, for lowering cholesterol levels or for immune regulation (Bellomo et al., 1980; Borgia et al., 1982; Bruno and Frigerio, 1981; Buydens and Debeuckelaere, 1996; Chen et al., 2010; D’Souza et al., 2002; Franz et al., 2011; Gade and Thorn, 1989; Greuter et al., 2020; Im et al., 2023; Lewenstein et al., 1979; McFarland and Dublin, 2008; Wunderlich et al., 1989). In the light of recent insights into the population structure of E. faecium strains and the finding that many clade B strains may actually belong to the species E. lactis (Belloso Daza et al., 2021), this study aimed to re-appraise the taxonomy of probiotic E. faecium SF68. On the other hand, the availability of genomic data and further investigation into its competitive nature is aimed to increase knowledge on the functional properties of this strain.
2 Materials and methods
Strains, media and culturing conditions
The probiotic E. faecium SF68 from Cerbios-Pharma SA (Holzapfel et al., 2018) was used for taxonomic and antagonistic activity investigations. All other bacterial strains used in competitive exclusion experiments are shown in Table 1. All enterococci strains were grown in De Man, Rogosa and Sharpe (MRS) agar (VWR International, Darmstadt, Germany) medium aerobically at 30 °C for routine pre-culturing of the strains, while all experiments were conducted at 37 °C.

Characteristics and sources of bacteria used in this study
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
Taxonomic considerations based on chromosome comparison
The unannotated SF68 chromosomal sequence was obtained from Cerbios-Pharma SA under a confidentiality agreement, while other chromosomal sequences of E. faecium and E. lactis, including the type strains DSM 20477T and LMG 25958T, respectively, were downloaded from the NCBI website (Table 2). The software used for the comparison was either the BioNumerics software (Applied Maths, a BioMerieux company, Belgium) version 7.6 to compare strains by phylogenomic tree analysis or the Orthologous Average Nucleotide Identity Tool (OAT) (Lee et al., 2016) to calculate average nucleotide identity between investigated strains, both run on a local Windows computer.
Details of the chromosomal sequences used (besides the SF68 chromosome)1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
In the genome comparison with BioNumerics 7.6, the following parameters were used: minimum of 15 matches in a window size of 25 bases and minimum of 6 matches in a window size of 10 amino acids; minimum stretch length of 30.
Studies on antagonistic activity of SF68 by in silico analysis for bacteriocin genes
The presence of bacteriocin genes was determined from the genome sequence of SF68 using BAGEL4, a web-based bacteriocin mining tool (Van Heel et al., 2018) capable of identifying genes encoding bacteriocins. In addition, the software investigates whether genes encoding proteins involved in bacteriocin biosynthesis, transport, regulation and/or immunity occur in the vicinity of the bacteriocin structural gene.
Competitive exclusion of pathogens
The influence of the probiotic SF68 on the growth of pathogenic enterococci was determined in co-culture. For this, the growth kinetics of individual strains were first determined in vitro in three technical replicates, as to evaluate whether any competitive effects were due to a result of better substrate utilisation. For this, SF68, E. faecalis FAIR-E 329 (potential pathogen based on virulence determinants) and the human pathogenic E. faecalis UKF 210 and E. faecium UKF 207 associated with infections and isolated from catheters (Table 1), were inoculated into 50 ml MRS broth (pH 6.9) with 1% of an overnight culture and then grown for 16 h at 37 °C. The enterococcal counts were determined by plating suitable dilutions in triplicate onto MRS agar and growth parameters such as growth rate and generation time were determined.
For the co-culture experiments, the probiotic, with one pathogen strain in each experiment, were cultured together in 50 ml of bacterial growth medium. MRS broth was used for enterococci, while Standard One (ST1) broth was used when co-culturing SF68 with non-enterococcal pathogens (Table 3).
Strains, inoculum densities and culture media used in co-culture and cell culture experiments
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
Competitive exclusion was studied in three replicates of growth experiments with the co-culture of both probiotic and pathogen strain. The growth of pathogens in coculture was compared to that of the pathogen growing in culture alone. The growth of the bacteria was followed over 7 days at 37 °C and each culture was transferred (1%) on a daily (24 h) basis into fresh 50 ml co-culture medium. Seven days was arbitrarily chosen as a time frame mimicking a probiotic intervention where bacteria had to compete over a time period of a week to see if that would suffice to allow the dominant strain to establish itself. For SF68, 1 × 107 cfu/ml were used for inoculation, while the amount of pathogen inoculated was related to its infective dose as described in the literature (Krämer and Prange, 2023; Table 3). The numbers of bacteria inoculated were confirmed by plate counting. Microbial counts were furthermore determined daily, ca. 8 h after re-inoculation of fresh medium with 1% of the previous batch culture. Selective media were used, which permitted only the growth of either SF68 or the pathogen (Table 3). For the enterococci, MRS agar with antibiotics was used to discriminate between the different strains (Table 3). The minimum inhibitory concentrations (MIC values) for the pathogenic enterococci towards erythromycin were determined using the E-test. All plates and co-cultures were incubated aerobically at 37 °C. The mean of the three replicate colony counts was calculated for each co-culture.
Prevention of pathogen adhesion and invasion in cell culture
Human colorectal adenocarcinoma HT29 cells were grown in DMEM (Dulbecco’s modified Eagle medium) with foetal calf serum (FCS) (1%) and antibiotics (1% penicillin and streptomycin). HT29 cells were incubated at 37 °C in an atmosphere containing 5% CO2. For adhesion and invasion assays, 24 well plates were used, and each well was seeded with 5 × 105 HT29 cells as described previously (Vizoso Pinto et al., 2007). These were grown for 4 days until a confluent monolayer was obtained, and washed twice to remove antibiotics and FCS. For further experimentation, DMEM without FCS and antibiotics was used. Fresh overnight cultures of probiotic and pathogen strains were centrifuged at 7,200
The adhesion data were evaluated after a further hour incubation. For this, the supernatant with non-adherent bacteria was removed. After this, the reversibly bound bacteria were detached from the cells by washing three times with sterile PBS. The eukaryotic cells were then disrupted by using 1.5 ml of Triton-X-100 (1% solution in PBS) and the lysate was diluted and spread-plated onto appropriate medium (Table 3). The number of bacteria counted corresponded to both the adherent and possibly invasive bacteria. The numbers of adherent bacteria were calculated by subtracting the number of invasive bacteria (see below) from this total number. The number of invasive bacteria was determined as follows: first, the non-adherent bacteria were removed by aspirating the supernatant and washing one time with sterile PBS. Subsequently, one ml of DMEM containing the antibiotics gentamicin (500 μg/ml), penicillin (100 μg/ml), ampicillin (100 μg/ml), and streptomycin (1 mg/ml) was added to each well. The DMEM containing antibiotics was left in the wells for 1.5 h to kill all adherent bacteria. After this, 100 μl of supernatant were removed and spread plated to test the killing efficiency. We found that adequate killing led to cell counts of less than 50 cfu/ml. The rest of the supernatant was discarded, and the cells were washed three times with PBS to remove residual antibiotics. The cells were then lysed, similarly to the adhesion assay, and samples were spread plated to determine the numbers of bacteria that invaded the cells. The values obtained for the 4 wells were averaged and the inhibition values were statistically evaluated using ANOVA, followed by a t-test using GraphPad Prism 6, a value of
3 Results and discussion
Taxonomic considerations based on whole genome analyses
Comparison of SF68 genome to genomes of other E. lactis and E. faecium strains
The result of the comparison is shown in Figure 1. The results suggest that both species form separate clusters. It should be noted that by increasing the number of strains, the heterogeneity of the individual clusters also increases and the separation of both species becomes less clear. Figure 1 also illustrates the relatively high heterogeneity of the E. faecium species. However, from this chromosome comparison, there is no doubt that SF68 is a member of the species E. lactis. E. lactis were previously reported to have been isolated from diverse ecological niches, including raw milk cheese, pig and human faeces, dairy products and rice wine koji (Lu et al., 2023).
Result of the chromosome comparison between chromosomes from Enterococcus faecium, Enterococcus lactis strains and SF68. See Table 2 for strain details. Values are expressed in arbitrary units of similarity.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
Comparison of SF68 to the type strains of E. lactis and E. faecium by average nucleotide identity
Using the software OAT (Lee et al., 2016) the average nucleotide identity (ANI) and the Ortho ANI values (Lee et al., 2016) were also calculated. The results presented in Table 4 and Figure 2 clearly show that the type strain of E. faecium can be differentiated from two strains of E. lactis, including the type strain and SF68, by at least 5% ANI, i.e. the ANI levels are lower than 96% between the E. faecium type strain DSM 20477T and the E. lactis strain CX 2-6-2 and strain SF68, respectively. As the species cut-off level for ANI is established to be 96%, this clearly indicates that E. faecium and E. lactis are separate and distinct species and that SF68 belongs to the species E. lactis, rather than E. faecium. On the other hand, the E. lactis type strain LMG 25958T and strain SF68 show 99.58% similarity, indicating these to belong to the same species.
Heatmap generated with OrthoANI values calculated from the OAT software (Lee et al., 2016).
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
Average nucleotide identity (ANI) and ortho ANI values between strains of Enterococcus lactis and the type strain of Enterococcus faecium
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
Bacteriocin gene detection
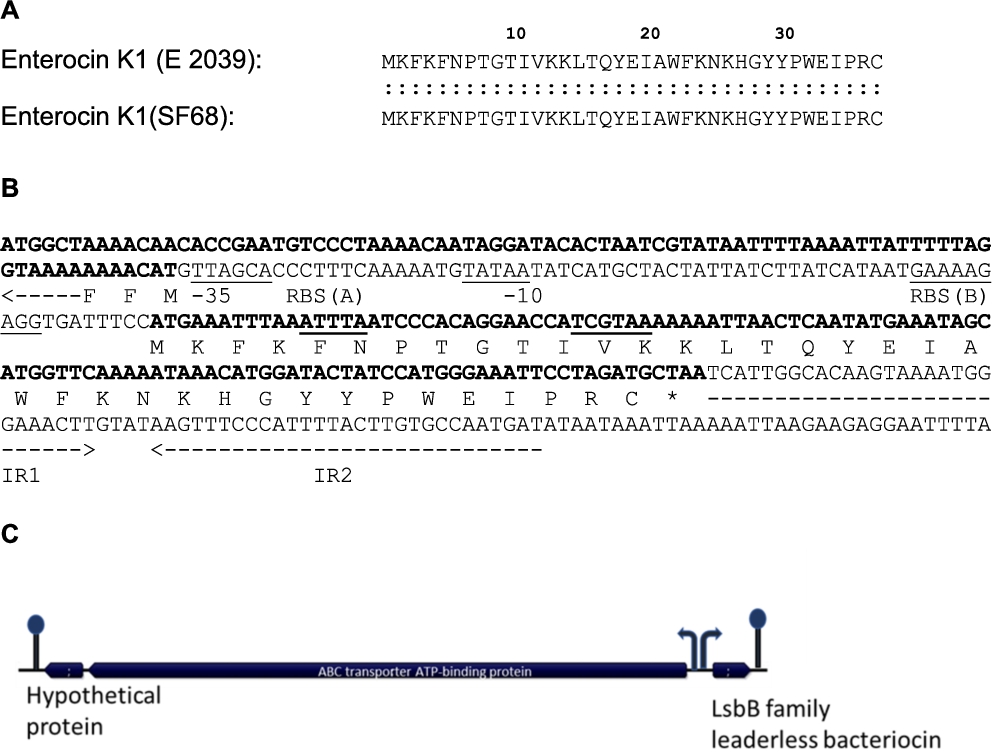
The in silico search for bacteriocin genes on the SF68 genome using the web-based BAGEL identification tool revealed a bacteriocin gene located on the plasmid pF9, for which the predicted protein product was identical to the previously reported 37 amino acid enterocin K1 produced by E. faecium E2039 (Ovchinnikov et al., 2017) (Figure 3A). Together with the enterocins EntQ and EJ97, EntK1 belongs to a relatively new bacteriocin group which is characterised by relatively small peptides (30-50 amino acids) that contain no post-translational modifications, and which are encoded as leaderless peptides lacking a N-terminal leader peptide (Ovchinnikov et al., 2017). Based on Expasy calculations, the encoded EntK1 peptide has cationic (basic) properties with an isoelectric point of 9.73 and a molecular weight of 4,564.39 Da. The target bacterium spectrum for EntK1 was previously reported by Ovchinnikov et al. (2017) and was found to be rather narrow, being restricted mostly to strains of the same genus/species. Importantly, Ovchinnikov et al. (2017) and Reinseth et al. (2021) also found that the bacteriocin was active against nosocomial strains of E. faecium from different European hospitals, including vancomycin-resistant strains, at very low concentrations.
(A) Amino acid sequence comparison of enterocin K1 produced by SF68 and by Enterococcus faecium E 2039 (Ovchinnikov et al., 2017). (B) The genetic locus of the enterocin B bacteriocin gene showing the putative ribosome binding site (RBS), putative promotor (−35 and −10 sites) sequences (underlined) and inverted repeat sequences (IR1 and IR2, inverted arrows) of a putative rho-independent terminator. (C) The annotated genes of the bacteriocin locus show the LsbB family bacteriocin (EntK1) gene, as well genes for an ABC transporter and a hypothetical protein located upstream of the bacteriocin gene in reverse orientation. Putative promoter sequences are shown as arrows, putative terminators as stem-loop structures, indicating that the bacteriocin gene alone, as well as the ABC transporter gene together with the hypothetical protein gene, occur in operons.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
We found the bacteriocin locus to be identical to the open reading frames already present in the GenBank database with accession no MT501398 for the identical bacteriocin structural gene (CDS from position 9,119 to position 9,232) with protein ID QLG04473.1 and the ‘ABC transporter gene’ which is the putative immunity gene (starting at position 9,047 and ending at position 7,344) with a protein ID QLG04472.1. Sequence analysis of the bacteriocin locus in the pF9 nucleotide sequence clearly showed that the bacteriocin gene did not encode a N-terminal leader or a signal peptide (Figure 3B). An AG-rich potential ribosome binding site (5′-GAAAAGAGG-3′) sequence occurred at an optimum 8 bp spacing upstream of the ATG start codon of the bacteriocin structural gene [RBS (B) in Figure 3B]. Putative −35 (5′-GTTAGCA-3′) and −10 (5′-TATAA-3′) promoter sequences are underlined in Figure 3b. The plasmid pF9 sequence from SF68 also harboured an ORF which encoded a predicted immunity gene, similar to that of the Lactococcus lactis bacteriocins LsbA and LsbB (Gajic et al., 2003). This putative immunity gene encodes a 567 amino acid protein and is located immediately upstream of the SF68 EntK1 bacteriocin (Figure 3B,C). The putative immunity protein revealed extensive homology (49.8% identity, 84.6% similarity) to the LmrB immunity protein of Lc. lactis over the entire 567 amino acid length. The ORF encoding the putative immunity protein is located on the opposite DNA strand and the ATG start codon occurs 71 bp upstream of the bacteriocin start codon (Figure 3B). A potential ribosome binding site (5′-GAAAGGG-3′, shown in the reverse orientation, i.e. 3′-CCCTTTC-5′; RBS (A) in Figure 3B) could be located 7 bp upstream of the start codon. The first three deduced amino acids of the immunity protein (MFF) are also shown in Figure 3B. Putative promoter sequences (−35, 3′-TCGTAA-5′; and −10, 3′-ATTTA-5′, both shown in reverse orientation and underlined) are also indicated in Figure 3B. The absence of bacteriocin regulatory genes in the bacteriocin operon could suggest that the bacteriocin is constitutively expressed. The EntK1 bacteriocin was previously determined to be highly active against E. faecium strains, including nosocomial, vancomycin-resistant isolates, while no activity could be shown against E. faecalis strains (Ovchinnikov et al., 2017).
It thus appears that SF68 may potentially be capable of producing a bacteriocin with a reported narrow activity spectrum, but which has been reported to be active against nosocomial vancomycin-resistant E. faecium strains. This bacteriocin activity may therefore potentially play an important role as a functional property in the suppression of potentially pathogenic VRE strains in the human gut. Nevertheless, no inhibition assays were performed to determine that the bacteriocin genes were actually expressed, and bacteriocin production in the MRS medium used was not investigated. This was only assumed to be possibly the case from the fact that the bacteriocin gene appeared to be constitutively expressed (absence of regulatory genes) and the inhibitory activity for this bacteriocin was demonstrated before by Ovchinnikov et al. (2017) and Reinseth et al. (2021), using a similarly nutrient rich bacterial growth medium (brain heart infusion) in their studies.
Competitive exclusion of pathogens in vitro
Co-culturing was used in this study to test whether SF68 would be capable of suppressing the growth of pathogenic, competitor enterococci in vitro. E. faecalis FAIR-E 329 was used as a representative food-borne Enterococcus strain that encodes multiple typical enterococcal virulence factors (Table 1), while the E. faecium strain DSM 13590 was chosen for co-culturing as it represents a typical vancomycin-resistant strain. Pathogenic E. faecium and E. faecalis strains were obtained from the Freiburg University clinic to represent typical human pathogens (Table 1). The growth kinetics of these different Enterococcus strains was pre-assessed in a pilot study to ensure that the strains exhibit similar growth rates or generation times in order not to influence the co-culture experiment. The co-culture experiment revealed that under these conditions SF68 and E. faecium UKF 207 and E. faecalis FAIR-E 329 showed a similar generation time of about 50 min, while the generation time of E. faecalis UKF 210 was somewhat longer at 67 min (results not shown). Despite this longer generation time, this strain reached a maximum viable count of 2.1 × 109 cfu/ml, which was comparable to that of the other strains (result not shown).
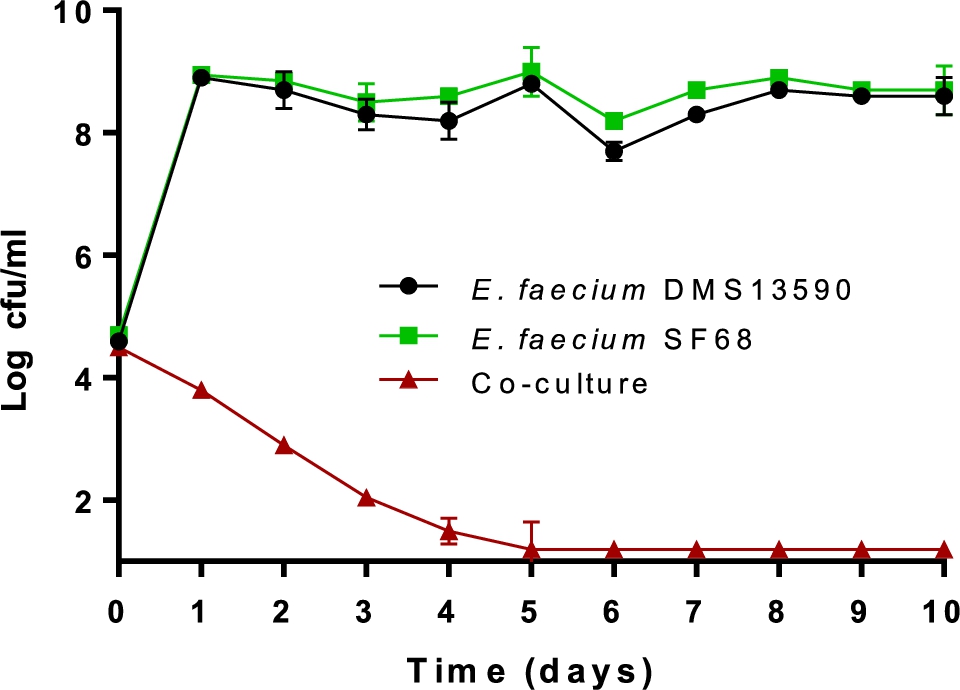
The inhibitory effect of SF68 on the growth of pathogenic bacteria was clearly strain specific. Nevertheless, it generally appeared to affect Gram-positive more than Gram-negative bacteria, which may be indicative of bacteriocin activity, as Gram-negative bacteria are generally more sensitive to organic acids and low pH. The result of an initial co-culture pilot experiment of the vancomycin-resistant E. faecium DSM 13590T (type strain from human faeces) with the probiotic SF68 is shown in Figure 4. Co-culturing of SF68 with the vancomycin-resistant E. faecium DSM 13590T showed that the probiotic strain grew from ca. log 4.5 cfu/ml to ca. log 9 cfu/ml within one day and stayed at this high level during continued subculturing and incubation. In contrast, the vancomycin-resistant E. faecium DSM 13590T strain steadily declined to below log 2 cfu/ml after day 5, staying at this level up to 10 days of subculturing (Figure 4).
Co-culture of probiotic SF68 together with vancomycin-resistant Enterococcus faecium 13590 during a pilot experiment performed in MRS broth medium at 37 °C for 10 days. The numbers (log10 cfu/ml) of probiotic E. faecium SF68 (RIFREC) were determined using MRS agar containing rifampicin (64 μg/ml), of vancomycin resistant E. faecium DSM 13590 (VREC) on agar containing vancomycin (64 μg/ml) and of total enterococci (TEC) were determined on MRS agar without antibiotic. The detection limit reached was log 1.3 cfu/ml.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
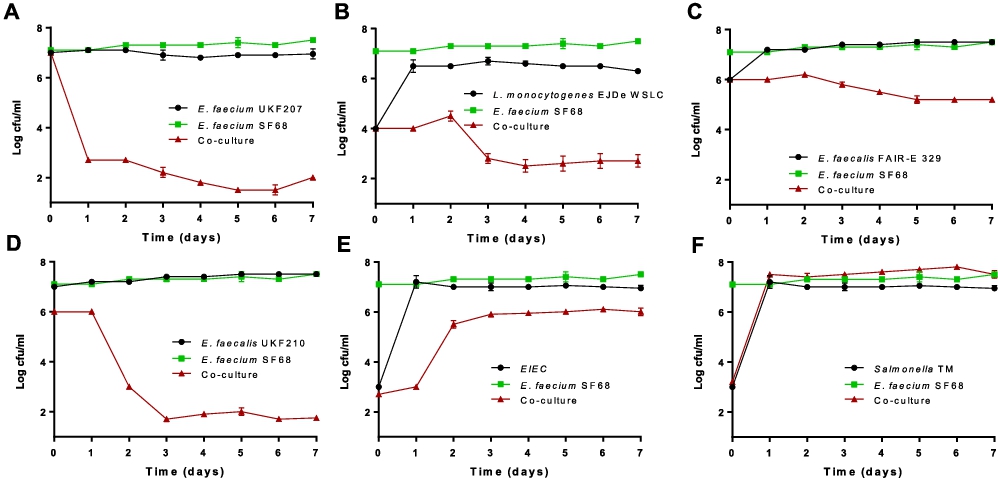
Following this pilot experiment, the effect of the probiotic SF68 was also tested on enterococcal pathogens isolated from infections, as well as other pathogens. The growth of the pathogenic E. faecium 207 and L. monocytogenes EJDe was also inhibited by SF68 (almost 5 log units decrease in viable numbers for the enterococci and 1.5 log decrease for the L. monocytogenes strains, respectively) (Figure 5A,B). The growth of E. faecalis FAIR-E 329 was also reduced by approx. 1 log unit (Figure 5C). One exception was E. faecalis UKF 210 that inhibited SF68 by 4 logs (Figure 5D). However, it should be noted that at the starting point SF68 was ca. 1 log less concentrated at ca. 1 × 106 cfu/ml, which may have been an advantage for strain UKF 210 and which could have caused it to outgrow the probiotic SF68. Furthermore, the strain E. faecalis UKF 210 was also found to produce cytolysin in a PCR test for the cytolysin gene (results not shown). This compound has cytolytic activity against prokaryotic as well as eukaryotic cells (Coburn and Gilmore, 2003) and may explain why the E. faecalis UKF 210 strain was able to inhibit the probiotic SF68 in co-culture. Thus, while SF68 clearly exhibited a strong inhibitory effect against pathogenic and vancomycin-resistant E. faecium strains, as well as against pathogenic L. monocytogenes, competitive exclusion of E. faecalis strains clearly showed different outcomes.
Numbers of probiotic Enterococcus faecium SF68 (RifR) in co-culture with (A) E. faecium strains UKF207 (pathogenic, EryR), (B) Listeria monocytogenes EJDe WSLC, (C) FAIR-E 329 (semi-hard cheese, EryR), (D) Enterococcus faecalis strain UKF210 (pathogenic, EryR), (E) enteroinvasive Escherichia coli EIEC 12860 and (F) Salmonella enterica serovar Typhimurium S.TM S1 1344 alone or during co-culturing experiment for 7 days. The detection limit was log 1.3 cfu/ml. Note that in these graphs the red ‘coculture’ curve indicates the pathogen counts except for (D) where the co-culture curve indicates the SF68 counts.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
The effect of SF68 on Gram-negative pathogens is also shown in Figure 5. The growth inhibition effect of pathogenic Gram-negative bacteria was poor, if present at all. Only the entero-invasive E. coli 12860 was slightly, but significantly (
Competitive exclusion has already been suggested to be based on several modes of action that eliminate pathogens, i.e. (1) direct and indirect competition for nutrients, (2) competition for physical attachment sites, (3) production of antimicrobial compounds (e.g. bacteriocins or volatile fatty acids including lactic acid), (4) enhancement of host immune system activity and (5) a synergistic interaction of two or more of the above activities (Callaway et al., 2008). In food animals, competitive exclusion was already shown to be effective to control Salmonella colonization in poultry or enterotoxigenic E. coli in swine (Nisbet et al., 1993; Underdahl et al., 1982, Ushe and Nagy 1985). The probiotic activity of SF68 in prevention of diarrhea (Bellomo et al., 1980; Buydens and Debeuckelaere, 1996; Wunderlich et al., 1989) may be hypothetically at least partially explained by the competitive exclusion properties observed here for the strains in vitro, and potentially also by the production of antimicrobial metabolites such as lactic acid and possibly bacteriocin.
Inhibition of pathogen adherence and invasion in HT29 cell culture
Viable counts (log cfu/ml) of (A) adhering or invading Enterococcus faecium SF68, (B) adhering or invading Gram-positive pathogens Enterococcus faecalis FAIR-E 329, (C) E. faecalis UKF 210, (D) E. faecium UKF 207, and (E) Listeria monocytogenes EJDe. Shown is the adhesion of pathogens to cells not treated (adhesion) and pre-treated with the probiotic SF68 (inhibition of adhesion), and the invasion of pathogens into HT 29 cells in non-treated (invasion) and cells pre-treated with the probiotic SF68 (inhibition of invasion). All values were determined in quadruplicate and the standard error is shown. A significant inhibition of either adhesion or invasion is indicated with an asterisk (*).
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
The adherence and invasion of SF68 were tested in a pilot study. SF68 adhered well to HT29 cell monolayers, but it was non-invasive (Figure 6A) and could therefore be used as a negative control strain for invasion assays. The results for the competitive exclusion of Gram-positive pathogen adhesion and invasion with the probiotic SF68 are also shown in Figure 6. The inhibition of adhesion and invasion of these pathogens by SF68 was strain specific. Similar to SF68, the Enterococcus strain FAIR-E 329 from semi-hard cheese was not invasive under the conditions of our study (result not shown). The adhesion of strain FAIR-E 329 was significantly (
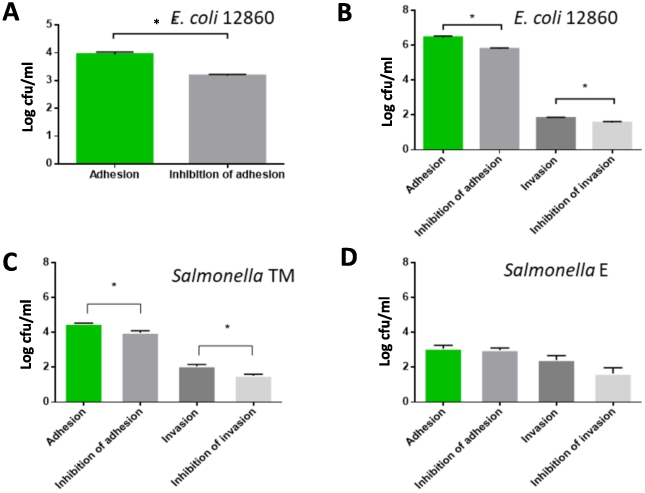
Similar to the competitive exclusion in growth medium results, the Gram-negative pathogens were generally less inhibited in their adhesion and invasion potential by the probiotic. The E. coli strain 12860 was not invasive when used at a concentration of 2 × 106 cfu/ml under the conditions of this study. However, the adhesion of this strain could be significantly (
Viable counts (log cfu/mL) of (A) adhering or invading Gram-negative Escherichia coli 12860 with an inoculum density of 2 × 106 cfu/ml, (B) E. coli 12860 with an inoculum density of 2 × 109 cfu/ml, (C) Salmonella TM, and (D) Salmonella E. Shown is the adhesion of pathogens to cells not treated (first column, green) and pre-treated with the probiotic SF68 (second column, grey), and the invasion of pathogens into HT 29 cells in non-treated (third column, dark grey) and cells pre-treated with the probiotic SF68 (fourth column, light grey). All values were determined in quadruplicate and the standard error is shown. A significant (
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00005
The probiotic SF68 showed good adherence to HT29 colon carcinoma cells, while it was clearly non-invasive (Figure 6A). These properties, i.e. to adhere to, but not invade intestinal cells have previously been described as important traits for probiotic strains. In most cases, the probiotic SF68 also showed a significant inhibition of adherence and invasion of bacterial pathogens. The degree of inhibition of pathogens in terms of bacterial counts showed differences at strain level.
4 Conclusions
The results obtained in this study clearly show that the probiotic E. faecium SF68 can be unequivocally reclassified as E. lactis. Reclassification of bacteria is not uncommon with the advance of modern taxonomic tools based on whole genome comparisons. This was also the case for another lactic acid bacterial genus, i.e. Lactobacillus, which was recently re-classified to give rise to even 23 new genera, some of which contain probiotic species (Zeng et al., 2020). The data generated in this study shows that SF68 may have the capacity to produce a bacteriocin enterocin K1, that was previously described in another strain for its activity especially against pathogenic, vancomycin-resistant E. faecium strains. Nevertheless, as bacteriocin expression and activity were not determined in this study, we can only imply bacteriocin activity as a potential functional benefit of the strain, but not a confirmed property. Furthermore, our data show that SF68 adheres to intestinal cells and successfully inhibits the adhesion and prevents invasion of several pathogenic bacterial strains in cell culture. These data may serve as basis to explain why SF68 is such a successful probiotic with proven effectiveness and long history of safe use (Holzapfel et al., 2018). Previous reports also indicated that E. lactis strains do not contain virulence factors, such as haemolysin and gelatinase (Ben Braiek et al., 2018). This may bring us one step closer to answering the previously asked conundrum (Ferchichi et al., 2021; Franz et al., 2003) on the safety of E. faecium strains. These have previously been noted by many authors to be of dualistic nature, i.e. some are clearly pathogenic, whilst some are beneficial as probiotics (Franz et al., 2011), or are even important from a biotechnological point of view in, e.g. cheese ripening (Giraffa, 2003; Litopoulou-Tzanetaki et al., 1993; Sarantinopoulos et al., 2002). The answer may lie in the fact that certain strains may have been misclassified as in the case of E. lactis, a species which has low pathogenic but high biotechnological potential (Ben Braiek et al., 2018). Further investigations should focus on this issue in more depth.
Corresponding author; e-mail: charles.franz@mri.bund.de
Acknowledgements
The authors would like to thank M. Aeschbacher for his great input in the discussion of this research
Authors’ contribution
Conceptualization: CF, BP, AA, RC, WH; methodology: CF, BP, MV-P, WH; investigations: CF, BP, MV-P, data analysis: CF, BP, MV-P, WH; writing – original draft preparation: CF, BP, MV-P; writing – review and editing: CF, BP, MV-P, AA, RC, WH. All authors have read and agreed to the published version of the manuscript.
Conflict of interest
BP is also employed by Yakult Europe BV., WH is also associated with HEM Pharma Inc., RC and AA are employed by Cerbios-Pharma S.A.
References
Bellomo, G., Mangiagle, A., Nicastro, L. and Frigerio, G., 1980. A controlled double-blind study of SF68 strain as a new bio logical preparation for the treatment of diarrhoea in pediatrics. Current Therapeutic Research 28: 927-934.
Belloso Daza, M.V., Cortimiglia, C., Bassi, D. and Cocconcelli, P.S., 2021. Genome-based studies indicate that the Enterococcus faecium Clade B strains belong to Enterococcus lactis species and lack of the hospital infection associated markers. International Journal of Systematic and Evolutionary Microbiology 71: 004948.
Ben Braïeka, O., Morandic, S., Cremonesid, P., Smaouie, S., Hanif, K. and Ghrairia, T., 2018. Biotechnological potential, probiotic and safety properties of newly isolated enterocin-producing Enterococcus lactis strains. LWT – Food Science and Technology 92: 361-370.
Borgia, M., Sepe, N., Brancato, V. and Borgia, R., 1982. A controlled clinical study on Streptococcus faecium preparation for the prevention of side reactions during long-term antibiotic treatments. Current Therapeutic Research 31: 265-271.
Bruno, F. and Frigerio, G., 1981. Eine neuartige Möglichkeit zur Behandlung der Enteritis – Kontrollierte Doppelblindversuche mit dem Stamm SF68. Schweizerische Rundschau für Medizin Praxis 39: 1717-1720.
Buydens, P. and Debeuckelaere, S., 1996. Efficacy of SF 68 in the treatment of acute diarrhea – a placebo-controlled trial. Scandinavian Journal of Gastroenterology 31: 887-891.
Callaway, T.R., Edrington, T.S., Anderson, R.C., Harvey, R.B., Genovese, K.J., Kennedy, C.N., Venn, D.W. and Nisbet, D.J., 2008. Probiotics, prebiotics and competitive exclusion for prophylaxis against bacterial disease. Animal Health Research Reviews 9: 217-225.
Chen, C.-C., Kong, M.-S., Lai, M.-W., Chao, H.-S., Chang, K.-W., Chen, S.A.-Y., Huang, Y.-C., Chiu, C.-H., Li, W.-C., Lin, P.-Y., Chen, C.-J. and Lin, T.-Y., 2010. Probiotics have clinical, microbiologic, and immunologic efficacy in acute infectious diarrhea. Pediatric Infectious Disease Journal 29: 135-138.
Chenoweth, C. and Schaberg, D., 1990. The epidemiology of enterococci. European Journal of Clinical Microbiology and Infectious Diseases 9: 80-89.
Coburn, P.S. and Gilmore, M.S., 2003. The Enterococcus faecalis cytolysin: a novel toxin active against eukaryotic and prokaryotic cells. Cellular Microbiology 5: 661-669.
D’ Souza, A.L., Rajkumar, C., Cooke, J. and Bulpitt, C.J., 2002. Probiotics in prevention of antibiotic associated diarrhoea: meta-analysis. British Medical Journal 324: 1361.
Devriese, L.A., Hommez, J., Pot, B. and Haesebrouck, F., 1994. Identification and composition of the streptococcal and enterococcal flora of tonsils, intestines and faeces of pigs. Journal of Applied Bacteriology 77: 31-36.
Devriese, L.A. and Pot, B., 1995. The genus Enterococcus. In: Wood, B.J.B. and Holzapfel, W.H. (eds.) The genera of lactic acid bacteria. The lactic acid bacteria vol. 2. Springer, Berlin, Germany, pp. 327-367.
Ferchichi, M., Sebei, S., Boukerb, A.M., Karray-Bouraoui, N., Chevalier, S., Feuilloley, M.G.J., Connil, N. and Zommiti, M., 2021. Enterococcus spp.: is it a bad choice for a good use-a conundrum to solve? Microorganisms 9: 2222.
Franz, C.M.A.P., Holzapfel, W.H. and Stiles, M.E., 1999. Enterococci at the crossroads of food safety? International Journal of Food Microbiology 47: 1-24.
Franz, C.M.A.P., Stiles, M.E., Schleifer, K.-H. and Holzapfel, W.H., 2003. Enterococci in foods – a conundrum for food safety. International Journal of Food Microbiology 88: 105-122.
Franz, C.M.A.P., Huch, M., Abriouel, H., Holzapfel, W.H. and Gálvez, A., 2011. Enterococci as probiotics and their implications in food safety. International Journal of Food Microbiology 151: 125-140.
Gade, J. and Thorn, F., 1989. Paraghurt for patients with irritable bowel syndrome: a controlled clinical investigation from general practice. Scandinavian Journal of Primary Health Care 7: 23-26.
Gajic, O., Buist, G., Kojic, M., Topisirovic, L., Kuipers, O.P. and Kok, J., 2003. Novel mechanism of bacteriocin secretion and immunity carried out by lactococcal multidrug resistance proteins. Journal of Biological Chemistry 278: 34291-34298.
Giraffa, G., 2003. Functionality of enterococci in dairy products. International Journal of Food Microbiology 88: 215-222.
Gorrie, C., Higgs, C., Carter, G., Stinear, T.P. and Howden, B., 2019. Genomics of vancomycin-resistant Enterococcus faecium. Microbial Genomics 5: e000283.
Greuter, T., Michel, M.C., Thomann, D., Weigmann, H. and Vavricka, S.R., 2020. Randomized, placebo-controlled, double-blind and open-label studies in the treatment and prevention of acute diarrhea with Enterococcus faecium SF68. Frontiers in Medicine 7: 276.
Hanchi, H., Mottawea, W., Sebei, K. and Hammami, R., 2018. The genus Enterococcus: between probiotic potential and safety concerns-an update. Frontiers in Microbiology 9: 1791.
Holzapfel, W.H., Arini, A., Aeschbacher, M., Coppolecchia, R. and Pot, B., 2018. Enterococcus faecium SF68 as a model for efficacy and safety evaluation of pharmaceutical probiotics. Beneficial Microbes 9: 375-388.
Im, E.J., Lee, H.H., Kim, M. and Kim, M.K., 2023. Evaluation of enterococcal probiotic usage and review of potential health benefits, safety, and risk of antibiotic-resistant strain emergence. Antibiotics 12: 1327.
Krämer, J. and Prange, A., 2023. Lebensmittel-Mikrobiologie. 8th ed. Verlag Eugen Ulmer, Stuttgart, Germany, pp. 25-90.
Lebreton, F., van Schaik, W., Manson McGuire, A., Godfrey, P., Griggs, A., Mazumdar, V., Corander, J., Cheng, L., Saif, S., Young, S., Zeng, Q., Wortman, J., Birren, B., Willems, R.J.L., Earl, A.M. and Gilmore, M.S., 2013. Emergence of epidemic multidrug-resistant Enterococcus faecium from animal and commensal strains. mBio 4: e00534-13.
Leclerc, H., Devriese, L.A. and Mossel, D.A.A., 1996. Taxonomical changes in intestinal (faecal) enterococci and streptococci: consequences on their use as indicators of faecal contamination in drinking water. Journal of Applied Bacteriology 81: 459-466.
Lee, I., Kim, Y.O., Park, S.-C. and Chun, J., 2016. OrthoANI: an improved algorithm and software for calculating average nucleotide identity. International Journal of Systematic and Evolutionary Microbiology 66: 1100-1103.
Lee, T., Pang, S., Abraham, S. and Coombs, G.W., 2019. Antimicrobial-resistant CC17 Enterococcus faecium: the past, the present and the future. Journal of Global Antimicrobial Resistance 16: 36-47.
Li, Y.Q. and Gu, C.Z., 2021. Proposal of Enterococcus xinjiangensis Ren et al. 2020 as a later heterotypic synonym of Enterococcus lactis Morandi et al. 2012. International Journal of Systematic and Evolutionary Microbiology 71: 004716.
Litopoulou-Tzanetaki, E., Tzanetakis, N. and Vafopoulou-Mastrojiannaki, A., 1993. Effect of type of lactic starter on microbiological, chemical and sensory characteristics of Feta cheese. Food Microbiology 10: 31-41.
Lu, J., Shen, T., Zhang, Y., Ma, X., Xu, S., Awad, S., Du, M. and Zhong, Z., 2023. Safety assessment of Enterococcus lactis based on comparative genomics and phenotypic analysis. Frontiers in Microbiology 14: 1196558.
McFarland, L.V. and Dublin, S., 2008. Meta-analysis of probiotics for the treatment of irritable bowel syndrome. World Journal of Gastroenterology 14: 2650-2661.
Montealegre, M.C., Roh, J.H., Rae, M., Davlieva, M.G., Singh, K.V., Shamoo, Y. and Murray, B.E., 2017. Differential penicillin-binding protein 5 (PBP5) levels in the Enterococcus faecium clades with different levels of ampicillin resistance. Antimicrobial Agents in Chemotherapy 61: e02034-16.
Morandi, S., Cremonesi, P., Povolo, M. and Brasca, M., 2012. Enterococcus lactis sp. nov., from Italian raw milk cheeses. International Journal of Systematic and Evolutionary Microbiology 62: 1992-1996.
Morrison, D., Woodford, N. and Cookson, B., 1997. Enterococci as emerging pathogens of humans. Journal of Applied Microbiology Symposium Supplement 83: 89S-99S.
Murray, B.E., 1990. The life and times of the Enterococcus. Clinical Microbiology Reviews 3: 46-65.
Nisbet, D.J., Corrier, D.E., Scanlan, C.M., Hollister, A.G., Beier, R.C. and DeLoach, J.R., 1993. Effect of a defined continuous-flow derived bacterial culture and dietary lactose on Salmonella typhimurium colonization in broiler chickens. Avian Diseases 37: 1017-1025.
Ovchinnikov, K.V., Kristiansen, P.E., Straume, D., Jensen, M.S., Aleksandrzak-Piekarczyk, T., Nes, I.F. and Diep, D.B., 2017. The leaderless bacteriocin enterocin K1 is highly potent against Enterococcus faecium: a study on structure, target spectrum and receptor. Frontiers in Microbiology 8: 774.
Reinseth, I., Tønnesen, H.H., Carlsen, H. and Diep, D.B., 2021. Exploring the herapeutic potential of the leaderless enterocins K1 andEJ97 in the treatment of vancomycin-resistant enterococcal infection. Frontiers in Microbiology 12: 649339.
Ren, X., Li, M. and Guo, D., 2016. Enterococcus xinjiangensis sp. nov., isolated from yogurt of Xinjiang, China. Current Microbiology 73: 374-378.
Sarantinopoulos, P., Kalantzopoulos, G. and Tsakalidou, E., 2002. Effect of Enterococcus faecium on microbiological, physicochemical and sensory characteristics of Greek Feta cheese. International Journal of Food Microbiology 76: 93-105.
Selleck, E.M., Van Tyne, D. and Gilmore, M.S., 2019. Pathogenicity of enterococci. Microbiology Spectrum 7: 10.1128.
Svec, P. and Franz, C.M.A.P., 2014. The genus Enterococcus. Chapter 15. In: Holzapfel, W.H. and Wood, B.J.B. (eds.) Lactic acid bacteria – biodiversity and taxonomy. John Wiley and Sons Ltd., Chichester, UK, pp. 391-404.
Underdahl, R., Torres-Medina, A. and Doster, A.R., 1982. Effect of Streptococcus faecium C-68 in control of Escherichia coli induced diarrhea in gnotobiotic pigs. American Journal of Veterinary Research 43: 2227-2232.
Ushe, T.C. and Nagy, B., 1985. Inhibition of small intestinal colonization of enterotoxigenic Escherichia coli by Streptococcus faecium M74 in pigs. Zentralblatt für Bakteriologie und Hygiene. I. Abteilung Originale B 181: 374-382.
Van Heel, A.J., De Jong, A., Song, C., Viel, J.H., Kok, J. and Kuipers, O.P., 2018. BAGEL4: a user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Research 46(W1): W278-W281.
Vancanneyt, M., Lombardi, A., Andrighetto, C., Knijff, E., Torriani, S.K., Björkroth, J., Franz, C.M.A.P., Foulquié Moreno, M.R., Revets, H., De Vuyst, L., Swings, J., Kersters, K., Dellaglio, F. and Holzapfel, W.H., 2002. Intraspecies genomic groups in Enterococcus faecium and their correlation with origin and pathogenicity. Applied and Environmental Microbiology 68: 1381-1391.
Wessels, D., Jooste, P.J. and Mostert, J.F., 1990. Technologically important characteristics of Enterococcus isolates from milk and dairy products. International Journal of Food Microbiology 10: 349-352.
Wunderlich, P.F., Braun, L., Fumagalli, I., D’Apuzzo, V., Heim, F., Karly, M., Lodi, R., Politta, G., Vonbank, F. and Zeltner, L., 1989. Double-blind report on the efficacy of lactic acid-producing Enterococcus SF68 in the prevention of antibiotic-associated diarrhoea and in the treatment of acute diarrhoea. Journal of International Medical Research 17: 333-338.
Zheng, J., Wittouck, S., Salvetti, E., Franz, C.M.A.P., Harris, H.M.B., Mattarelli, P., O’Toole, P.W., Pot, B., Vandamme, P., Walter, J., Watanabe, K., Wuyts, S., Felis, G.E., Gänzle, M.G. and Lebeer, S., 2020. A taxonomic note on the genus Lactobacillus: description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. International Journal of Systematic and Evolutionary Microbiology 70: 2782-2858.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}