Abstract
For animals living in social groups, cooperation is a key factor to success. It has been postulated that in social systems with cooperative breeding or a tolerant dominance style, individuals will benefit each other. Cooperation is, therefore, not expected in long-tailed macaques, since they do not breed cooperatively and experience a steep unidirectional dominance hierarchy. However, previous studies have shown that they can be prosocial in a dyadic setting. This would comply with the more recently postulated dyadic interdependence hypothesis. To be able to compare their cooperative performances with other species, we set up a group service paradigm similar to that which has been tested in a number of species. We presented three groups of socially housed long-tailed macaques with a swing set apparatus: a beam outside their enclosure with a rope attached in the middle and a reward location on one side. The provider pulled the rope in the middle to provide a reward to another individual, but could not reach the reward itself. The macaques showed prosocial behaviour in the test significantly more often than in two control conditions. They preferably provided to kin. The prosocial behaviour of the despotic, individual breeding long-tailed macaques counters the cooperative breeding and self-domestication hypotheses, yet supports the dyadic interdependence hypothesis, although future studies on other macaque species with more tolerant dominance styles should elucidate the effect of dominance styles on prosociality.
1. Introduction
Prosocial behaviour, acts from an individual that directly reward the receiver but do not benefit the provider (Massen et al., 2011; Burkart et al., 2018; Melis, 2018; Martin et al., 2021), is considered crucial for initiation and maintenance of cooperation. This is an important benefit of living in a social group (primates: Silk & House, 2011; Burkart et al., 2014; Callaghan & Corbit, 2018; humans: Hill et al., 2009; Antón et al., 2014). Originally, it was presumed that only humans could act prosocially (Maclean, 2016; Hare, 2017), however, a multitude of studies showed some form of prosociality in a wide range of primate species (reviewed in Cronin, 2012). Yet several studies on chimpanzees (Pan troglodites) and bonobos (P. paniscus) have failed to show prosocial tendencies, even though these two ape species are considered very cooperative (Silk et al., 2005; Vonk et al., 2008; Silk & House, 2011; Tennie et al., 2016, 2021; Verspeek et al., 2022), although other studies in chimpanzees did show prosocial tendencies (Horner et al., 2011; Claidière et al., 2015; van Leeuwen et al., 2021). Similarly, in a non-reciprocal prosocial task cotton-top tamarins (Saguinus oedipus) were not prosocial (Cronin et al., 2010), while common marmosets (Callitrix jacchus) were (Burkart et al., 2007). This between-species variation in prosocial behaviour has been linked to species differences in social systems, namely depending on cooperative breeding (Burkart et al., 2014) and self-domestication (Hare, 2017), i.e. egalitarian dominance hierarchies.
To understand the evolution of prosociality, comparative studies are required, however the multitude of different experimental paradigms, set-ups and procedures (reviewed in Cronin, 2012; Dale et al., 2016) make such comparisons difficult. In a first attempt to standardize the experimental paradigm and its procedures across a diverse group of species, Burkart and van Schaik designed a group service paradigm (Burkart & Van Schaik, 2013). In such an open setting with active partner choice, individuals can voluntarily provide food to their group members, while they are not able to reach for that food themselves, nor are they rewarded for doing so in any other way. In a multi-lab effort, they subsequently tested this paradigm in 15 different primate species, and found that prosocial behaviour correlates with extensive allomaternal care (Burkart et al., 2014). These findings supported the cooperative breeding hypothesis (Burkart et al., 2009), which suggests that helping others to rear their young requires high social tolerance and prosociality. Accordingly, empirical evidence for a link between prosociality and cooperation is also found in the cooperatively breeding common marmosets, where in an experimental set-up cooperative success increases twofold when there was at least one prosocial individual in the test dyad (Martin et al., 2021). Further support for this hypothesis comes from a recent avian study, i.e., a comparison of eight different corvid species, which also found the highest prosocial tendencies among the cooperatively breeding birds (Horn et al., 2020). The study on corvids, however, also showed that colonially nesting species have relatively high prosocial tendencies, and these authors suggest that heightened social tolerance (at the nest) may have been an evolutionary pathway to prosociality (Horn et al., 2020).
The evolutionary selection pressure towards the most friendly individuals leads to self-domestication (Hare et al., 2012), and prosociality may, therefore, be restricted to the more egalitarian primate species. This subsequently enhances the probability of prosocial behaviour, such as in Tonkean macaques (Macaca tonkeana; Scopa & Palagi, 2016; Joly et al., 2017). Interestingly, however, using a similar group service paradigm as the previous comparative studies (Burkart & Van Schaik, 2013; Burkart et al., 2014; Horn et al., 2020), Bhattacharjee and colleagues recently showed prosocial tendencies in Japanese macaques, Macaca fuscata. This species is considered very despotic (on a scale from I, despotic, to IV, egalitarian, they are classified as class I: Thierry, 2000, 2007), which makes it a less obvious candidate (Bhattacharjee et al., 2023). They argue that social tolerance at the dyadic level may also spark prosocial motivations, particularly when individuals rely on social bonds for alliances to cope with the complexities of their social organisation. This overarching interdependency hypothesis (Tomasello et al., 2012; Bhattacharjee et al., 2023), therefore, suggests that both interdependencies at the dyadic level (e.g. alliances; kinship) as on the group level (cooperative or colonial breeding) may have caused the evolution of prosociality in different taxa. They therefore call for more analyses on the dyadic level, as well as for more studies on species that are considered rather despotic (Bhattacharjee et al., 2023).
The open setting of the group service paradigm allows, due to testing this in a group setting, for more natural interactions between the participants. In a group setting, prosociality can be classified as proactive or reactive (review: Kopp et al., 2024). When the provider seems triggered by an external motivation, like an audience present at a close social distance (Jaeggi et al., 2010) a threat or social pressure (Horn et al., 2020), prosocial behaviour is considered reactive. Alternatively, prosocial behaviour is considered proactive when a solicitation by a group member is absent. This proactive prosociality is considered a prerequisite for human social development (Burkart & Van Schaik, 2013). Note that this classification is descriptive and does not concern the internal motivation of the prosocial individual.
Long-tailed macaques (Macaca fascicularis), while not as despotic as the Japanese macaques, are also categorized as a despotic species (class II: Thierry, 2007; Adams et al., 2015). Females are philopatric, whereas males disperse at the age of 3–4 years old. Infants inherit the rank under their mother due to their mother’s support, leading to nepotism (Van Noordwijk & Van Schaik, 1985; Van Schaik & Van Noordwijk, 1987). This means that they will favour kin and are less tolerant towards non-related group members. On top of this, these macaques do not help to raise other’s offspring, therefore any prosociality from these macaques would not conform to the cooperative breeding hypothesis (Burkart et al., 2014). More dominant individuals can monopolise food resources yet unlike brown capuchin monkeys do not share food, which is also seen as a prerequisite for prosocial behaviour (Brosnan, 2010). Nevertheless, a previous study indicated that these macaques do benefit group members in a dyadic behavioural experimental set-up, that dominant individuals are more prosocial than subordinate ones, and that they are most prosocial towards kin (Massen et al., 2012). Further tests showed that when the long-tailed macaques could choose whom to benefit in a triadic set-up, that they consider the relative dominance position of their partners more than the relationship quality between themselves and the potential receiver (Massen et al., 2011). In a forced choice-task, however, long-tailed macaques prefer to take a high value food reward by themselves, above providing prosocially (Sterck et al., 2015). This calls for exploring prosocial behaviour in this despotic species in a group setting.
The aim of this study was to test prosocial behaviour of long-tailed macaques with a group service paradigm (Burkart & Van Schaik, 2013; Burkart et al., 2014; Horn et al., 2020; Martin et al., 2021; Bhattacharjee et al., 2023). In line with the previous studies in this species, we expect dominant individuals to participate more often than subordinates. Moreover, we expect that younger animals will participate more often than adults, because they are more playful and explorative (Tan et al., 2018). Because females will stay in their natal group (Van Schaik et al., 1983), and have to cooperate more intensively with group members, we expect them to be more prosocial than males. Alternatively, males may work together because they form male-male coalitions and disperse together (Van Noordwijk & Van Schaik, 1985; Mishra et al., 2020) and males may provide to females, for example as a strategy to gain mating success (cf. friendships: Massen et al., 2012; Sterck et al, 2024). In addition, we expect that the individuals that act prosocially will provide food rewards more frequently to kin than non-kin (Massen et al., 2010a,b, 2011). Finally, we expect an individual to provide a group member when they have a positive relationship (a high relationship quality), or when the other has a higher rank in the dominance hierarchy (Massen et al., 2010a,b, 2011).
2. Methods
2.1. Subjects and housing
The observations and behavioural experiments took place at the Biomedical Primate Research Centre (BPRC) in Rijswijk, The Netherlands. Three groups of long-tailed macaques (group I,
The animals were used to observers and had access to lots of enrichments and climbing opportunities (Vernes & Louwerse, 2010). They were fed twice a day, in the morning with monkey chow, in the afternoon with fruit or vegetables, and water was available ad libitum. Engaging in the behavioural experiments was on a voluntary basis, without food or water deprivation. Permission for the research was granted by the institute’s Animal Welfare Body (IvD: Number IvD-019) and complies with Dutch and EU law.
2.2. Observational data collection
We observed the groups of long-tailed macaques to obtain information about the social relationships, using two different approaches: scan sampling (125 scans per group) and ad libitum observations (50 h per group). We made scans at least 30 min apart with a maximum of 8 per day and noted the exact location of all visible individuals, describing the proximity to individuals within one arm’s length. These scans were conducted in the inside enclosure on the same days as we did the behavioural experiments.
The relationship quality of a dyad was measured based on the mutual proximity in the scans, a valid measure to distinguish between friends and non-friends (Massen et al., 2010a,b). We summed up when two macaques were within one arm’s length of each other, divided by the total number of scans. When both individuals were not visible during a scan, we assumed that they were in proximity in the same ratio. The calculation is explained in more detail in Table S1 in the Supplementary materials. In addition, for analysis of individual characteristics, we calculated general sociality per individual as the average relationship quality, by averaging their dyadic Z-score transformed proximity (see below) with all other group members.
The dominance hierarchy was determined by scoring all submissive behaviours seen during encounters. Unidirectional behavioural elements indicating submission, i.e. silent bare teeth, make room and give ground (cf. Angst, 1974), were scored ad libitum (Martin & Bateson, 2018). For each group a dominance hierarchy was calculated using Matman (Schmid & De Vries, 2013), a package that was performed in R (version 4.1.3). The ensuing dominance rank is provided in Table 1, with number 1 being the highest-ranking individual per group. In both group II (linearity index

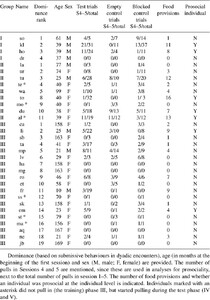
Characteristics of all individuals per group and their performance in test trials, empty trials and blocked control trials.
Citation: Behaviour 162, 15 (2025) ; 10.1163/1568539X-bja10338
Age and kinship were obtained from files of the BPRC colony management. To compare the age of individuals between different groups, we assigned age in months at the date the experiments started. Matrilineal relatedness of all group members was known and the assignment of kinship was based on matrilineal relatedness. Dyads with a relatedness of 0.5 (mother, brother, sister) or 0.25 (aunt, nephew, niece) according to the matrilines were considered kin, all other dyads were considered non-kin.
2.3. Experimental data collection
2.3.1. Test apparatus
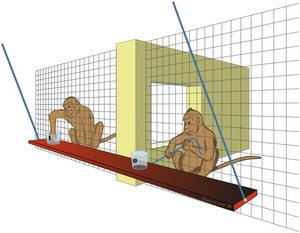
The group service paradigm (cf. Burkart & Van Schaik, 2013) was conducted using an adjusted apparatus, the ‘swing set’ (Figure 1). This swing set contained a beam of 1.2 m which hung from the ceiling by two steal chains. In the middle of the beam (position 0) and at the left-end side (position 1) a bucket was fastened to provide a small food reward. The swing set was attached in front of the enclosure, at a distance of 0.5 m approximately, out of reach for the macaques inside. The only way they could reach for the swing set was to pull a rope, attached to the middle of the beam at position 0 and attached with a carbine hook to the fence. The swing set was positioned in such a way, that a macaque at position 0 could not reach for the bucket at position 1. Inside the enclosure a wall obstructed direct access; the monkey who pulled had to release the rope to walk through an opening in the wall to be in position 1. Gravity consequently drew the beam out of reach for the macaque that pulled the rope. We also attached a rope at the human observer side of the swing set, to be able to control the swing set and to prevent it from swinging back and forth.
Swing set apparatus, the macaque (the provider) in position 0 is providing a food reward to another individual (the receiver) in position 1. The macaques can see and interact with each other through an opening in the wall. The macaque in position 0 cannot reach for the bucket in position 1, because it has to let go of the rope.
Citation: Behaviour 162, 15 (2025) ; 10.1163/1568539X-bja10338
2.3.2. Experimental design
The experiment consists of five different phases (cf. Burkart & Van Schaik, 2013): a habituation phase (I); a phase to test for social tolerance (II); followed by a training phase (III); and a test phase (IV). After the regular tests and controls, we conducted a blocked control phase (V). Every session was video recorded with two cameras. We tested two times a week, in an indoor setting, under comparable circumstances in all three groups.
2.3.2.1. Phase I: Habituation.
The beam was directly attached to the outside of the enclosure. All long-tailed macaques were able to reach into the bucket at position 0 to take a food reward (one piece of maize grain, pea or sunflower seed), during a session which lasted 90 min. Once an animal obtained 10 rewards in a single habituation session, we waited for that individual to leave and did not bait the apparatus again. When this individual was out of sight, we provided a new reward to attract other macaques. This was done to avoid monopolisation and to achieve a maximum number of individuals that participated. Our aim was to habituate as many animals as possible to the swing set.
When every individual that took a reward passed the criterium of 10 successful attempts per session, we ended the habituation phase and continued to the next phase. In group I 3 out of 4 macaques succeeded, in group II 6 out of 9 and in group III 12 out of 19 macaques passed the criterium.
2.3.2.2. Phase II: Social tolerance test.
In two separate sessions, on two different days, we tested the accessibility of the attached swing set for each individual in the different groups. While the beam was attached to the fence (cf. phase I), we baited the middle bucket (position 0) every 2 min during 5n trials (
2.3.2.3. Phase III: Training.
During the training phase, the beam could move like a swing, hanging from the ceiling outside of the enclosure, with a rope attached to the fence. If a monkey pulled the rope, the beam came forth and the monkey was able to take a reward out of the bucket in position 0. Our criterium to consider the training completed, was 10 pulls where the macaque obtained a reward in a session. Again, if an individual pulled more than 10 times during one session, we waited for that individual to leave to avoid monopolisation and to achieve a maximum of individuals that participated (similar to phase I). The next phase started when no new macaques participated voluntarily any further. To ensure that the animals understood the contingencies of the test apparatus, we performed sufficient training sessions in the different groups (Group I: 76 trials in 2 sessions; Group II: 177 trials in 4 sessions; and Group III: 130 trials in 3 sessions). The number of animals that pulled the rope at least 10 times in at least one session was in group I: 4/4 (100%), in group II: 5/9 (55%) and in group III: 13/19 (68%). In total 22 macaques passed our training criteria, which corresponds to 69% of all long-tailed macaques in this study.
2.3.2.4. Phase IV: Group service test.
In this phase, during the regular test trials, we baited only the bucket at position 1 with a food reward (maize grain, pea or sunflower seed) and registered which individual pulled the rope in position 0 to move the swing set towards the fence. We also noted the individuals that were present or approached bucket 1 and whether they successfully obtained the food reward. To keep the participants motivated, we provided bucket 0 with a food reward in motivational trials. Every session started with a motivational trial, followed by five regular test trials. These six trials were repeated in each session as many times as we had participating macaques (n) in the habituation phase in that group. We ended every session with a motivational trial. The number of trials per session for each group was calculated. Each trial started after a position was baited (or pseudo-baited) and lasted 2 min. In 217 out of 356 trials where a provider pulled (61%) one reward was present. In 159 trials receiver took one reward. When in a test trail food was not taken from position 1, we added a reward in the next trail. This happened 5 times with more than 10 rewards (see Table S2 in the Supplementary materials for an overview of all test sessions) and the maximum number of accumulated rewards was 12. In 37% of all trials, the receiver took only one reward.
In group I the number of participating macaques
If three consecutive motivational trials were neglected, we would end the session. This occurred once in session 5 in Group III, because many individuals were attracted to something outdoors. We stopped this session after 31 trials and started all over again a day later. This incomplete session was ignored in our analyses and we treated the next, completed, session as session number 5. To be sure that the macaques understood the contingencies of the task; i.e. if they would keep pulling without rewarding themselves, we executed five sessions in total per test and control conditions (cf. Burkart & Van Schaik, 2013; Bhattacharjee et al., 2023). In the first three sessions, the results may be affected by learning attempts. Therefore, following previous studies (cf. Burkart et al., 2014; Horn et al., 2016, 2020; Bhattacharjee et al., 2023), in the analyses we only used the results from the last two sessions of these test trials and of the controls (see below).
To ensure the monkeys understood the consequence of their behaviour, two types of controls were done: an empty control in phase IV and a blocked control in phase V. We alternated the regular test-sessions in Phase IV with empty control sessions, where we conducted the exact same experiment without a reward in position 1 in the empty control trials. By pretending to bait bucket 1, we drew attention to each trial and scored the responses of the macaques when there was no reward in bucket 1. In the empty control trials we also counted the pulls by the participating individuals, no food provisions were possible, because rewards were absent. These empty control sessions consisted of the same number of trials, and, similar to the test trials, in the intermitted (once every 6 trials) motivational trials we baited bucket 0 to keep the macaques interested. In our calculations we compared the test results with the empty control results. The other type of control was conducted in Phase V.
The food provisions in regular test trials resulted in providing another individual, the receiver, with a food reward. For descriptive purposes, we made a distinction between reactive prosociality, when the provider pulled in position 0 and a receiver was present near position 1, and proactive prosociality (Kopp et al., 2024), when the provider started to pull when no receiver was present, yet allowed another group member to take a reward.
2.3.2.5. Phase V: Blocked control.
The second control experiment was a blocked control situation where a transparent screen, attached to the fence in front of position 1, prevented a receiver from taking the reward. The aim of this set up was to check if the provider would pull anyway, even if the screen made it impossible for the receivers to reach for the reward. We included both baited blocked control and empty blocked sessions to make both phases comparable (cf. Burkart & Van Schaik, 2013; Bhattacharjee et al., 2023). These were interspaced with motivational trials with the same time schedule as in Phase IV. We counted per session for each individual the number of pulls in the baited blocked control trials with a reward in bucket 1, further called blocked control trials. Note that within one trial, an individual could pull more often in the blocked control condition. In addition, due to COVID-19 regulations we could not remove the rewards in the blocked control and these were accumulating in the unreachable position 1.
In the first sessions of phase VI and V, we saw some attempts by specific individuals to reach for the reward in bucket 1 by themselves. To obstruct these events, we pulled the swing set back (out of reach for the macaques inside the enclosure) with a rope attached to our side of the beam. However, when a macaque succeeded in its attempt to take the food reward in bucket 1, we omitted this trial from the dataset, i.e., we did not count this as a normal pull.
2.4. Data analysis
We used R version 4.4.0 for statistical analysis of the data and to produce all graphs.
To determine tolerance in the social tolerance test—phase II—the evenness scores resulting in a Pielou’s J (Pielou, 1966) were determined for each test day and averaged. To measure the rate of participation of each individual, we counted the number of rewards they took. Pielou’s J ranges from 1.0, every macaque gets the same number of food rewards, to 0, one individual obtains all food rewards.
We compared the number of pulls, irrespective of whether a receiver obtained the reward, during the regular test trials in the test sessions versus the empty control sessions of phase IV and the blocked control sessions of phase V to determine prosocial behaviour. We performed a Friedmans test to explore differences in number of pulls, and post-hoc a Wilcoxon test to distinguish between the conditions. For this comparison only sessions 4 and 5 were taken into account, to avoid an effect of learning (cf., Burkart et al., 2014; Horn et al., 2016, 2020; Bhattacharjee et al., 2023). Similarly, we compared the number of pulls in the final two sessions of the test and empty and blocked controls per individual, using chi-squared tests, to determine on an individual level whether an individual could be considered prosocial. Note here that the number of pulls each individual could perform in the (test) sessions was restricted by the number of pulls of all other participating individuals.
To analyse the characteristics of prosocial individuals, we considered all individuals that pulled significantly more in the test sessions than in at least one of the controls as prosocial individuals (cf. Horn et al., 2016, 2020; Bhattacharjee et al., 2023; Table 1). To analyse what characteristics, be it age, rank, sex, or general sociality, had an effect on the likelihood of an individual to be prosocial, we ran a binomial GLM.
The combination of provider and receiver for food provisions is a key to unravel which characteristics determine a successful delivery of the reward. However, in many possible dyads we observed no prosociality, which made our data heavily zero-inflated. Our solution to this was fitting a two step analysis to investigate (1) which dyadic factors predicted the likelihood of food provisions within a dyad (binomial GLMM) and (2) which dyadic factors predicted the total number of food provisions within dyads in which at least one food provision had been observed (Poisson GLM). The first analysis concerns the likelihood of food provisions within a dyad, further called the presence of food provisions; the second analysis concerns the number of food provisions observed within a dyad, further called number of food provisions.
To describe each dyad, we determined these characteristics: same sex (provider: male and receiver: male or both female) or different sex (provider: male and receiver: female or provider: female and receiver: male), age difference (age of provider in months minus age of receiver in months) and maternal kin or non-kin. Other factors we took into account were rank-difference and relationship quality. To standardise the rank-difference (rank provider minus rank receiver) and relationship quality in all groups, we z-transformed these data.
For the first model, we fitted a GLMM with a binomial error distribution using the glmmTMB package (Brooks et al., 2017), with the presence of food provisions within a dyad (yes/no) as dependent variable (
In the second model, we used a GLM with a Poisson error distribution with the total number of food provisions within each dyad as the response variable, regarding only the provisioning dyads (
3. Results
3.1. Social tolerance test
Because of their despotic characterisation (Thierry et al., 2004), we predicted a great inequity in access to resources between the individuals of each group. However, in the Tolerance Test (phase II) in group I, 3 out of 4 macaques took a food reward out of bucket 0, which was considered a successful attempt. In group II 5 macaques out of a total of 9 participated successfully, and in group III we had 12 macaques from a group of 19 individuals, who took at least one reward from bucket 0. The calculated evenness score Pielou’s J on average was 0,63 (Group I:
3.2. Group service test, group level
To test for prosocial preferences across our test animals, we compared the number of pulls in regular test trials during test sessions with the number of pulls during the empty control sessions and the blocked control sessions. When the provider acts in order to provide food to another individual, he or she would pull less often when food is absent in location 1 (in an empty control trial), or the food reward in location 1 is not accessible (in a blocked control trial). During these test and control trials, three additional animals in group II and three in group III started participating (Table 1, indicated with an asterisk), this renders our final sample size to 27.
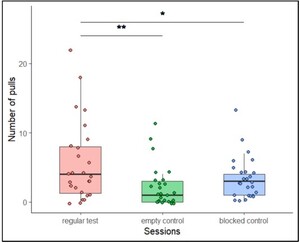
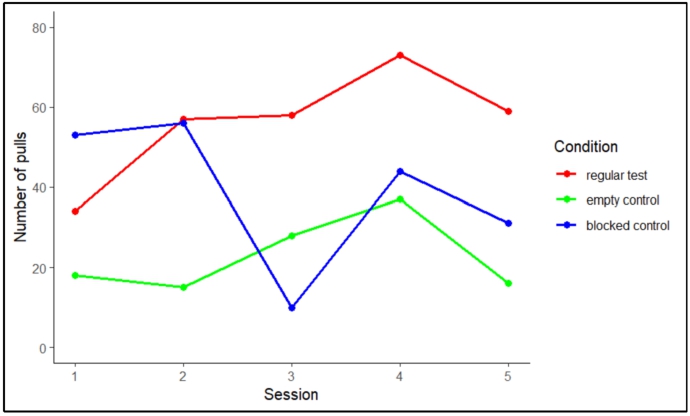
The number of pulls in the regular test sessions increased and remained high, compared to the total number of pulls in both control sessions (Figure 2). The number of pulls in both control sessions seemed to decrease over time. Pulling in the motivational trials remained high in both test and control trials.
The total number of pulls in each of the five sessions in the regular test trials (phase IV), the empty control trials (phase IV) and the blocked control trials (phase V).
Citation: Behaviour 162, 15 (2025) ; 10.1163/1568539X-bja10338
To ensure that in these trials all test animals understood the implications of both regular test and control sessions, the combined pulling of each individual in Sessions 4 and 5 of the test and control was compared. We found an overall significant difference in the number of pulls per condition (Friedman’s test:
The number of pulls in the last two sessions (4 and 5) in the regular test condition (phase IV), the empty control condition (phase IV) and the blocked control condition (phase V).
Citation: Behaviour 162, 15 (2025) ; 10.1163/1568539X-bja10338
3.3. Group service test, individual level
We explored whether individuals with specific characteristics (sex, age, dominance rank and average relationship quality) differed in how often they provided rewards. To this end, we first determined for each providing individual whether it was prosocial or not in Sessions 4 and 5 (Table S5 in the Supplementary materials). One individual, Horsten, pulled significantly more often in regular test sessions compared to both the empty and blocked control sessions (Test vs Empty control:
3.4. Group service test, food provisioning
On average, the long-tailed macaques in the three tested groups pulled in 68% of the regular test trials (group I: 60 out of 100 trials; group II: 124 out of 150 trials; group III: 100 out of 165 trials). However, rewards were not always taken by group members. If we consider trials in which there was a food provision, on average only 28% of the regular test trials led to a food provisioning (group I: 20 out of 100 trials; group II: 57 out of 150 trials; group III: 41 out of 165 trials). Of these 118 food provisions, 58 were proactive and 60 were reactive (reactive food provisions: in group I: 11 out of 20 trials; group II: 21 out of 57 trials, group III: 28 out of 41 trials). If we then only consider those seven prosocial individuals that showed sufficient understanding of the task (cf. Burkart et al., 2013, 2014; Horn et al., 2016, 2020; Bhattacharjee et al., 2023), they are responsible for 58% (68 out of 118 trials) of the food provisions. When a receiver took a reward in position 1 that was provided by another macaque who pulled in position 0, it could potentially lead to some aggression by the provider towards the receiver. In all 118 pulls with a receiver, however, only 7 were followed by a threat from the provider. In 94% of the food provisions the provider allowed a receiver to take the reward. We never observed aggressive behaviour from the receiver to the provider during a trial.
3.5. Dyadic predictors of food provisions
To investigate the dyadic predictors of food provisions, we performed an analysis in two steps. As a first step, we investigated the presence of food provisions within a dyad for each potential dyad using a binomial GLMM (see Table S8 in the Supplementary materials for the dyadic characteristics; Table S9 for the results of the GLMM). We found no significant interaction between provider age and receiver age (
The dyadic presence of food provisions and the dyadic number of food provisions in non-kin or kin dyads.
Citation: Behaviour 162, 15 (2025) ; 10.1163/1568539X-bja10338
The second step, to investigate which variables predicted the number of food provisions within a dyad that had at least provided once, we used a Poisson GLM. We again found no significant interaction between provider sex and receiver sex (
4. Discussion
Prosocial behaviour in a group service paradigm is expected in cooperatively breeding or self-domesticated primates (Jaeggi et al., 2010; Burkart & Van Schaik, 2013). Nevertheless, despotic macaques have also been shown to be prosocial in other prosociality tasks (long-tailed macaques: Massen et al., 2010a,b, 2012) or in a group service paradigm (Japanese macaques: Bhattacharjee et al., 2023), possibly due to interdependency, i.e. when cooperators can benefit as a secondary consequence of helping their recipients (Roberts, 2005). Here we present evidence that long-tailed macaques, a species that does not breed cooperatively and is not egalitarian, showed relatively high tolerance at group level and acted prosocially in a group service task. Prosocial behaviour did not depend on individual characteristics of age, sex or dominance rank, yet food provisions occurred more often among kin than non-kin dyads. Even in their unidirectional hierarchic organised groups, long-tailed macaques were rarely aggressive towards the beneficiary, when it took a food reward. Altogether, this supports earlier findings that prosociality is present in this despotic macaque species.
4.1. Social tolerance
The first notable outcome of our study is the relatively high level of tolerance the long-tailed macaques exhibited. Many individuals participated in the tolerance tests and in all three tested groups we found a relatively high evenness score that is comparable to the score of the more egalitarian and food sharing brown capuchin monkeys (
Still, the long-tailed macaque dominance hierarchy typically explains the highly unidirectional type of aggression which can lead to differential resource access (Overduin-de Vries et al., 2020). In addition, long-tailed macaques have a unidirectional submission signal (i.e. the bared teeth display: Angst, 1974). Thus, the dominance relationships seem as obvious as in class I macaques. However, long-tailed macaques are considered class II due to their (slightly) higher rate of reconciliation, that may be possible due to relatively high tolerance for the opponent after a conflict (cf. Kempes et al., 2009). While aggression may result in access to resources, dominance position in itself does not (Overduin-de Vries et al., 2020), suggesting that less aggressive individuals may be more tolerant. Accordingly, in our study beneficiaries of the food rewards rarely received aggression from the provider (in only 7 out of 116 cases). Whether this difference in tolerance indeed distinguishes between despotic macaques of class I and class II, and of course is even higher in class III and VI (cf. Thierry et al., 2000), remains to be further explored.
4.2. Prosocial behaviour
The long-tailed macaques were prosocial in a group setting, both at individual and group level. They performed better in the test trials than in both control trials. This is congruent with earlier findings of prosociality in a dyadic or triadic setting in this species (Massen et al., 2010a,b, 2011). In addition, this was found in each of the three groups, despite their different composition and size. This suggests that this is a general feature of this species and not a result of one outlier group. However, animals of this species were not prosocial in a dyadic setting when they had to forego a reward and choose the option without food to just provide someone else (Sterck et al., 2015). Yet, the results in the current group setting indicate that they can be prosocial in a less demanding setting where they receive no reward for providing another individual too. Moreover, the long-tailed macaques that participated in the test phase seemed to understand the contingencies of the task with the group service paradigm, also in our alternative design with a swing set apparatus, because in sessions 4 and 5 they pulled significantly more often when a food reward was available to a partner, compared to the phases when there was no food reward available (empty control) or when access was prevented (blocked control). Pulls in the motivational trials remained high both during test and control sessions. Because of the similar set up and occurrence of the motivational trials, the difference in pulls between test and controls is not caused by the availability of a reward in the motivational trials during the regular test and control trials.
Our findings indicate that despotic long-tailed macaques, similar to Japanese macaques (Bhattacharjee et al., 2023), can be prosocial. This contradicts both the cooperative breeding and self-domesticated species hypotheses, but may support the alternative and overarching interdependency hypothesis (Tomasello et al., 2012; Bhattacharjee et al., 2023). This hypothesis argues that interdependency at either group or dyadic level or at both levels may lead to prosocial behaviour. While cooperative breeders may be dependent at the group level, in other group-living species interdependency may be found at the dyadic level, for example among kin or individuals with good relationships.
4.3. Individual characteristics of prosocial individuals
Out of 27 participating individuals, 7 were significantly prosocial at the individual level. This is similar to the relative number of prosocial individuals in the despotic Japanese macaque (9 out of 25 individuals: Bhattacharjee et al., 2023). This is a conservative estimate of the proportion of prosocial individuals, since these prosocial individuals pulled relatively often and may have prevented other individuals from pulling and showing their prosocial behaviour at the individual level. In our experiment, the individual characteristics age, sex, dominance rank and general sociality did not affect whether an individual was prosocial. This contrasts with the earlier finding that dominants were more prosocial in a dyadic setting (Massen et al., 2010a,b). However, in a triadic setting, subordinates were more likely to pull for dominant individuals (Massen et al., 2011). In dyadic and triadic settings, prosocial behaviour always has a specific receiver, similar to the reactive setting in a group service paradigm. This suggests that the set-up of the experiment, i.e. dyadic, triadic or a group setting, may determine the importance of dominance rank in relation to prosocial behaviour.
4.4. Dyadic characteristics of prosocial behaviour
The interdependency hypothesis (Tomasello et al., 2012; Bhattacharjee et al., 2023) argues that interdependence at the dyadic level may promote prosocial behaviour in a despotic species like the long-tailed macaque. Indeed, when analysing behaviour during food provisions i.e. when someone obtained the food, we found that the presence of food provisions was more likely in a dyad with kin than with non-kin. This preference to provide food for kin is similar to long-tailed macaque behaviour in a dyadic paradigm (kin: Massen et al., 2010a,b) and triadic paradigm (friends: Massen et al., 2011). However, in long-tailed macaques in dyads with prosocial behaviour, providers showed a lower number of food provisions for kin than for non-kin. This pattern contrasts with the pattern in Japanese macaques in a group service paradigm, where dyadic tolerance predicted the likelihood (presence) of food provisions and kinship predicted the number of food provisions (Bhattacharjee et al., 2023). Nevertheless, this outcome is consistent with the prediction of the interdependency hypothesis (Tomasello et al., 2012; Bhattacharjee et al., 2023) that food provisions are more likely found among dyadically interdependent individuals.
That the macaques showed a lower number of food provisions in prosocial dyads for kin than for non-kin individuals was an unexpected outcome that may be explained in several ways. First, subjects may need to be more prosocial to non-kin than to kin. To maintain a good relationship with kin may require less investment than a good relationship with non-kin. However, this was not borne out by our analyses, as we do not find an effect of relationship quality on the number of food provisions. Second, some individuals may be more eager to obtain food and be more often close to the provisioning location. This may, for example, concern individuals with a bold or persistent personality. Such persistent or bold individuals may obtain relatively large numbers of rewards, irrespective of who was provisioning. Unfortunately, we do not have data on personality. While this may explain the number of food provisions obtained, it does not explain that subjects were more likely to provision for kin. This line of reasoning suggests that the presence of food provisions was not a by-product of personality. Third, receivers may be relatively dominant, keeping other individuals away from the provisioned location. Since individuals have more non-kin in the group than kin, when providing is not directed at a specific individual, they will more often obtain food from non-kin than kin. A post-hoc analysis of the characteristics of the receivers (Table S10 in the Supplementary materials) showed more dominant individuals more often obtained a reward (Est.
Another descriptive finding was that about half of the food provisions were proactive, without a receiver present, and half were reactive, i.e. when a receiver was present in position 1. Also despotic Japanese macaques showed both types of provisioning (22% of the food provisions were reactive and 78% proactive: Bhattacharjee et al., 2023). Further exploring these two types of food provisioning may reveal whether reactive food provisions are related to provisioning specific individuals, while proactive food provisions may depend on group characteristics (Jaeggi et al., 2010) or are conducted by specific (e.g., bold, persistent, dominant) individuals. Unfortunately, for such analyses our data-set had to be split and these reduced data-sets rendered not enough statistical power to perform these analyses.
4.5. Strengths and weaknesses
To compare prosocial behaviour in long-tailed macaques with other species, we conducted a group service paradigm (cf. Burkart & Van Schaik, 2013; Bhattacharjee et al., 2023) in three different groups. The large number of participating individuals (
Due to COVID-19 regulations at the BPRC, we were not allowed to come close to the enclosures (<1 m), therefore we baited the swing set apparatus with a stick. In the blocked phase trials we could not remove the rewards, which results in a piling of food items. This could have an exaggerated impulse for some macaques to keep trying to obtain the accumulating price, which would yield results contra to our predictions. However, despite this caveat and the increasing number of rewards, the number of pulls was higher in the regular test trails than in the blocked control trials.
We did not perform a new pair of test sessions after the blocked phases (cf. Horn et al., 2016; Bhattacharjee et al., 2023) to test the monkey’s understanding of the swing set apparatus. This was due to restrictions by the BPRC during the COVID-19 pandemic. Nevertheless, our results show an increase in the number of pulls during regular test sessions and a reduction in both types of control sessions, indicating that the long-tailed macaques understood the task.
5. Conclusion
Prosocial behaviour in long-tailed macaques results in providing food rewards in a group service paradigm, although this is a despotic species with independent breeding that does not share food in a naturalistic environment. Consistent with the interdependency hypothesis, prosocial behaviour was more often directed towards kin, yet unexpectedly the monkeys provided food less often for kin than non-kin. However, receivers were relatively often dominants, possibly preventing kin of the provider from receiving food. About half of the food provisions were reactive, when a receiver was already present, the other half were proactive, when no-one was yet present. Thus, long-tailed macaques showed prosocial behaviour both for specific individuals (reactive) and to the group in general (proactive). Altogether, the prosocial behaviour of the despotic, individual breeding long-tailed macaques supports the dyadic interdependence hypothesis. Future studies on other macaque species with more tolerant dominance styles should elucidate the effect of dominance styles on prosociality.
Corresponding author’s e-mail address: e.j.a.m.delaat@uu.nl
Acknowledgements
J.J.M. Massen and E.H.M. Sterck share last authorship. We thank Debottam Bhattacharjee for providing additional information on group I and Dian Zijlmans for her assistance during data collection. Besides these colleagues, we thank caretakers, veterinarians and staff of the BPRC for their dedication and professionalism which resulted in optimal housing conditions for the macaques. This publication is part of the project Teachers Grant with file number 023.013.052 of the research programme “Role models for juvenile long-tailed macaques: do they learn from experienced adults or from peers”, partly financed by the Dutch Research Council (NWO).
References
Adams, M.J., Majolo, B., Ostner, J., Schülke, O., De Marco, A., Thierry, B., Engelhardt, A., Widdig, A., Melissa, S.G. & Weiss, A. (2015). Personality structure and social style in macaques. — J. Pers. Soc. Psychol. 109: 338-353. DOI:10.1037/pspp0000041.
Angst, W. (1974). Das Ausdrucksverhalten des Javaneraffen Macaca fascicularis Raffles. — Adv. Ethol. 15: 1-90.
Antón, S.C., Potts, R. & Aiello, L.C. (2014). Evolution of early Homo: an integrated biological perspective. — Science 345: 1236828.
Appleby, M.C. (1983). The probability of linearity in hierarchies. — Anim. Behav. 31: 600-608. DOI:10.1016/S0003-3472(83)80084-0.
Bhattacharjee, D., Cousin, E., Pflüger, L.S. & Massen, J.J. (2023). Prosociality in a despotic society. — Iscience 26: 106587.
Brooks, B.G.J., Lee, D.C., Pomara, L.Y., Hargrove, W.W. & Desai, A.R. (2017). Quantifying seasonal patterns in disparate environmental variables using the PolarMetrics R package. — In: 2017 IEEE international conference on data mining workshops (ICDMW). IEEE, New York, NY, p. 296-302.
Brosnan, S.F. (2010). What do capuchin monkeys tell us about cooperation? — In: For the greater good of all: Perspectives on individualism, society, and leadership (Forsyth, D.R. & Hoyt, C.L., eds). Palgrave Macmillan, New York, NY, p. 11-27.
Burkart, J.M. & Van Schaik, C. (2013). Group service in macaques (Macaca fuscata), capuchins (Cebus apella) and marmosets (Callithrix jacchus): a comparative approach to identifying proactive prosocial motivations. — J. Comp. Psychol. 127: 212-225. DOI:10.1037/a0026392.
Burkart, J.M., Fehr, E., Efferson, C. & van Schaik, C.P. (2007). Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. — Proc. Natl. Acad. Sci. USA 104: 19762-19766.
Burkart, J.M., Hrdy, S.B. & Van Schaik, C.P. (2009). Cooperative breeding and human cognitive evolution. — Evol. Anthropol. 18: 175-186. DOI:10.1002/evan.20222.
Burkart, J.M., Allon, O., Amici, F., Fichtel, C., Finkenwirth, C., Heschl, A., Huber, J., Isler, K., Kosonen, Z.K., Martins, E., Meulman, E.J., Richiger, R., Reuth, K., Spillmann, B., Wiesendanger, S. & Van Schaik, C.P. (2014). The evolutionary origin of human hyper-cooperation. — Nature Commun. 5: 5747. DOI:10.1038/ncomms5747.
Burkart, J.M., Guerreiro Martins, E., Miss, F. & Zürcher, Y. (2018). From sharing food to sharing information. — Interact. Stud. Soc. Behav. Commun. Biol. Artif. Syst. 19: 136-150. DOI:10.1075/is.17026.bur.
Callaghan, T. & Corbit, J. (2018). Early prosocial development across cultures. — Curr. Opin. Psychol. 20: 102-106. DOI:10.1016/j.copsyc.2017.07.039.
Claidiere, N., Whiten, A., Mareno, M.C., Messer, E.J., Brosnan, S.F., Hopper, L.M., Lambeth, S.P., Schapiro, S.J. & McGuigan, N. (2015). Selective and contagious prosocial resource donation in capuchin monkeys, chimpanzees and humans. — Sci. Rep. 5: 7631.
Cronin, K.A. (2012). Prosocial behaviour in animals: the influence of social relationships, communication and rewards. — Anim. Behav. 84: 1085-1093. DOI:10.1016/j.anbehav.2012.08.009.
Cronin, K.A., Schroeder, K.K. & Snowdon, C.T. (2010). Prosocial behaviour emerges independent of reciprocity in cottontop tamarins. — Proc. Roy. Soc. Lond. B: Biol. Sci. 277: 3845-3851.
Dale, R., Quervel-Chaumette, M., Huber, L., Range, F. & Marshall-Pescini, S. (2016). Task differences and prosociality; investigating pet dogs’ prosocial preferences in a token choice paradigm. — PLoS ONE 11: e0167750. DOI:10.1371/journal.pone.0167750.
Engqvist, L. (2005). The mistreatment of covariate interaction terms in linear model analyses of behavioural and evolutionary ecology studies. — Anim. Behav. 70: 967-971.
Hare, B. (2017). Survival of the friendliest: Homo sapiens evolved via selection for prosociality. — Annu. Rev. Psychol. 68: 155-186. DOI:10.1146/annurev-psych-010416-044201.
Hare, B., Wobber, V. & Wrangham, R. (2012). The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression. — Anim. Behav. 83: 573-585. DOI:10.1016/j.anbehav.2011.12.007.
Hartig, F. (2022). DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.4.6. — R Foundation for Statistical Computing, Vienna.
Hill, K., Barton, M. & Hurtado, A.M. (2009). The emergence of human uniqueness: characters underlying behavioral modernity. — Evol. Anthropol. 18: 187-200.
Horn, L., Scheer, C., Bugnyar, T. & Massen, J.J. (2016). Proactive prosociality in a cooperatively breeding corvid, the azure-winged magpie (Cyanopica cyana). — Biol. Lett. 12: 20160649.
Horn, L., Bugnyar, T., Griesser, M., Hengl, M., Izawa, E.I., Oortwijn, T., Rössler, C., Schiestl, M., Suyama, M., Taylor, A.H., Vanhooland, L., Von Bayern, A.M.P., Zürcher, Y. & Massen, J.J.M. (2020). Sex-specific effects of cooperative breeding and colonial nesting on prosociality in corvids. — ELife 9: 235-244. DOI:10.7554/eLife.58139.
Horner, V., Carter, J.D., Suchak, M. & De Waal, F.B. (2011). Spontaneous prosocial choice by chimpanzees. — Proc. Natl. Acad. Sci. USA 108: 13847-13851.
Jaeggi, A.V., Burkart, J.M. & Van Schaik, C.P. (2010). On the psychology of cooperation in humans and other primates: combining the natural history and experimental evidence of prosociality. — Philos. Trans. Roy. Soc. B: Biol. Sci. 365: 2723-2735. DOI:10.1098/rstb.2010.0118.
Joly, M., Micheletta, J., De Marco, A., Langermans, J., Sterck, E.H. & Waller, B. (2017). Comparing physical and social cognitive skills in macaque species with different degrees of social tolerance. — Proc. Roy. Soc. Lond. B: Biol. Sci. 284: 20162738. DOI:10.1098/rspb.2016.2738.
Kempes, M.M., Den Heijer, E., Korteweg, L., Louwerse, A.L. & Sterck, E.H.M. (2009). Socially deprived rhesus macaques fail to reconcile: do they not attempt or not accept reconciliation? — Anim. Behav. 78: 271-277.
Kopp, K.S., Kanngiesser, P., Brügger, R.K., Daum, M.M., Gampe, A., Köster, M., Van Schaik, C.P., Liebal, K. & Burkart, J.M. (2024). The proximate regulation of prosocial behaviour: towards a conceptual framework for comparative research. — Anim. Cogn. 27: 5. DOI:10.1007/s10071-024-01846-w.
Maclean, E.L. (2016). Unraveling the evolution of uniquely human cognition. — Proc. Natl. Acad. Sci. USA 113: 6348-6354. DOI:10.1073/pnas.1521270113.
Martin, J.S., Koski, S.E., Bugnyar, T., Jaeggi, A.V. & Massen, J.J.M. (2021). Prosociality, social tolerance and partner choice facilitate mutually beneficial cooperation in common marmosets, Callithrix jacchus. — Anim. Behav. 173: 115-136. DOI:10.1016/j.anbehav.2020.12.016.
Martin, P. & Bateson, P. (2018). Measuring behaviour. — Cambridge University Press, Cambridge.
Massen, J.J.M., Sterck, E.H.M. & De Vos, H. (2010a). Close social associations in animals and humans: functions and mechanisms of friendship. — Behaviour 147: 1379-1412. DOI:10.1163/000579510X528224.
Massen, J.J.M., van den Berg, L.M., Spruijt, B.M. & Sterck, E.H.M. (2010b). Generous leaders and selfish underdogs: pro-sociality in despotic macaques. — PLoS ONE 5: e9734. DOI:10.1371/journal.pone.0009734.
Massen, J.J.M., Luyten, I.J.A.F., Spruijt, B.M. & Sterck, E.H.M. (2011). Benefiting friends or dominants: prosocial choices mainly depend on rank position in long-tailed macaques (Macaca fascicularis). — Primates 52: 237-247. DOI:10.1007/s10329-011-0244-8.
Massen, J.J.M., Van Den Berg, L.M., Spruijt, B.M. & Sterck, E.H.M. (2012). Inequity aversion in relation to effort and relationship quality in long-tailed macaques (Macaca fascicularis). — Am. J. Primatol. 74: 145-156. DOI:10.1002/ajp.21014.
Melis, A.P. (2018). The evolutionary roots of prosociality: the case of instrumental helping. — Curr. Opin. Psychol. 20: 82-86. DOI:10.1016/j.copsyc.2017.08.019.
Mishra, P.S., Pal, A., Velankar, A.D., Kumara, H.N. & Singh, M. (2020). Do males bond? A study of male-male relationships in Nicobar long-tailed macaques Macaca fascicularis umbrosus. — J. Biosci. 45: 1-8. DOI:10.1007/s12038-020-9995-y.
Overduin-de Vries, A.M., De Vries, H., Vermande, M.M., Reijntjes, A.H.A. & Sterck, E.H.M. (2020). Both aggressive and affiliative behaviour facilitate resource access in high-ranking female long-tailed macaques (Macaca fascicularis). — Behaviour 157: 267-287. DOI:10.1163/1568539X-bja10001.
Pielou, E.C. (1966). The measurement of diversity in different types of biological collections. — J. Theor. Biol. 13: 131-144. DOI:10.1016/0022-5193(66)90013-0.
Roberts, G. (2005). Cooperation through interdependence. — Anim. Behav. 70: 901-908. DOI:10.1016/j.anbehav.2005.02.006.
Schmid, V.S. & De Vries, H. (2013). Finding a dominance order most consistent with a linear hierarchy: an improved algorithm for the I&SI method. — Anim. Behav. 86: 1097-1105. DOI:10.1016/j.anbehav.2013.08.019.
Scopa, C. & Palagi, E. (2016). Mimic me while playing! social tolerance and rapid facial mimicry in macaques (Macaca tonkeana and Macaca fuscata). — J. Comp. Psychol. 130: 153-161. DOI:10.1037/com0000028.
Silk, J.B. & House, B.R. (2011). Evolutionary foundations of human prosocial sentiments. — Proc. Natl. Acad. Sci. USA 108: 10910-10917. DOI:10.1073/pnas.1100305108.
Silk, J.B., Brosnan, S.F., Vonk, J., Henrich, J., Povinelli, D.J., Richardson, A.S., Lambeth, S.P., Mascaro, J. & Schapiro, S.J. (2005). Chimpanzees are indifferent to the welfare of unrelated group members. — Nature 437: 1357-1359. DOI:10.1038/nature04243.
Sterck, E.H.M., Olesen, C.U. & Massen, J.J.M. (2015). No costly prosociality among related long-tailed macaques (Macaca fascicularis). — J. Comp. Psychol. 129: 275-282. DOI:10.1037/a0039180.
Sterck, E.H.M., Crockford, C., Fischer, J., Massen, J.J.M., Tiddi, B., Perry, S., Sueur, C., Schülke, O. & Ostner, J. (2024). The evolution of between-sex bonds in primates. — Evol. Human Behav. 45: 108828. DOI:10.1016/j.evolhumbehav.2024.106628.
Tan, A.W.Y., Hemelrijk, C.K., Malaivijitnond, S. & Gumert, M.D. (2018). Young macaques (Macaca fascicularis) preferentially bias attention towards closer, older, and better tool users. — Anim. Cogn. 21: 1-13. DOI:10.1007/s10071-018-1188-9.
Tennie, C., Jensen, K. & Call, J. (2016). The nature of prosociality in chimpanzees. — Nature Commun. 7: 13915. DOI:10.1038/ncomms13915.
Thierry, B. (2007). Unity in diversity: lessons from macaque societies. — Evol. Anthropol. 16: 224-238. DOI:10.1002/evan.20147.
Thierry, B., Iwaniuk, A.N. & Pellis, S.M. (2000). The influence of phylogeny on the social behaviour of macaques (Primates: Cercopithecidae, genus Macaca). — Ethology 106: 713-728.
Thierry, B., Singh, M. & Kaumanns, W. (2004). Macaque societies: a model for the study of social organization. — Cambridge University Press, Cambridge.
Tomasello, M., Melis, A.P., Tennie, C., Wyman, E. & Herrmann, E. (2012). Two key steps in the evolution of human cooperation: the interdependence hypothesis. — Curr. Anthropol. 53: 673-692. DOI:10.1086/668207.
Van Leeuwen, E.J.C., Detroy, S.E., Kaufhold, S.P., Dubois, C., Schütte, S., Call, J. & Haun, D.B.M. (2021). Chimpanzees behave prosocially in a group-specific manner. — Sci. Adv. 7: eabc7982.
Van Noordwijk, M.A. & Van Schaik, C.P. (1985). Male migration and rank acquisition in wild long-tailed macaques (Macaca fascicularis). — Anim. Behav. 33: 849-861. DOI:10.1016/S0003-3472(85)80019-1.
Van Schaik, C.P. & Van Noordwijk, M.A. (1987). Competition among female long-tailed macaques, Macaca fascicularis. — Anim. Behav. 35: 577-589.
Van Schaik, C.P., Van Noordwijk, M.A., de Boer, R.J. & den Tonkelaar, I. (1983). The effect of group size on time budgets and social behaviour in wild long-tailed macaques (Macaca fascicularis). — Behav. Ecol. Sociobiol. 13: 173-181. DOI:10.1159/000156646.
Vernes, M.K. & Louwerse, A.L. (2010). BPRC’s enrichment manual for macaques & marmosets. — Biomedical Primate Research Center, Rijswijk.
Verspeek, J., van Leeuwen, E.J.C., Laméris, D.W., Staes, N. & Stevens, J.M.G. (2022). Adult bonobos show no prosociality in both prosocial choice task and group service paradigm. — PeerJ 10: 1-24. DOI:10.7717/peerj.12849.
Vonk, J., Brosnan, S.F., Silk, J.B., Henrich, J., Richardson, A.S., Lambeth, S.P., Schapiro, S.J. & Povinelli, D.J. (2008). Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members. — Anim. Behav. 75: 1757-1770. DOI:10.1016/j.anbehav.2007.09.036.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}