Abstract
Previous studies have demonstrated that soymilk and Lacticaseibacillus paracasei YIT 9029 (strain Shirota: LcS) each beneficially affect the gut microbiota and defecation habits. To investigate the effects of daily consumption of fermented soymilk containing LcS (FSM), we conducted a randomised, double-blind, placebo-controlled study of 112 healthy Japanese adults with a low faecal Bifidobacterium count. They consumed 100 ml FSM or placebo (unfermented soymilk base) once daily for 4 weeks. Their gut microbiota was analysed by 16S rRNA gene amplicon sequencing and quantitative reverse transcription-polymerase chain reaction (PCR), and faecal short-chain fatty acids (SCFAs) and urinary putrefactive products were assessed during the pre- and post-consumption periods. Defecation habits were examined weekly using a subjective questionnaire. In the post-consumption period, living LcS were not detected in two subjects in the FSM group (n = 57) but were detected in one subject in the SM group (n = 55). The FSM group had a significantly higher number and relative abundance of faecal lactobacilli compared with the placebo group. The relative abundance of Bifidobacterium, alpha-diversity of microbiota, and concentrations of acetate and total SCFAs in faeces were significantly increased in the FSM group, although no significant differences were detected between the groups. The number of defecations and defecation days per week significantly increased in both groups. Subgroup analysis of 109 subjects, excluding 3 with inconsistent LcS detection (2 and 1 subjects in the FSM and SM groups, respectively), revealed that the FSM group (n = 55) had significantly greater increases in faecal acetate concentration compared with the SM group (n = 54) and significant upregulation of pathways related to energy production or glucose metabolism in the gut microbiota. These findings suggest that daily FSM consumption improves the gut microbiota and intestinal environment in healthy adults and may help to maintain health and prevent diseases.
Registered at the University Hospital Medical Information Network (UMIN) clinical trials registry under: UMIN 000035612.
1 Introduction
The gut microbiota is a complex ecosystem consisting of over 100 trillion microorganisms belonging to more than 1000 species (Rajilić-Stojanović et al., 2007) that have an impact on the health of the host via their metabolites, which may be beneficial or harmful (Nicholson et al., 2012). Short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, are produced mainly by the fermentation of carbohydrates in the colon and play important roles in maintaining intestinal homeostasis, not only functioning as an energy source for colon cells (Den Besten et al., 2013) but also suppressing colonisation by pathogenic bacteria (Havenaar, 2011), preventing infectious diseases (Fukuda et al., 2011), strengthening the intestinal mucosal barrier (Hamer et al., 2010), modulating immune function (Furusawa et al., 2013), and enhancing anti-inflammatory activity (Verbeke et al., 2015). Recently, many studies have reported close associations between the gut microbiota and various disorders, including obesity (Turnbaugh et al., 2006), diabetes (Wang et al., 2012a), inflammatory bowel disease (Hiutung et al., 2016), colorectal cancer (Wang et al., 2012b), and mental illness (Aizawa et al., 2019); patients with these disorders have lower levels of SCFA producers in their gut microbiota (Markowiak-Kopeć and Śliżewska, 2020). In contrast, as another type of metabolite of the gut microbiota, putrefactive products such as phenol, p-cresol, indole, and ammonia generated by the fermentation of undigested protein or the decomposition of amino acids in the colon are harmful to the host (Yao et al., 2016). These putrefactive products are reported to inhibit the maintenance of intestinal homeostasis by disrupting the intestinal mucosal barrier and immunosuppression (Carbonero et al., 2012; Shiba et al., 2014); moreover, they promote the onset of colorectal cancer and chronic kidney disease (Evenepoel et al., 2009; Toden et al., 2005). Hence, balancing the gut microbiota and promoting a good intestinal environment through an increase of SCFAs and a decrease of putrefactive products are considered to be important to maintain health and prevent diseases.
Probiotics are defined as ‘live microorganisms that, when administered in adequate amounts, confer a health benefit on the host’ (Hill et al., 2014). Lacticaseibacillus paracasei YIT 9029 (strain Shirota, LcS, formerly Lactobacillus casei strain Shirota) is a representative probiotic that protects against upper respiratory tract infection (Shida et al., 2017), induces immunomodulation (Nagao et al., 2000), reduces the risk of some cancers (Ishikawa et al., 2005; Naito et al., 2008; Toi et al., 2013), relieves stress (Kato-Kataoka et al., 2016), and improves sleep quality (Takada et al., 2017). Furthermore, several clinical studies have demonstrated that the daily consumption of fermented milk containing LcS provides a good balance of gut microbiota and improves the intestinal environment (Nagata et al., 2016; Wang et al., 2015) and defecation habits, including ameliorating constipation and loose stools (Matsumoto et al., 2006, 2010). In addition, a recent epidemiological study of healthy Japanese adults observed that the frequency of consuming fermented milk containing LcS was positively correlated with stool frequency and faecal bifidobacteria density (Shima et al., 2019).
Soymilk (SM) contains not only soy proteins and peptides but also various functional ingredients, such as saponins, oligosaccharides, and isoflavones, and is consumed worldwide as a functional beverage. Soy isoflavones, which are typical functional ingredients in SM, and their metabolites act as antagonists and agonists of oestrogen receptors to modulate the activity of 17β-oestradiol, a potent oestrogen, because of their similar structures (Mueller et al., 2004). Soy isoflavones and their metabolites exert beneficial effects regarding the prevention of breast and prostate cancers and the improvement of skin conditions, menopausal symptoms, and osteoporosis (Gómez-Zorita et al., 2020). Moreover, raffinose and stachyose, which are oligosaccharides in SM, reach the gut without undergoing digestion and function as prebiotics in humans (Gibson et al., 2004). Soy oligosaccharides are utilised by various species of Bifidobacterium, thus stimulating their proliferation in vitro (Inoguchi et al., 2012). Moreover, several clinical studies have demonstrated that the consumption of soy oligosaccharides or SM increases the viable count of intrinsic Bifidobacterium in the gut of healthy adults and improves defecation habits in healthy subjects with a tendency for constipation (Fujisawa et al., 2017; Hayakawa et al., 1990).
Because fermented SM with LcS (FSM) contains probiotic LcS and prebiotic soy oligosaccharides, it is expected to have stronger effects as a symbiotic compared with LcS or SM alone, thereby balancing the gut microbiota and improving the intestinal environment more effectively. Our pilot study demonstrated that the consumption of FSM twice daily for 8 weeks improves the gut microbiota in healthy pre-menopausal women, such as decreasing Enterobacteriaceae significantly and increasing Bifidobacterium non-significantly compared with the pre-consumption period in the FSM group (Nagino et al., 2018). However, the effect on the intestinal environment was not evaluated in that study and the evidence was not sufficient. Therefore, to examine the effects of the daily consumption of FSM on the gut microbiota and intestinal environment, we conducted a randomised, double-blind, placebo-controlled study on healthy Japanese adults with a low gut population of intrinsic Bifidobacterium. Moreover, we examined the effects of FSM consumption on defecation habits.
2 Materials and methods
Test beverages
FSM and unfermented placebo SM were used as the test beverages, as described previously (Nagino et al., 2015). FSM was produced via SM (Fuji Oil Co., Ltd., Osaka, Japan) fermentation using LcS obtained from the Yakult Central Institute (Tokyo, Japan), and SM was unfermented SM with the same nutritional content, colour, flavour, taste, and pH as FSM. Lactic acid was added to the placebo to achieve the same pH as the FSM. FSM contained more than 3.0 × 1010 cfu/100 ml LcS, 0.09 g/100 ml raffinose, and 0.37 g/100 ml stachyose, whereas SM contained 0.09 g/100 ml and 0.39 g/100 ml, respectively. Beverage composition is detailed in Supplementary Table S1.
Subjects
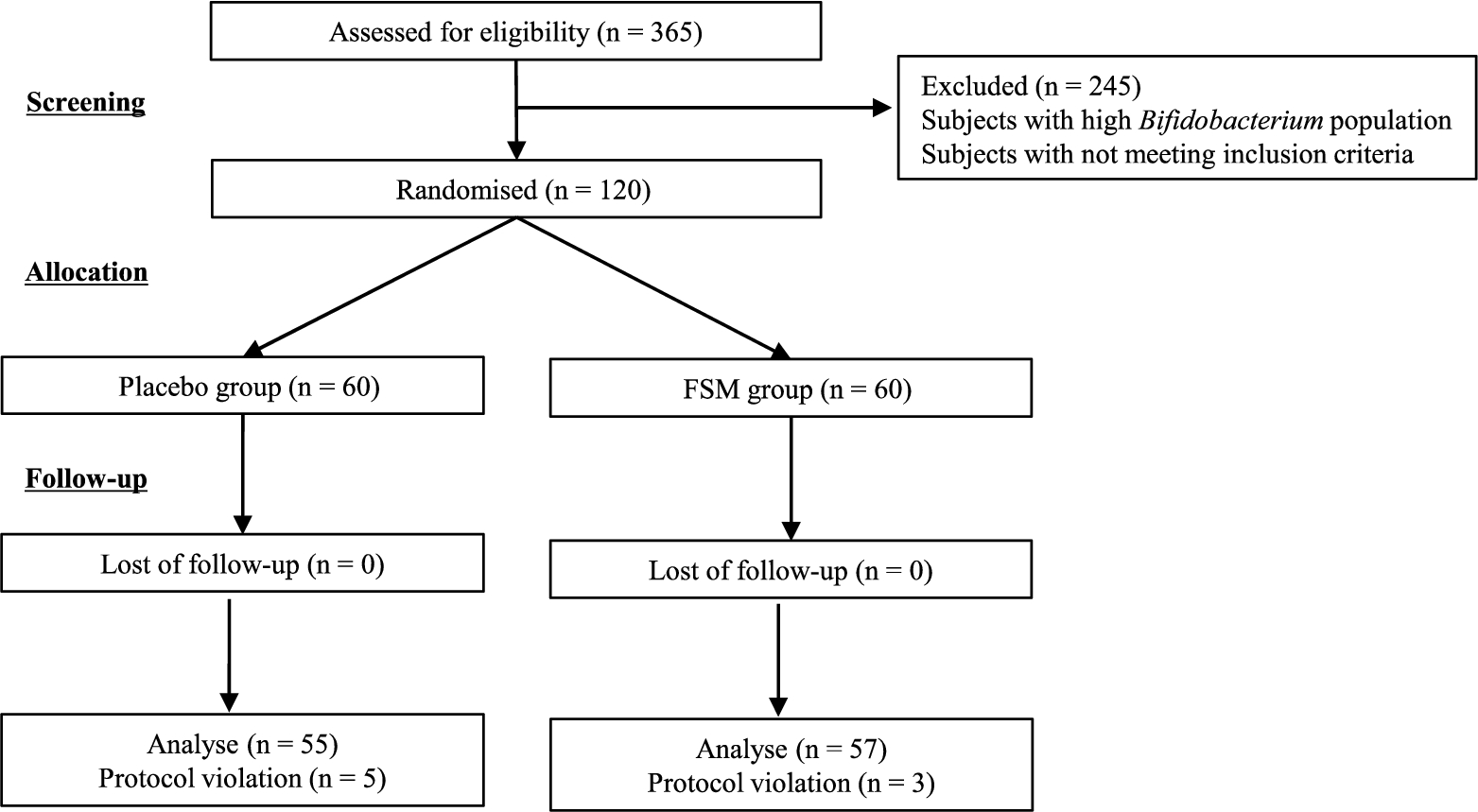
This study was conducted according to the guidelines of the Declaration of Helsinki, and all procedures involving human subjects were approved by the ethics committee of the Chiyoda Paramedical Care Clinic, Tokyo, Japan (registration number: YKH18C1). Written informed consent was obtained from all subjects prior to enrolment. To screen 120 healthy subjects in the study, 365 healthy Japanese adults were initially enrolled. The inclusion criteria were age greater than 20 and less than 60 years, no medications, no diseases, and the maintenance of a habitual lifestyle during the study. The exclusion criteria were a history of severe disorders, habitual consumption of laxative products, food allergy, drug allergy, SM allergy, milk allergy, inability to consume SM, pregnancy or lactation, planning to become pregnant, enrolment in another clinical study, and unsuitability for the study as judged by the director. During the study, the subjects complied with dietary restrictions to avoid the intake of other fermented milk, yogurt, lactic acid bacteria-containing beverages, and pro- and prebiotic products other than the test beverage.
Study design
A double-blind, placebo-controlled, and parallel-group study was conducted at the Chiyoda Paramedical Care Clinic and registered at the University Hospital Medical Information Network (UMIN) clinical trials registry under the following registration number: UMIN 000035612. After screening the subjects (shown in Figure 1 and in the ‘Subjects’ subsection of the Results), 120 healthy subjects were allocated randomly to the FSM or placebo group, and they consumed a dose of 100 ml FSM or placebo once a day for 4 weeks (from March to April 2019). The subjects were asked to self-record their consumption of the beverages and defecation habits every day in a diary. Faeces and first-morning urine were collected within three days prior to the end of pre- and post-consumption periods. The primary endpoints of the study were the effects of FSM consumption on the increase in the number and relative abundance of Bifidobacterium and lactobacilli, and the secondary endpoints were its effects on changes in the number and relative abundance of other bacteria, increases in the concentrations of SCFAs and defecation frequency, and decreases in the concentrations of putrefactive products.
Treatment of faecal samples
Faecal samples were treated for DNA and RNA extraction as described previously (Kubota et al., 2010). Briefly, approximately 0.5 g faeces was collected into a sterile faecal-collection tube (Sarstedt AG & Co., Nümbrecht, Germany) containing 2 ml RNAlater (Ambion, Inc., Austin, TX, USA). The tubes were maintained at 4 °C, transferred to a laboratory, and stored at 4 °C until processing. Faecal samples were weighed, suspended in 9 volumes of RNAlater, and homogenised. The faecal homogenate was divided into two new tubes of 200 μl each. One was washed with phosphate-buffered saline (PBS) and stored at −80 °C until DNA extraction. The other was added to 1 ml PBS and centrifuged at 13,000× g for 5 min. The supernatant was discarded and the pellet was stored at −80 °C until RNA extraction.
Gut microbiota analysis
The relative abundance and diversity of faecal gut bacteria were determined by 16S ribosomal RNA (rRNA) gene amplicon sequencing as described previously (Kato-Kataoka et al., 2016). Briefly, DNA was extracted from faecal samples, and the V1-V2 region of the 16S rRNA gene of gut microbiota was amplified using the forward 27Fmod2-MiSeqV2 and reverse 338RMiSeqV2-001 primers on an ABI 7500 Real-Time PCR System (Life Technologies Japan Ltd., Tokyo, Japan) (Caporaso et al., 2012). The amplified DNA was purified using an AMPure XP Kit (Beckman Coulter, Brea, CA, USA), quantified using a Quant-iT PicoGreen dsDNA Assay Kit (Life Technologies Japan Ltd.), and sequenced on a MiSeq system (Illumina, San Diego, CA, USA) using a MiSeq Reagent Kit v2 (Illumina). The amplicon sequence reads were processed using Quantitative Insights Into Microbial Ecology (QIIME) 2 software (version. 2019. 01) (Bolyen et al., 2019), and taxonomy was assigned using the SILVA 138 reference database.
The total number of faecal bacteria was determined by quantitative real-time polymerase chain reaction (qPCR) using primer sets and DNA as a template on an ABI PRISM 7900HT Sequence Detection System (Life Technologies Japan Ltd.) as described previously (Shima et al., 2019). The number of two beneficial bacteria (Bifidobacterium and lactobacilli) and five pathogenic bacteria (Enterobacteriaceae, Staphylococcus, Pseudomonas, Clostridioides difficile, and Clostridium perfringens) in faeces was determined by quantitative reverse transcription (qRT)-PCR using the Yakult Intestinal Flora Scan (Matsuda et al., 2007). RNA was extracted from faecal samples for qRT-PCR using specific primer sets as described previously (Shima et al., 2019). qRT-PCR was performed on an ABI PRISM 7900HT Sequence Detection System (Life Technologies Japan Ltd.). The total number of lactobacilli was calculated as the sum of the counts of six lactobacilli subgroups and two species (Shima et al., 2019). The number of bacteria in the ‘not detected’ samples was assigned as half the detection limit of the corresponding primer sets.
Quantification of LcS in faeces
Detection of total LcS containing living and dying cells in faeces was performed via qPCR using an LcS-specific primer set and DNA as a template as described previously (Fujimoto et al., 2008). The number of living LcS in faeces was determined using a culture method as described previously (Kato-Kataoka et al., 2016). Briefly, approximately 0.5 g faeces was collected and suspended in 3 ml anaerobic dilution solution. The faecal samples were maintained at 4 °C, transferred to a laboratory, and stored at 4 °C. The faecal samples were weighed, suspended in 9 volumes of PBS, and cultured on lactitol-Lactobacillus selection (LBS)-vancomycin agar plates at 37 °C for 72 h. The colonies that formed on the agar plates were identified by colony PCR using the primer set described above to count the cfu of LcS.
Predictive metagenomic analysis
Predictive metagenomic analysis of the 16S rRNA gene of gut microbiota was performed using Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) 2 (Douglas et al., 2020).
Measurement of faecal and urinary parameters
The measurement of faecal and urinary parameters was outsourced to the Kyoto Institute of Nutrition & Pathology (Kyoto, Japan). Faecal and urinary samples were frozen immediately after collection, transferred to the Kyoto Institute of Nutrition & Pathology in a frozen state, and stored at −30 °C. The concentrations of faecal organic acids, such as succinate, lactate, formate, acetate, propionate, iso-butyrate, n-butyrate, iso-valerate, and n-valerate, were determined by ion-exclusion high-performance liquid chromatography as described previously (Tsukahara et al., 2014). Total SCFAs were calculated as the sum of the concentrations of succinate, lactate, formate, acetate, propionate, n-butyrate, and n-valerate, whereas total branched-chain fatty acids (BCFAs) were the sum of iso-butyrate and iso-valerate concentrations. Faecal moisture content was derived from the difference between the wet and dry weights of faeces. Faecal pH was measured with a B212 Twin pH Meter (Horiba Ltd., Kyoto, Japan). The concentrations of urinary putrefactive products, such as phenol, p-cresol, and indole, were determined by ultra-performance liquid chromatography tandem mass spectrometry as described previously (Kawase et al., 2019), and were normalised to urinary creatinine levels, which were assayed using a Creatinine Assay Kit (Oxford Biochemical Research, Rochester Hills, MI, USA). Total putrefactive products were calculated as the sum of the normalised urinary concentrations of phenol, p-cresol, and indole.
Questionnaire survey of defecation habits
The frequency of defecation was evaluated using a self-administrated questionnaire. The subjects were asked to record the timing of defecation every day, and the number of defecations and defecation days per week were calculated.
Statistical analysis
Data are expressed as the mean ± standard deviation or the median. All data were analysed using R ver. 3.6.1. Background data between the groups were analysed with Fisher’s exact test for the ratio of males/females or one-way analysis of variance for the other parameters. Taxonomic data from the 16S rRNA amplicon sequencing, qRT-PCR, and qPCR were analysed by the Wilcoxon signed-rank test within groups and by the Mann-Whitney U test between groups. Faecal and urinary data were analysed by a paired t-test within groups and by Student’s t-test between groups. Defecation data were analysed by Dunnett’s multiple comparisons test within groups and by Student’s t-test between groups. Predicted metagenomic data were analysed by the Wilcoxon signed-rank test within groups. Correlation analysis was performed using Spearman’s rank correlation coefficient. Significance was set at

Participant flow diagram. FSM = fermented soy milk containing Lacticaseibacillus paracasei strain Shirota.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
3 Results
Subjects
Figure 1 shows the participant flow diagram of this study. Among the 365 participants who gave consent to participate, 120 healthy subjects with a low Bifidobacterium faecal population were screened, considering health conditions and the inclusion criteria. The average number of Bifidobacterium was 9.45 ± 1.0 (median: 9.69)
Background data of the subjects included in the analysis1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
Detection of LcS and compliance
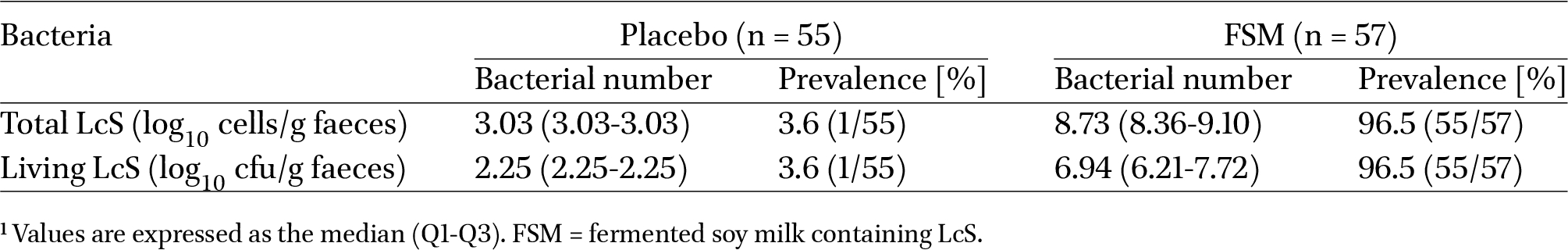
The self-reported compliance rate regarding the consumption of the test and placebo beverages was >99.0% in both groups. Table 2 shows the number of total and living LcS in faeces in the post-consumption period in both groups. In the post-consumption period in the FSM group, living LcS was detected in the faeces of 55 subjects, ranging from 6.21 to 7.72
Number of Lacticaseibacillus paracasei strain Shirota (LcS) in faeces1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
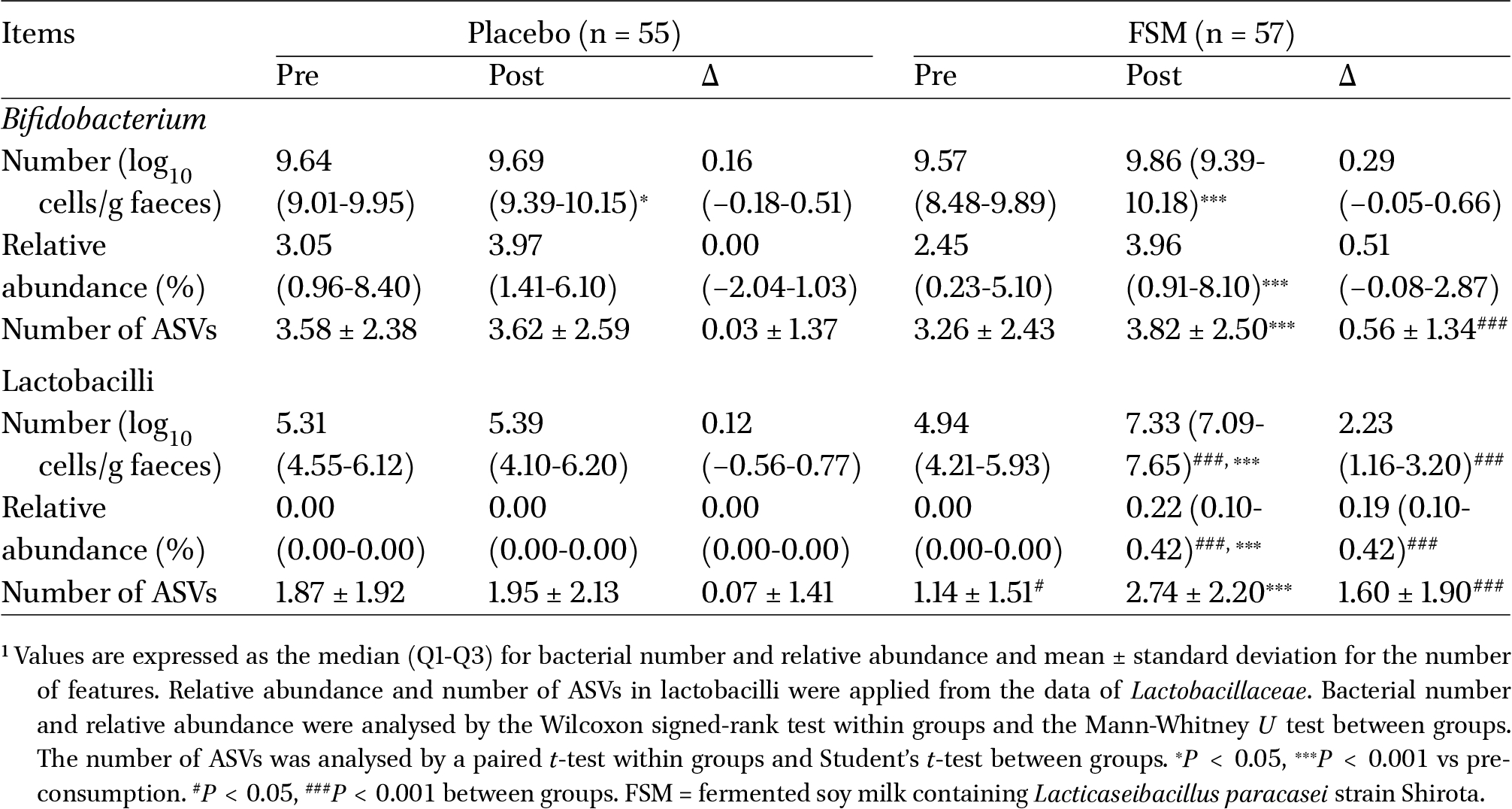
Number, relative abundance, and number of ASVs of Bifidobacterium and lactobacilli (Lactobacillaceae) in faeces1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
Gut microbiota
Table 3 shows the effects of the consumption of the test and placebo beverages on the number, relative abundance, and number of amplicon sequence variants (ASVs) of Bifidobacterium and lactobacilli in faeces as the primary endpoints. In both groups, the number of Bifidobacterium was significantly increased in the post-consumption period compared with the pre-consumption period (placebo group:
The effects of the consumption of the test and placebo beverages on the composition of gut microbiota were evaluated by 16S rRNA amplicon analysis. Supplementary Figure S1 shows the effects on the observed ASVs, which is an index of alpha-diversity. In the FSM group alone, the observed ASVs were significantly increased in the post-consumption period compared with the pre-consumption period (
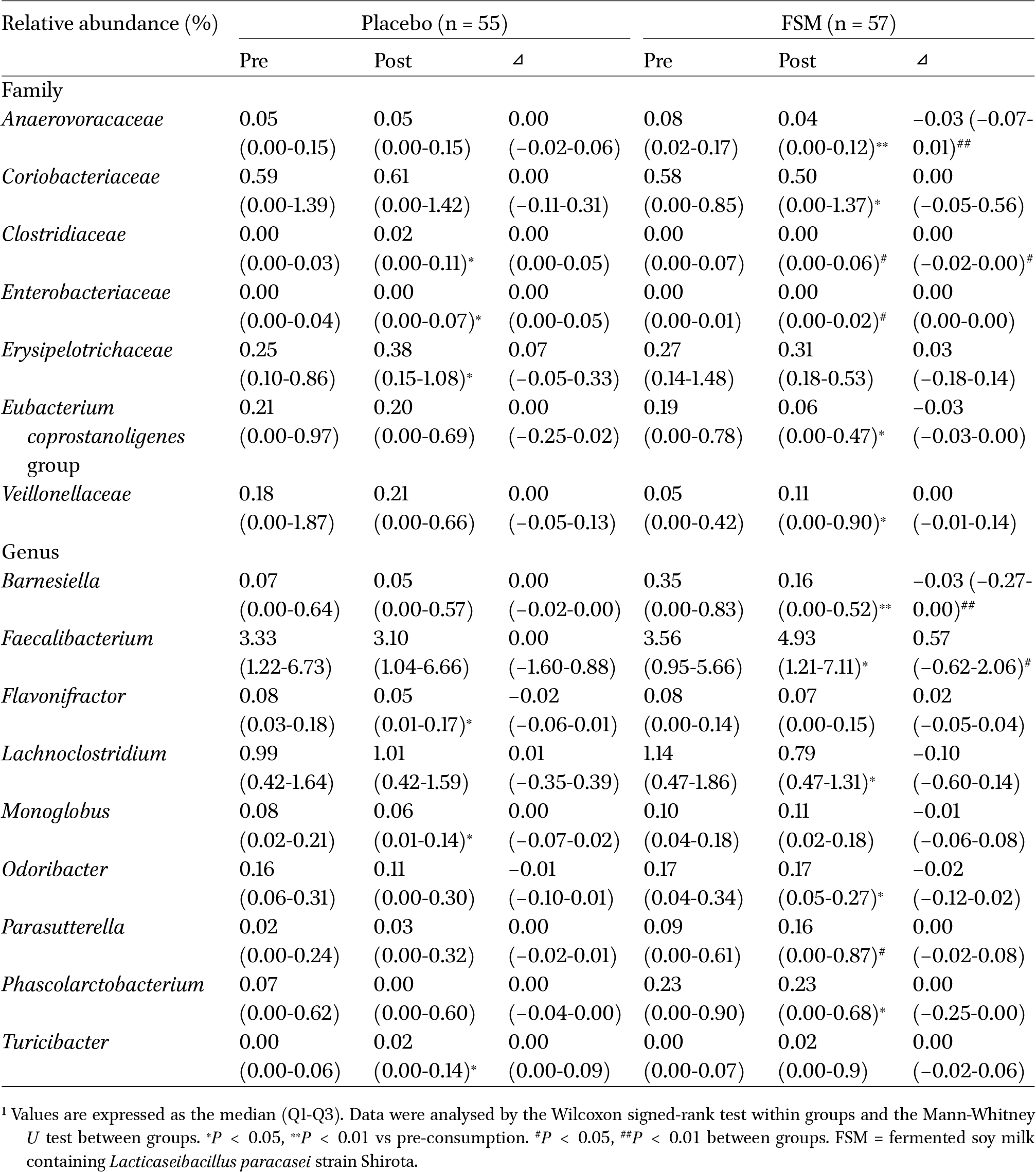
Bacterial families and genera that were significantly different in their relative abundance following the consumption of the test beverages, with the exception of Bifidobacterium and lactobacilli1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
Supplementary Table S2 shows the effects of the consumption of the test and placebo beverages on low-level pathogenic bacteria and the total number of bacteria, which were analysed using the Yakult Intestinal Flora Scan because of the difficulty of detection using 16S rRNA amplicon analysis. The FSM group had a marginally lower number of Enterobacteriaceae (
Gut environment
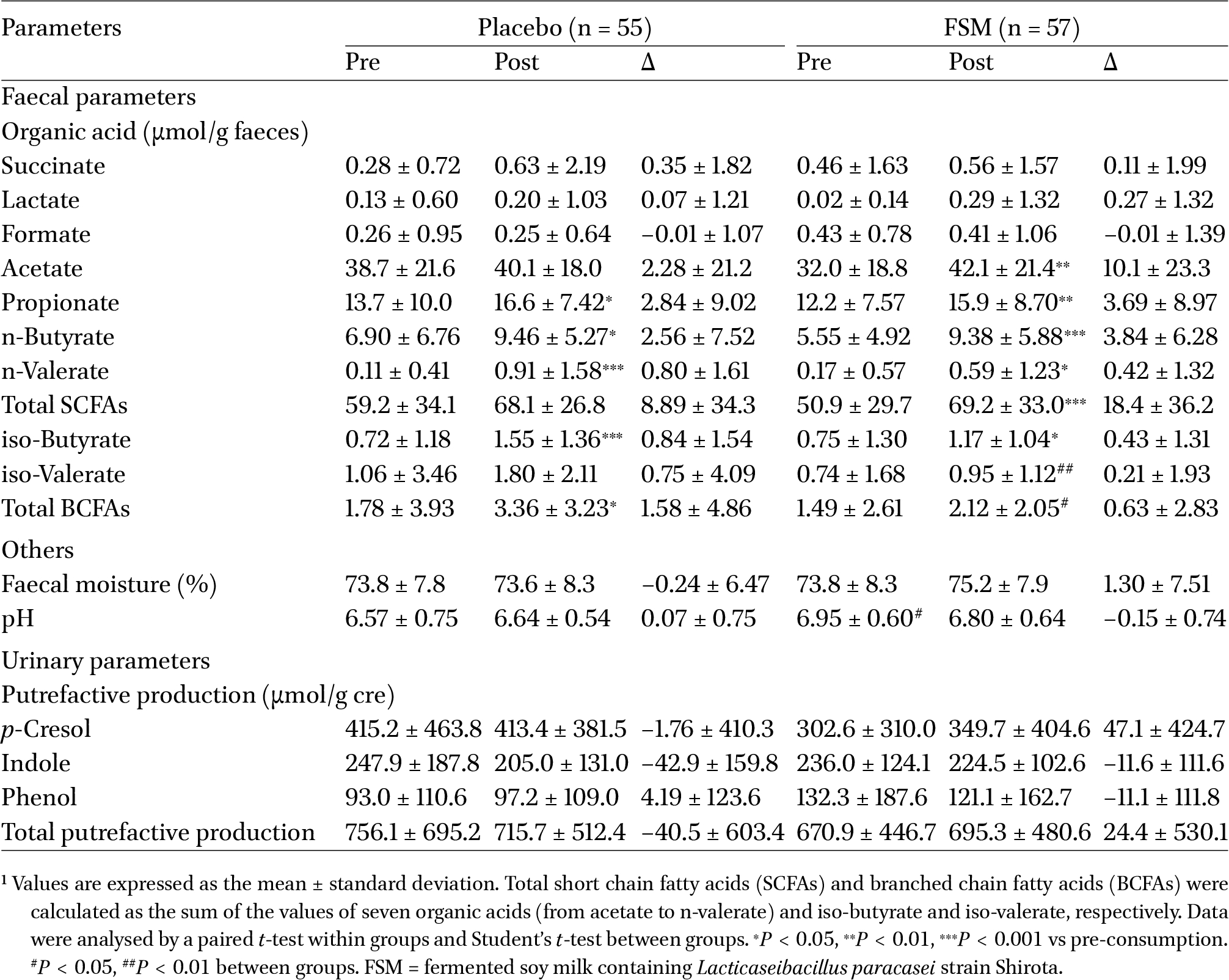
Table 5 lists the effects of the consumption of the test and placebo beverages on faecal and urinary parameters, such as organic acids and putrefactive products. Compared with the pre-consumption period, the faecal concentrations of propionate (placebo:
Faecal and urinary parameters1
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
Defecation habits
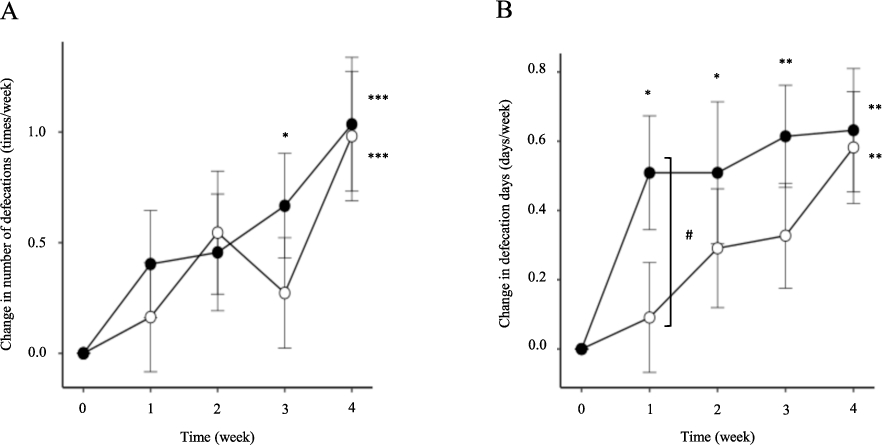
Figure 2 depicts the time course of the change in the number of defecations and defecation days per week during the consumption period. Both groups reported an average of about 5 defecation days per week in the pre-consumption period. The placebo group exhibited a significant increase in both parameters at 4 weeks compared with the pre-consumption value (0 weeks). Conversely, the FSM group reported significant increases in the number of defecations after 3 weeks and in the number of defecation days after 1 week compared with the pre-consumption values (0 weeks). In particular, the change in the number of defecation days was marginally higher at 1 week in the FSM group compared with the placebo group (
Time course of the change in the number of defecations (A) and defecation days (B) per week. Data are expressed as the mean ± standard error and analysed using Dunnett’s multiple comparisons test within groups and Student’s t-test between groups. *
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
Subgroup analysis
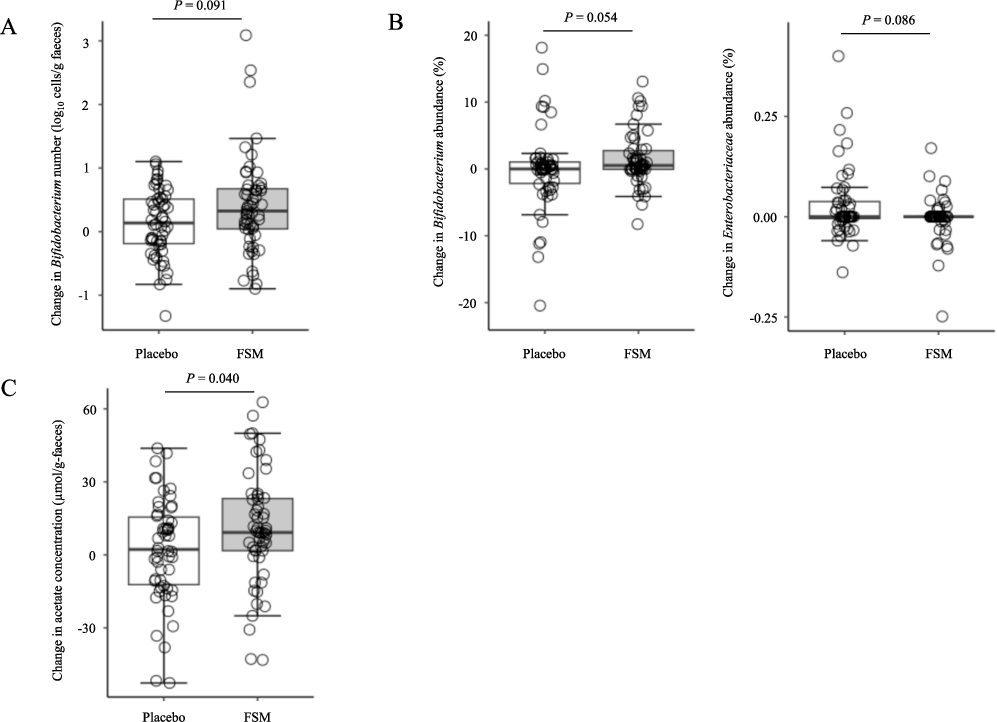
To accurately evaluate the effect of FSM on the gut microbiota and intestinal environment, subgroup analysis performed on 109 subjects, excluding 2 subjects in the FSM group in whom no living LcS was detected and 1 subject in the SM group in whom living LcS was detected. During the consumption period, the FSM group showed marginally greater changes in the number and relative abundance of Bifidobacterium in faeces, which served as the primary endpoint, compared with the SM group, although no significant differences were observed (Figure 3A and B, number:
Effect of Lacticaseibacillus paracasei strain Shirota (LcS) that reached the gut alive on the gut microbiota and faecal parameters. Comparison of the changes in number of Bifidobacterium (A), relative abundance of Bifidobacterium (B), and acetate (C). Data are expressed as the median value and interquartile range and analysed using the Mann-Whitney U test between groups.
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
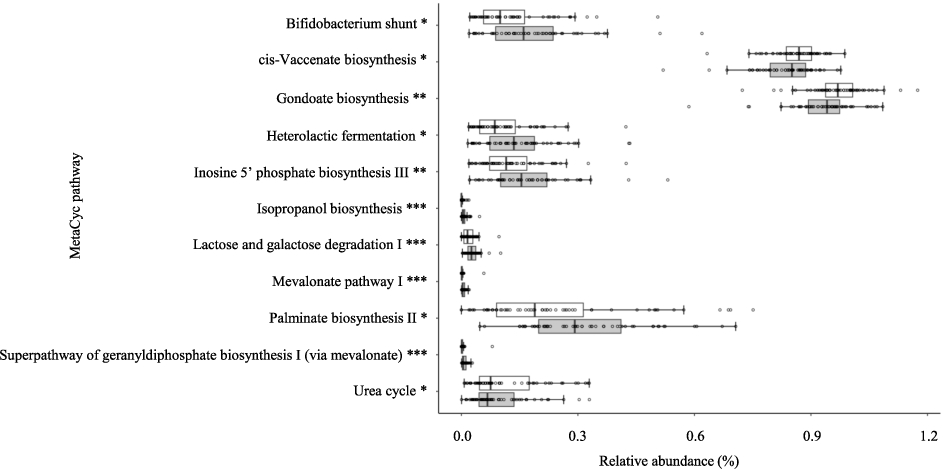
To investigate how FSM may be involved in the beneficial effects mentioned above, predictive metagenomic analysis was performed using PICRUSt2 to evaluate the effect on the metagenomic profile of the gut microbiota. Figure 4 shows the significantly upregulated or downregulated pathways in the gut microbiota during the consumption period in the FSM group, in which living LcS was detected. We detected eight significantly upregulated pathways, many of which were related to energy production or glucose metabolism, and three significantly downregulated pathways, two of which were involved in the synthesis of unsaturated fatty acids. In contrast, the SM group, in which no living was LcS detected, had no significantly changed pathways in the gut microbiota during the consumption period.
Predicted metagenomic functions with significant up- or downregulation in the group with Lacticaseibacillus paracasei strain Shirota (LcS) that reached the gut alive. The relative abundance of each metagenomic function predicted based on the MetaCyc database is expressed as the median value and interquartile range. Data were analysed using the Wilcoxon sighed-rank test adjusted for the Benjamini-Hochberg false discovery rate. *
Citation: Beneficial Microbes 15, 2 (2024) ; 10.1163/18762891-bja00003
4 Discussion
The gut microbiota plays important roles in the maintenance of health and the prevention of diseases in hosts. It has been reported that the modulation of the gut microbiota, especially an increase in probiotic and/or intrinsic bifidobacteria and lactobacilli in the gut, exerts beneficial effects on the host (Havenaar, 2011).
The present study demonstrated that both FSM and SM group had significant increases in the number of Bifidobacterium (Table 3). It has also been reported that the number of faecal Bifidobacterium is significantly increased by the daily consumption of SM containing soy oligosaccharides, such as raffinose and stachyose (Fujisawa et al., 2017), indicating that soy oligosaccharides contributed to the increase of Bifidobacterium in the placebo group of the present study. In contrast, the FSM group had a greater increase in the relative abundance and number of ASVs of Bifidobacterium in faeces alone, in particularly, the changed level of ASVs for Bifidobacterium was remarkable compared to the SM group. The richness of Bifidobacterium diversity is crucial as it allows Bifidobacterium to occupy a niche in the gut microbiota, which is a positive change for improving gut microbiota. These suggest that FSM has a greater impact on Bifidobacterium as primary endpoint in the gut microbiota compared with SM. In addition, the FSM group alone exhibited significant increases in the number, relative abundance, and number of ASVs of lactobacilli in faeces as the other primary endpoints during the consumption period; moreover, we observed significant differences in these parameters between both groups in the post-consumption period (Table 3). These findings indicate that the observed increases depended on the daily consumption of FSM and were attributable to LcS in FSM.
The present study also revealed that the FSM group alone had significant increases or decreases in the relative abundance of several bacterial groups in the gut microbiota during the consumption period (Table 4). In particular, Faecalibacterium, which was significantly increased in the only FSM group, is a typical butyrate-producing bacterium that has been suggested to be a good candidate next-generation probiotic because its faecal level is lower in patients with inflammatory bowel disease, irritable bowel syndrome, and colorectal cancer (Martı́n et al., 2017). Interestingly, the daily consumption of FSM significantly increased not only the relative abundance of Faecalibacterium but also the faecal concentration of n-butyrate (Table 5). Our previous pilot study demonstrated that the relative abundance of Enterobacteriaceae in the gut microbiota was significantly decreased during the consumption period in the FSM group (Nagino et al., 2018). However, its relative abundance did not change in the present study (Table 3). There was a remarkable difference in the relative abundance of Enterobacteriaceae in the pre-consumption period between the pilot study (0.14 [0.02-0.75]; median [Q1-Q3]) and the present study (0.00 [0.00-0.01]), suggesting that this discrepancy can be attributed to differences in the subjects included in both studies. Nevertheless, in the present study, the FSM group had a significantly and marginally lower relative abundance and number of Enterobacteriaceae during the post-consumption period, respectively, compared with the SM group. A significant increase in the relative abundance of Enterobacteriaceae was observed only in the SM group, which may have been suppressed in the FSM group. These observations suggest that daily consumption of FSM contributes to improvement of the gut microbiota more effectively than does SM.
The present study revealed that the FSM group had a greater increase in the faecal concentrations of acetate, propionate, n-butyrate, and total SCFAs compared with the SM group, and that increases in acetate and total SCFAs were detected in the FSM group alone (Table 5). There is good agreement between the present study and several previous clinical studies (Wang et al., 2015; Nagata et al., 2016) showing that the daily consumption of fermented milk containing LcS contributes to an increase in the number of Bifidobacterium and acetate concentration in faeces. SCFAs, mainly acetate, propionate, and n-butyrate, are metabolites of gut microbiota that utilise sugar as a substrate, and have various beneficial functions in the host, as well as being used as an energy source (Den Besten et al., 2013). In contrast, the SM group had significant increases in the faecal concentrations of total BCFAs and some SCFAs (Table 5). BCFAs are metabolites derived from branched-chain amino acids, such as valine, leucine, and isoleucine, and have been reported as markers of the protein metabolism involved in the production of putrefactive products, such as ammonia and amines (Macfarlane et al., 1992). Although only beneficial SCFAs were produced in the FSM group, not only SCFAs but also harmful BCFAs were produced in the SM group, indicating that the daily consumption of FSM was more effective than SM in improving the intestinal environment.
This study found that the FSM and SM groups had significant increases in the number of defecations and defecation days per week, and that these increases were observed earlier in the FSM group compared with the SM group (Figure 2). It has been reported that defecation habits are associated with gut microbiota and the intestinal environment (Kim et al., 2015, 2020). It is known that constipation-prone individuals have a lower number of Bifidobacterium and lactobacilli in faeces compared with healthy individuals, and that an increase in faecal SCFAs improves the symptoms of constipation. Therefore, these findings support the contention that the daily consumption of FSM and SM improves the gut microbiota and intestinal environment but that FSM acts more effectively than SM towards this improvement, although the detailed mechanisms underlying this observation remain unknown. In addition, the subjects enrolled in this study were in the normal range, with an average number of defecation days per week of about 5. In the future, it may be possible to clarify the difference between FSM and SM by targeting subjects with a stronger tendency towards constipation.
In this study, one subject in the SM group was found to have living LcS (Table 2). A clinical study reported that living LcS could be detected in faeces up to 2 weeks after stopping intake of LcS (Kato-Kataoka et al., 2016). In addition, products containing LcS are easily accessible in Japan; therefore, it is possible that the subject in the SM group may have consumed such products during the study period. In contrast, 2 subjects in the FSM group were found to have no living LcS. Other clinical studies have similarly reported rare cases in which LcS was not detected despite subjects ingesting live LcS (Kato-Kataoka et al., 2016). In the present study, the 2 subjects in the FSM group in whom LcS was not detected were checked not only by a culture method but also by qPCR, which can identify both living and dead cells. Furthermore, Lacticaseibacillus paracasei containing LcS was also not detected in these subjects. It is possible that they did not consume the test beverage, but the details are unknown.
To accurately examine the effects of FSM on the gut microbiota and intestinal environment, subgroup analyses were performed in the SM and FSM groups, excluding the 3 subjects with inconsistent LcS detection. These analyses showed that only the FSM group had significant increases in the number and relative abundance of Bifidobacterium in faeces during the consumption period, and that these changes were marginally higher in the FSM group than in the SM group (Figure 3A and B). In contrast, the increase in the abundance of Enterobacteriaceae observed in the SM group was suppressed in the FSM group during the consumption period, and the level was marginally lower in the FSM group than in the SM group (Figure 3B). These analyses also revealed that only the FSM group showed increases in faecal concentrations of acetate and total SCFAs during the consumption period, and that the changed concentration of faecal acetate was significantly higher in the FSM group than in the SM group (Figure 3C). Acetate is reported to reduce the number of harmful bacteria such as Enterobacteriaceae (Wang et al., 2015), supporting the findings of the present study. Furthermore, predictive metagenomic analysis using PICRUSt2 revealed that only the FSM group showed significantly upregulated metabolic pathways in the gut microbiota, including lactose and galactose degradation I, which is involved in glucose metabolism; heterolactic fermentation, which is involved in SCFA production; and Bifidobacterium shunt, which is involved in both glucose metabolism and SCFA production in the post-consumption period (Figure 4). It is considered that the consumption of FSM induces the upregulation of these metabolic pathways through changes in the gut microbiota and stimulates the production of SCFAs (especially acetate) in the gut.
Limitations
This study had several limitations that should be considered. First, the effects of lifestyle habits, such as diet, smoking, alcohol intake, and exercise, during the study period were not considered. Among them, diet has a considerable impact on the gut microbiota in a short period (Sugimoto et al., 2020). To further clarify the effects of the test beverages on the gut microbiota, it is necessary to analyse the data in light of these effects. Second, the effect of LcS that reached the gut alive on the metagenome profile was evaluated exclusively via prediction using 16S rRNA sequence information, and functional genes were not analysed based on shotgun metagenomic sequencing and their expression was not analysed. Because probiotics have very complex interactions not only with the host but also with many gut bacteria, the detailed mechanism underlying the improvement of the gut bacteria and intestinal environment of the host remains unknown, even for LcS. These analyses are expected to provide important knowledge for the elucidation of this complex mechanism.
5 Conclusions
This study demonstrated that the daily consumption of FSM for 4 weeks balanced the gut microbiota and contributed to an improvement of the intestinal environment and defecation habits in healthy Japanese adults, and that these effects may be greater than those afforded by SM alone. It is considered that daily consumption of FSM may help to maintain health and prevent diseases.
Corresponding author; e-mail: chiaki-kaga@yakult.co.jp
Supplementary material
Supplementary material is available online at: https://doi.org/10.6084/m9.figshare.25118018
Table S1. Composition of test beverages.
Table S2. Number of total bacteria and pathogenic bacteria.
Figure S1. Effect on observed ASVs as an index of alpha-diversity.
Acknowledgements
The authors thank all subjects who participated in this study. We also thank Mr. Michitoshi Anbe, Mr. Osamu Watanabe, Mr. Toshiyuki Harada, Mr. Daisuke Shigihara, and Ms. Yuki Kataoka for production of the test beverages as well as Mr. Toshio Sasai for support with the predicted metagenomic analysis and statistical analysis.
Authors’ contribution
CK, TN, AT, and KS designed the study. CK and TN acquired and analysed data. AG, AT, KM, YY, and SK supervised the project. CK prepared the first draft of the manuscript. AG, KM, YY, and KS reviewed the draft. All authors read and approved the final manuscripts.
Conflict of interest
This study was supported by a grant from the Yakult Honsha Co., Ltd. All authors are employed by Yakult Honsha Co., Ltd.
References
Aizawa, E., Tsuji, H., Asahara, T., Takahashi, T., Teraishi, T., Yoshida, S., Koga, N., Hattori, K., Ota, M. and Kunugi, H., 2019. Bifidobacterium and Lactobacillus counts in the gut microbiota of patients with bipolar disorder and healthy controls. Frontiers in Psychiatry 10: 730. https://doi.org/10.3389%2Ffpsyt.2018.00730
Bolyen, E., Rideout, J.R., Dillon, M.R., Bokulich, N.A., Abnet, C.C., Al-Ghalith, G.A., Alexander, H., Alm, E.J., Arumugam, M., Asnicar, F., Bai, Y., Bisanz, J.E., Bittinger, K., Brejnrod, A., Brislawn, C.J., Brown, C.T., Callahan, B.J., Caraballo-Rodrı́guez, A.M., Chase, J., Cope, E.K., Da Silva, R., Diener, C., Dorrestein, P.C., Douglas, G.M., Durall, D.M., Duvallet, C., Edwardson, C.F., Ernst, M., Estaki, M., Fouquier, J., Gauglitz, J.M., Gibbons, S.M., Gibson, D.L., Gonzalez, A., Gorlick, K., Guo, J., Hillmann, B., Holmes, S., Holste, H., Huttenhower, C., Huttley, G.A., Janssen, S., Jarmusch, A.K., Jiang, L., Kaehler, B.D., Kang, K.B., Keefe, C.R., Keim, P., Kelley, S.T., Knights, D., Koester, I., Kosciolek, T., Kreps, J., Langille, M.G.I., Lee, J., Ley, R., Liu, Y.-X., Loftfield, E., Lozupone, C., Maher, M., Marotz, C., Martin, B.D., McDonald, D., McIver, L.J., Melnik, A.V., Metcalf, J.L., Morgan, S.C., Morton, J.T., Naimey, A.T., Navas-Molina, J.A., Nothias, L.F., Orchanian, S.B., Pearson, T., Peoples, S.L., Petras, D., Preuss, M.L., Pruesse, E., Rasmussen, L.B., Rivers, A., Michael II, S.R., Rosenthal, P., Segata, N., Shaffer, M., Shiffer, A., Sinha, R., Song, S.J., Spear, J.R., Swafford, A.D., Thompson, L.R., Torres, P.J., Trinh, P., Tripathi, A., Turnbaugh, P.J., Ul-Hasan, S., van der Hooft, J.J.J., Vargas, F., Vázquez-Baeza, Y., Vogtmann, E., von Hippel, M., Walters, W., Wan, Y., Wang, M., Warren, J., Weber, K.C., Williamson, C.H.D., Willis, A.D., Xu, Z.Z., Zaneveld, J.R., Zhang, Y., Zhu, Q., Knight, R. and Caporaso, J.G., 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnology 37: 852-857. https://doi.org/10.1038/s41587-019-0209-9
Caporaso, J.G., Lauber, C.L., Walters, W.A., Berg-Lyons, D., Huntley, J., Fierer, N., Owens, S.M., Betley, J., Fraser, L., Bauer, M., Gormley, N., Gilbert, J.A., Smith, G. and Knight, R., 2012. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME Journal 6: 1621-1624. https://doi.org/10.1038/ismej.2012.8
Carbonero, F., Benefiel, A.C., Alizadeh-Ghamsari, A.H. and Gaskins, H.R., 2012. Microbial pathways in colonic sulfur metabolism and links with health and disease. Frontiers in Physiology 3: 448. https://doi.org/10.3389/fphys.2012.00448
Den Besten, G., Van Eunen, K., Groen, A.K., Venema, K., Reijngoud, D.J. and Bakker, B.M., 2013. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. Journal of Lipid Research 54: 2325-2340. https://doi.org/10.1194%2Fjlr.R036012
Douglas, G.M., Maffei, V.J., Zaneveld, J.R., Yurgel, S.N., James, R., Taylor, C.M., Huttenhower, C. and Langille, M.G.I., 2020. PICRUSt2 for prediction of metagenome functions. Nature Biotechnology 38: 685-688. https://doi.org/10.1038/s41587-020-0548-6
Evenepoel, P., Meijers, B.K.I., Bammens, B.R.M. and Verbeke, K., 2009. Uremic toxins originating from colonic microbial metabolism. Kidney International 76: S12-S19. https://doi.org/10.1038/ki.2009.402
Fujimoto, J., Matsuki, T., Sasamoto, M., Tomii, Y. and Watanabe, K., 2008. Identification and quantification of Lactobacillus casei strain Shirota in human feces with strain-specific primers derived from randomly amplified polymorphic DNA. International Journal of Food Microbiology 126: 210-215. https://doi.org/10.1016/j.ijfoodmicro.2008.05.022
Fujisawa, T., Ohashi, Y., Shin, R., Narai-Kanayama, A. and Nakagaki, T., 2017. The effect of soymilk intake on the fecal microbiota, particularly Bifidobacterium species, and intestinal environment of healthy adults: a pilot study. Bioscience of Microbiota, Food and Health 36: 33-37. https://doi.org/10.12938/bmfh.16-017
Fukuda, S., Toh, H., Hase, K., Oshima, K., Nakanishi, Y., Yoshimura, K., Tobe, T., Clarke, J.M., Topping, D.L., Suzuki, T., Taylor, T.D., Itoh, K., Kikuchi, J., Morita, H., Hattori, M. and Ohno, H., 2011. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 469: 543-549. https://doi.org/10.1038/nature09646
Furusawa, Y., Obata, Y., Fukuda, S., Endo, T.A., Nakato, G., Takahashi, D., Nakanishi, Y., Uetake, C., Kato, K., Kato, T., Takahashi, M., Fukuda, N.N., Murakami, S., Miyauchi, E., Hino, S., Atarashi, K., Onawa, S., Fujimura, Y., Lockett, T.J., Clarke, J.M., Topping, D.L., Tomita, M., Hori, S., Ohara, O., Morita, T., Koseki, H., Kikuchi, J., Honda, K., Hase, K. and Ohno, H., 2013. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504: 446-450. https://doi.org/10.1038/nature12721
Gibson, G.R., Probert, H.M., Van Loo, J., Rastall, R.A. and Roberfroid, M.B., 2004. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutrition Research Reviews 17: 259-275. https://doi.org/10.1079/nrr200479
Gómez-Zorita, S., González-Arceo, M., Fernández-Quintela, A., Eseberri, I., Trepiana, J. and Portillo, M.P., 2020. Scientific evidence supporting the beneficial effects of isoflavones on human health. Nutrients 12: 1-25. https://doi.org/10.3390/nu12123853
Hamer, H.M., Jonkers, D.M.A.E., Renes, I.B., Steven, A.L.W., Vanhoutvin, A.K., Troost, F.J., Venema, K. and Brummer, R.-J.M., 2010. Butyrate enemas do not affect human colonic MUC2 and TFF3 expression. European Journal of Gastroenterology and Hepatology 22: 1134-1140. https://doi.org/10.1097/meg.0b013e32833a6ca0
Havenaar, R., 2011. Intestinal health functions of colonic microbial metabolites: a review. Beneficial Microbes 2: 103-114. https://doi.org/10.3920/bm2011.0003
Hayakawa, K., Mizutani, J., Wada, K., Masai, T., Yoshihara, I. and Mitsuoka, T., 1990. Effects of soybean oligosaccharides on human faecal flora. Microbial Ecology in Health and Disease 3: 293-303. https://doi.org/10.1271/nogeikagaku1924.66.127
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C. and Sanders, M.E., 2014. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology and Hepatology 11: 506-514. https://doi.org/10.1038/nrgastro.2014.66
Hiutung, C., Khosravi, A., Kusumawardhani, I.P. and Kwon, A.H.K., 2016. Gene-microbiota interactions contribute to the pathogenesis of inflammatory bowel disease. Science 352: 1116-1120. https://doi.org/10.1126/science.aad9948
Inoguchi, S., Ohashi, Y., Narai-Kanayama, A., Aso, K., Nakagaki, T. and Fujisawa, T., 2012. Effects of non-fermented and fermented soybean milk intake on faecal microbiota and faecal metabolites in humans. International Journal of Food Sciences and Nutrition 63: 402-410. https://doi.org/10.3109/09637486.2011.630992
Ishikawa, H., Akedo, I., Otani, T., Suzuki, T., Nakamura, T., Takeyama, I., Ishiguro, S., Miyaoka, E., Sobue, T. and Kakizoe, T., 2005. Randomized trial of dietary fiber and Lactobacillus casei administration for prevention of colorectal tumors. International Journal of Cancer 116: 762-767. https://doi.org/10.1002/ijc.21115
Kato-Kataoka, A., Nishida, K., Takada, M., Kawai, M., Kikuchi-Hayakawa, H., Suda, K., Ishikawa, H., Gondo, Y., Shimizu, K., Matsuki, T., Kushiro, A., Hoshi, R., Watanabe, O., Igarashi, T., Miyazaki, K., Kuwano, Y. and Rokutan, K., 2016. Fermented milk containing Lactobacillus casei strain Shirota preserves the diversity of the gut microbiota and relieves abdominal dysfunction in healthy medical students exposed to academic stress. Applied and Environmental Microbiology 82: 3649-3658. https://doi.org/10.1128/aem.04134-15
Kawase, T., Kawakami, K., Harada, K., Chonan, O. and Tsukahara, T., 2019. Simultaneous analysis of glucuronyl- and sulpho-conjugates of intestinal putrefactive compounds in human urine by ultra-performance liquid chromatography tandem mass spectrometry. Chromatography 40: 9-18. https://doi.org/10.15583/jpchrom.2018.017
Kim, M.G., Jo, K., Chang, Y.B., Suh, H.J. and Hong, K.B., 2020. Changes in the gut microbiome after galacto-oligosaccharide administration in loperamide-induced constipation. Journal of Personalized Medicine 10: 161. https://doi.org/10.3390/jpm10040161.
Kim, S.E., Choi, S.C., Park, K.S., Park, M.I., Shin, J.E., Lee, T.H., Jung, K.W., Koo, H.S. and Myung, S.J., 2015. Change of fecal flora and effectiveness of the short-term VSL#3 probiotic treatment in patients with functional constipation. Journal of Neurogastroenterology and Motility 21: 111-120. https://doi.org/10.5056/jnm14048
Kubota, H., Tsuji, H., Matsuda, K., Kurakawa, T., Asahara, T. and Nomoto, K., 2010. Detection of human intestinal catalase-negative, gram-positive cocci by rRNA-targeted reverse transcription-PCR. Applied and Environmental Microbiology 76: 5440-5451. https://doi.org/10.1128/aem.03132-09
Macfarlane, G.T., Gibson, G.R., Beatty, E. and Cummings, J.H., 1992. Estimation of short-chain fatty acid production from protein by human intestinal bacteria based on branched-chain fatty acid measurements. FEMS Microbiology Letters 101: 81-88. https://doi.org/10.1016/0378-1097(92)90829-D
Markowiak-Kopeć, P. and Śliżewska, K., 2020. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 12: 1107. https://doi.org/10.3390/nu12041107
Martı́n, R., Miquel, S., Benevides, L., Bridonneau, C., Robert, V., Hudault, S., Chain, F., Berteau, O., Azevedo, V., Chatel, J.M., Sokol, H., Bermúdez-Humarán, L.G., Thomas, M. and Langella, P., 2017. Functional characterization of novel Faecalibacterium prausnitzii strains isolated from healthy volunteers: a step forward in the use of F. prausnitzii as a next-generation probiotic. Frontiers in Microbiology 8: 1226. https://doi.org/10.3389/fmicb.2017.01226
Matsuda, K., Tsuji, H., Asahara, T., Kado, Y. and Nomoto, K., 2007. Sensitive quantitative detection of commensal bacteria by rRNA-targeted reverse transcription-PCR. Applied and Environmental Microbiology 73: 32-39. https://doi.org/10.1128/aem.01224-06
Matsumoto, K., Takada, T., Shimizu, K., Kado, Y., Kawakami, K., Makino, I., Yamaoka, Y., Hirano, K., Nishimura, A., Kajimoto, O. and Nomoto, K., 2006. The effects of a probiotic milk product containing Lactobacillus casei strain Shirota irota on the defecation frequency and the intestinal microflora of sub-optimal health state volunteers: a randomized placebo-controlled cross-over study. Bioscience and Microflora 25: 39-48. https://doi.org/10.1016/j.jbiosc.2010.05.016
Matsumoto, K., Takada, T., Shimizu, K., Moriyama, K., Kawakami, K., Hirano, K., Kajimoto, O. and Nomoto, K., 2010. Effects of a probiotic fermented milk beverage containing Lactobacillus casei strain Shirota on defecation frequency, intestinal microbiota, and the intestinal environment of healthy individuals with soft stools. Journal of Bioscience and Bioengineering 110: 547-552. https://doi.org/10.1016/j.jbiosc.2010.05.016
Mueller, S.O., Simon, S., Chae, K., Metzler, M. and Korach, K.S., 2004. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicological Sciences 80: 14-25. https://doi.org/10.1093/toxsci/kfh147
Nagao, F., Nakayama, M., Muto, T. and Okumura, K., 2000. Effects of a fermented milk drink containing Lactobacillus casei strain Shirota on the immune system in healthy human subjects. Bioscience, Biotechnology, and Biochemistry 64: 2706-2708. https://doi.org/10.1271/bbb.64.2706
Nagata, S., Asahara, T., Wang, C., Suyama, Y., Chonan, O., Takano, K., Daibou, M., Takahashi, T., Nomoto, K. and Yamashiro, Y., 2016. The effectiveness of Lactobacillus beverages in controlling infections among the residents of an aged care facility: a randomized placebo-controlled double-blind trial. Annals of Nutrition and Metabolism 68: 51-59. https://doi.org/10.1159/000442305
Nagino, T., Kaga, C., Kano, M., Masuoka, N., Anbe, M., Moriyama, K., Maruyama, K., Nakamura, S., Shida, K. and Miyazaki, K., 2018. Effects of fermented soymilk with Lactobacillus casei Shirota on skin condition and the gut microbiota: a randomised clinical pilot trial. Beneficial Microbes 9: 209-218. https://doi.org/10.3920/bm2017.0091
Nagino, T., Kano, M., Masuoka, N., Kaga, C., Anbe, M., Miyazaki, K., Kamachi, K., Isozaki, M., Suzuki, C., Kasuga, C. and Tanaka, A., 2015. Intake of a fermented soymilk beverage containing moderate levels of isoflavone aglycones enhances bioavailability of isoflavones in healthy premenopausal Japanese women: a double-blind, placebo-controlled, single-dose, crossover trial. Bioscience of Microbiota, Food and Health 35: 9-17. https://doi.org/10.12938/bmfh.2015-011
Naito, S., Koga, H., Yamaguchi, A., Fujimoto, N., Hasui, Y., Kuramoto, H., Iguchi, A., Kinukawa, N. and Kyushu University Urological Oncology Group, 2008. Prevention of recurrence with epirubicin and lactobacillus casei after transurethral resection of bladder cancer. Journal of Urology 179: 485-490. https://doi.org/10.1016/j.juro.2007.09.031
Nicholson, J.K., Holmes, E., Kinross, J., Burcelin, R., Gibson, G., Jia, W. and Pettersson, S., 2012. Host-gut microbiota metabolic interactions. Science 336: 1262-1267. https://doi.org/10.1126/science.1223813
Rajilić-Stojanović, M., Smidt, H. and De Vos, W.M., 2007. Diversity of the human gastrointestinal tract microbiota revisited. Environmental Microbiology 9: 2125-2136. https://doi.org/10.1111/j.1462-2920.2007.01369.x
Shiba, T., Kawakami, K., Sasaki, T., Makino, I., Kato, I., Kobayashi, T., Uchida, K. and Kaneko, K., 2014. Effects of intestinal bacteria-derived p-cresyl sulfate on Th1-type immune response in vivo and in vitro. Toxicology and Applied Pharmacology 274: 191-199. https://doi.org/10.1016/j.taap.2013.10.016
Shida, K., Sato, T., Iizuka, R., Hoshi, R., Watanabe, O., Igarashi, T., Miyazaki, K., Nanno, M. and Ishikawa, F., 2017. Daily intake of fermented milk with Lactobacillus casei strain Shirota reduces the incidence and duration of upper respiratory tract infections in healthy middle-aged office workers. European Journal of Nutrition 56: 45-53. https://doi.org/10.1007/s00394-015-1056-1
Shima, T., Amamoto, R., Kaga, C., Kado, Y., Sasai, T., Watanabe, O., Shiinoki, J., Iwazaki, K., Shigemura, H. and Tsuji, H., 2019. Association of life habits and fermented milk intake with stool frequency, defecatory symptoms and intestinal microbiota in healthy Japanese adults. Beneficial Microbes 10: 841-854. https://doi.org/10.3920/bm2019.0057
Sugimoto, T., Shima, T., Amamoto, R., Kaga, C., Kado, Y., Watanabe, O., Shiinoki, J., Iwazaki, K., Shigemura, H., Tsuji, H. and Matsumoto, S., 2020. Impacts of habitual diets intake on gut microbial counts in healthy Japanese adults. Nutrients 12: 2414. https://doi.org/10.3390/nu12082414
Takada, M., Nishida, K., Gondo, Y., Kikuchi-Hayakawa, H., Ishikawa, H., Suda, K., Kawai, M., Hoshi, R., Kuwano, Y., Miyazaki, K. and Rokutan, K., 2017. Beneficial effects of Lactobacillus casei strain Shirota on academic stress-induced sleep disturbance in healthy adults: a double-blind, randomised, placebo-controlled trial. Beneficial Microbes 8: 153-162. https://doi.org/10.3920/bm2016.0150
Toden, S., Anthony, B., David, T. and Michael, C., 2005. Resistant starch attenuates colonic DNA damage induced by higher dietary protein in rats. Nutrition Cancer 51: 45-51. https://doi.org/10.1207/s15327914nc5101_7
Toi, M., Hirota, S., Tomotaki, A., Sato, N., Hozumi, Y., Anan, K., Nagashima, T., Tokuda, Y., Masuda, N., Ohsumi, S., Ohno, S., Takahashi, M., Hayashi, H., Yamamoto, S. and Ohashi, Y., 2013. Probiotic beverage with soy isoflavone consumption for breast cancer prevention: a case-control study. Current Nutrition and Food Science 9: 194-200. https://doi.org/10.2174/15734013113099990001
Tsukahara, T., Matsukawa, N., Tomonaga, S., Inoue, R., Ushida, K. and Ochiai, K., 2014. High-sensitivity detection of short-chain fatty acids in porcine ileal, cecal, portal and abdominal blood by gas chromatography-mass spectrometry. Animal Science Journal 85: 494-498. https://doi.org/10.1111/asj.12188
Turnbaugh, P.J., Ley, R.E., Mahowald, M.A., Magrini, V., Mardis, E.R. and Gordon, J.I., 2006. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444: 1027-1031. https://doi.org/10.1038/nature05414
Verbeke, K.A., Boobis, A.R., Chiodini, A., Edwards, C.A., Franck, A., Kleerebezem, M., Nauta, A., Raes, J., Van Tol, E.A.F. and Tuohy, K.M., 2015. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutrition Research Reviews 28: 42-66. https://doi.org/10.1017%2FS0954422415000037
Wang, C., Nagata, S., Asahara, T., Yuki, N., Matsuda, K., Tsuji, H., Takahashi, T., Nomoto, K. and Yamashiro, Y., 2015. Intestinal microbiota profiles of healthy pre-school and school-age children and effects of probiotic supplementation. Annals of Nutrition and Metabolism 67: 257-266. https://doi.org/10.1159/000441066
Wang, J., Qin, J., Li, Y., Cai, Z., Li, S., Zhu, J., Zhang, F., Liang, S., Zhang, W., Guan, Y., Shen, D., Peng, Y., Zhang, D., Jie, Z., Wu, W., Qin, Y., Xue, W., Li, J., Han, L., Lu, D., Wu, P., Dai, Y., Sun, X., Li, Z., Tang, A., Zhong, S., Li, X., Chen, W., Xu, R., Wang, M., Feng, Q., Gong, M., Yu, J., Zhang, Y., Zhang, M., Hansen, T., Sanchez, G., Raes, J., Falony, G., Okuda, S., Almeida, M., Lechatelier, E., Renault, P., Pons, N., Batto, J.M., Zhang, Z., Chen, H., Yang, R., Zheng, W., Li, S., Yang, H., Ehrlich, S.D., Nielsen, R., Pedersen, O., Kristiansen, K. and Wang, J., 2012a. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 490: 55-60. https://doi.org/10.1038/nature11450
Wang, T., Cai, G., Qiu, Y., Fei, N., Zhang, M., Pang, X., Jia, W., Cai, S. and Zhao, L., 2012b. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME Journal 6: 320-329. https://doi.org/10.1038/ismej.2011.109
Yao, C.K., Muir, J.G. and Gibson, P.R., 2016. Review article: insights into colonic protein fermentation, its modulation and potential health implications. Alimentary Pharmacology and Therapeutics 43: 181-196. https://doi.org/10.1111/apt.13456


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}