Abstract
Orang-utans are well-known for their vocal flexibility. However, the described vocal repertoire of female orang-utans is relatively limited. This anecdotal report discusses the occurrence of long call-like vocalisations in two zoo-housed Bornean orang-utan females. While Temmy demonstrated these vocalisations in response to playbacks of male long calls, Eloise proactively produced vocalisations during her ovulation before her reproductive senescence. The vocalisations produced by Temmy and Eloise seem to differ in many aspects from known orang-utan calls, although they bear some similarities to male long calls. Thus, the vocal repertoire of female orang-utans may be more extensive than previously thought. We carefully suggest that these calls might indicate proceptivity, while also discussing methodological limitations and theoretical considerations.
1. Introduction
The vocal repertoire of primates has been classically characterized as rather inflexible, consisting of mostly innate and fixed vocalisations, but recent studies nuanced this view. The classical perspective suggests that primate vocalisations lacked the complexity and adaptability that are thought to be characteristic of human language (Egnor & Hauser, 2004; Fischer & Price, 2017). However, recent studies have begun to challenge this assumption by revealing a surprising degree of vocal control and vocal learning (Zimmermann, 2017). Pivotal in this shift are findings about the vocal abilities of orang-utans: they can flexibly produce calls that are not part of their species-typical repertoire (Lameira et al., 2016) and have a high level of control over their vocal production, allowing them to produce voiced and unvoiced sounds at the same time (Lameira & Hardus, 2023). Such flexibility might also underlie the variation in repertoire size (Lameira et al., 2022) and call cultures (Wich et al., 2012) in wild orang-utan populations. These studies also suggests that much remains to be learned about orang-utan vocalisations. Therefore, in this anecdotal report, we describe two female orang-utan calls that seem to differ from previously described calls.
Orang-utans are well-known for their male long calls: acoustic signals that are produced by flanged males or males who are developing flanges (Delgado et al., 2008). These calls reach a loudness of approximately 100 decibels (Lameira & Wich, 2008) and carry for about 1 km (Mitani, 1985). These calls are thought to play an important role in regulating social interactions (Delgado, 2006), given that orang-utans are semi-solitary and only sporadically interact with conspecifics (van Schaik, 1999; Roth et al., 2020). Two main functions have been suggested, which are not mutually exclusive (van Schaik & van Hooff, 1996); first, long calls might facilitate inter-individual spacing, especially between males (Mitani, 1985). Second, long calls might function to attract females (Mitra Setia & van Schaik, 2007). Accordingly, Mitra Setia & van Schaik (2007) reported that females approached male long calls, and especially the calls of local dominant males. Altogether, it seems that long calls do not only play a role in intrasexual competition, but also in intersexual competition.
When it comes to female vocalisations, Hardus et al. (2008) report 4 call types that are unique to female orang-utans. Two of those concern vocalisations that are performed during copulation and travel only a short range (mating squeals & fear squeaks). Furthermore, a middle-range call can be found in the form of a roar, an agonistic call that has rarely been observed, and might signal submission. Lastly, females can produce long-distance vocalisations called ‘lork calls’. These calls share similarities with the long call, although their function is still debated. Rijksen (1978) suggested that female lork calls may reflect arousal or agonistic mood. However, the call might also aid in coordinating proximity with other individuals by advertising location (Rijksen, 1978). Altogether, the repertoire of wild orang-utan females contains few middle- and long-distance calls,
Here, we report two cases of long call-like vocalisations in zoo-housed Bornean orang-utan females. Our first case concerns Temmy, who showed a strong bodily and vocal response to male long call playbacks. We observed this phenomenon during a playback experiment in which she at first did not participate. She was temporarily housed in the outside enclosure while we conducted our playback experiment in the indoor enclosure. We opportunistically played long call recordings on 4 consecutive days while she was in the outdoor enclosure and videorecorded her behaviour and vocal responses. Our second case concerns Eloise, who appeared to produce long calls spontaneously during ovulation when she was still reproductively active. In this anecdotal report, we describe the characteristics of the vocalisations. We suggest some functional explanations for these vocalisations.
2. Method
2.1. Temmy
2.1.1. Group and housing
Data for the first case were gathered in Allwetter Zoo (Münster, Germany). During the first study period (23 May 2022 until 27 May 2022), the zoo housed five adult Bornean orang-utans (4F, 1M; Table A1 in the Appendix). During a short second study period (29 July 2022 until 30 July 2022), a new male was also present, who had recently been transported to the zoo. The orang-utan enclosure consisted of an indoor and outdoor enclosure, each capable of housing a subgroup. Alternatively, they could be connected to form one large enclosure.
Temmy, the female of interest in this case, had been living in Allwetter Zoo for over 20 years at the time of research. She had not been exposed to a flanged male since the death of the previous flanged male at the start of 2020. During the first study period, Temmy was housed solitarily in the outside enclosure until ±15.30, when Mr. Miyagi was also allowed to move to the outside enclosure. Between 16.30–17.00 they were called inside for the night. During the second study period, Temmy and Mr. Miyagi were housed together (either in the indoor or the outdoor enclosure), while the rest of the group was already introduced to the new flanged male.
2.1.2. Stimuli
The long call recordings that we used for the playback experiment were collected at Sikundur Research Station (Sumatra, Indonesia) by James Askew (2019). We used both long calls made by fully flanged males and long calls made by transitioning males. From all the recordings, we selected only calls classified as ‘spontaneous’. As control sounds we used two calls of the Great argus pheasant (Argusianus argus), a bird species that occurs in the natural habitat of orang-utans. We edited all stimuli to last 45 s so that each stimulus had the same duration. Furthermore, all recordings were normalized so that the volume level was similar across stimuli.
2.1.3. Design of trial
We made this observation during a playback study in which Temmy did not participate. She was housed in the outdoor enclosure while we performed our experiment indoors. For this experiment, we exposed the orang-utans to playbacks of flanged male long calls, transitioning male long calls, and control sounds. After observing Temmy’s strong reactions to the playbacks in the inside enclosure (see Results), we decided to opportunistically play her some of the stimuli in the outdoor enclosure as well. However, due to restricted time and the unexpected nature of the event, this was not in a fully controlled setup.
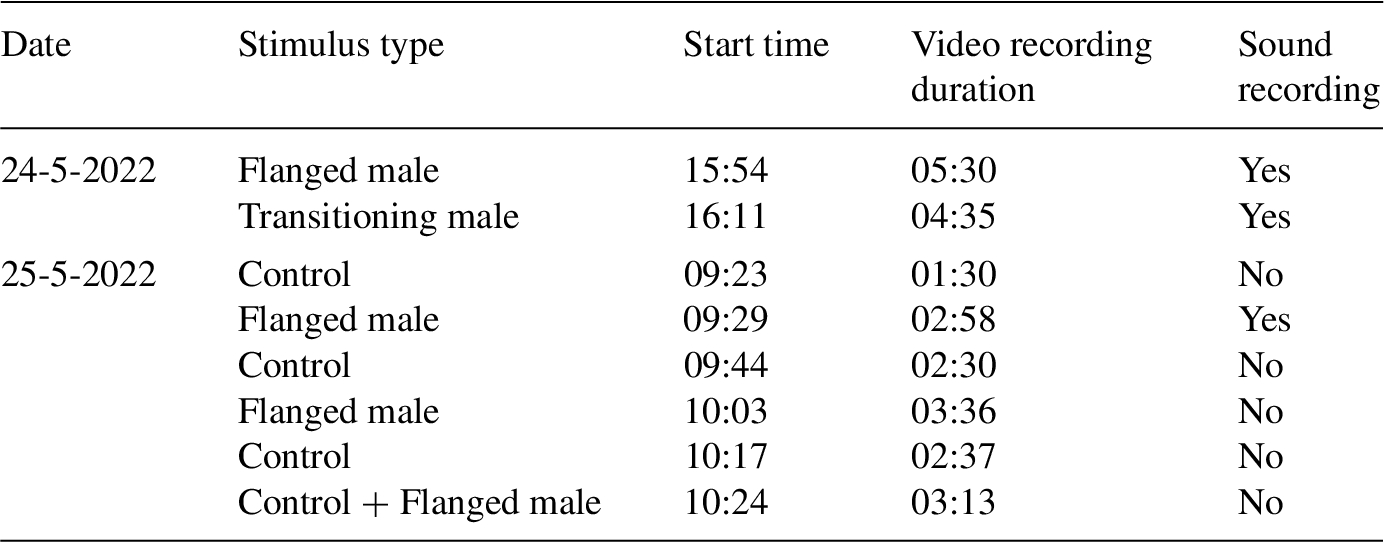
On the first day, we played her a flanged male long call and a transitioning male long call, after which she was moved inside. On the second day, we tried to employ an ABABA design where we switched between control sounds and flanged male sounds. Hereafter, we also added one trial where the control stimulus was immediately followed by a flanged male sound, so that we could clearly see the difference in behaviour while she was located in the same area of the enclosure for both playbacks. However, it is important to note that the restricted time resulted in very short intervals between the trials, which is not ideal given that carry-over effects might occur. We included an overview of the trials on the 24th and morning of the 25th, when she behaviourally and vocally responded to the long call playbacks, in Table A2 in the Appendix.
We made audio recordings of the vocal response using a Sennheiser ME 67 microphone and a Tascam DR-40X recorder. Two camcorders (study period 1) or one camcorder and one action camera (study period 2) were used to videorecord the behavioural response to the stimuli. To play the stimuli, we used a custom loudspeaker linked to a Marantz PMD 620 unit. We played the stimuli at ±90 dB at 1 m from the loudspeaker.
The setup of the speakers was in line with recommendations (Fischer et al., 2013): while one researcher was responsible for placing the speaker and playing the sound, one or two other researchers videorecorded the response. In addition, one of the researchers carried the microphone that was used to record Temmy’s vocalisations opportunistically. Before starting a trial, we would first identify Temmy’s location. After that, we would determine a suitable location for the speaker. We did not want the speaker to be clearly visible to Temmy at the onset of the trial. After we chose a location for the speaker, we would start the stimulus playback of 45 s and immediately videorecord her response. When seeing a clear behavioural and vocal response, we would continue videorecording her response until her behaviour seemed to be back to baseline (i.e., when she displayed no more conspicuous behavioural responses and/or stopped vocalising). If we saw no clear response, we aimed to record at least 30 s after the end of the stimulus, although some videos ended up slightly shorter.
2.2. Eloise
Data for the second case were obtained from Los Angeles Zoo (Los Angeles, CA, USA) as part of JA’s thesis (Askew, 2019). The recordings were made by MF in 2017. This case concerns Eloise, a Bornean orang-utan female who was born in LA Zoo on 10 November 1968, and has been living in LA Zoo since then. Over time, she has been housed with multiple individuals, including different males. She gave birth to 4 offspring from 3 different males between 1978 and 1987. In our case description, we focus on the period from 1999 until approximately 2018, date after which she is considered to be of post-reproductive age (based on the absence of a regular menstrual cycle). During the period that we describe, she was housed with a flanged male and with different females, and had visual and olfactory access to another flanged male in the same facility. Furthermore, she was not on contraceptives at any time during this period.
Eloise has a condition similar to cerebral palsy, which she has had since birth. Her condition mostly affects her locomotion, since she has limited use of her hands and especially her feet. She has learned over her years how to modify her movements, and has generally locomoted by rolling or crawling. However, she has still been able to have a normal social life with conspecifics, and has been able to climb and manoeuvre throughout her exhibit space.
Importantly, one of the authors of this manuscript (MF) was Eloise’s primary caregiver in the zoo during the period that we describe. Our description of Eloise’s behaviour is based on her experiences.
2.3. Vocalisation analyses
Bioacoustic analyses were performed using Audacity software (Audacity Team, 2011). Measurements were taken of the total call length, inter-onset-intervals of the separate vocalisations, peak frequencies, and fundamental frequencies. Based on these parameters, the vocalisations of Temmy and Eloise were compared to other orang-utan calls obtained from Orangutan network of Universität Zurich (https://www.aim.uzh.ch/en/research/orangutannetwork/orangutancallrepertoires.html; Hardus et al., 2008). Furthermore, we compared Temmy’s and Eloise’s call with male long calls recorded by James Askew (2019; also see ‘Stimuli’).
2.4. Data availability statement
The data associated with this research are available at Dataverse (https://doi.org/10.34894/JNUTYU).

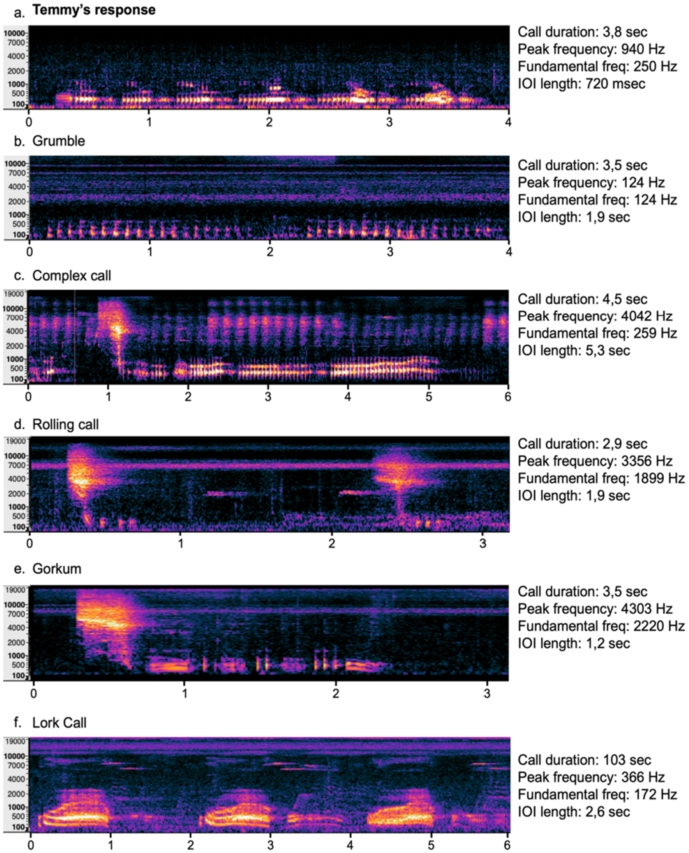
Comparison between Temmy’s vocal response to other vocalisations known in the orang-utan repertoire. Analysed aspects of the vocalisations were: total call duration (s), Peak frequency (loudest frequency of the call), fundamental frequency (lowest frequency of the call), Inter-onset-interval (time between start of one vocalisation to start of next vocalisation within the call). Panels b–f indicate 5 different orangutan calls that bear certain similarities to Temmy’s vocal response. Neither of them can be categorized as the same call that Temmy is producing.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10273
3. Results
3.1. Vocal analyses
3.1.1. Temmy
During the playbacks on the first two observation days, Temmy performed vocalisations (Audio 1 at https://stream.cadmore.media/player/397a06c8-afa9-4818-b2c5-55ae18034cb7). We were able to capture three good quality recordings of the vocalisation that Temmy produced. These calls consisted of short series of 5 to 6 consecutive bark-like vocalisations (hereafter pulses; Figure 1). Each call lasted on average 4.2 s (SD = 0.3), in the longest recording these calls were repeated consecutively for more than 3 min. Each bark-like pulse within the call had an average duration of 690 ms long (SD = 48 ms), with an average fundamental frequency of 250 Hz. These parameters remained consistent across all of Temmy’s vocalisations. Only the intensity of the calls decreased sharply after 65 s of vocalising. Still, the softer calls remained similar in the other acoustic parameters.
Interestingly, the calls that Temmy emitted in response to the playbacks seem to be different from any calls generally known to be uttered by orang-utans. To validate this, we compared Temmy’s calls to known sounds in the orang-utan repertoire: the grumble, complex call, rolling call, gorkum and lork call. These comparisons were based on 4 main features, call duration (s), peak frequency (Hz), fundamental frequency (Hz) and inter-onset-interval length (s) (Figure 1). The complex call, rolling call and gorkum all have one high frequency burst in the 3500 Hz range, something that is missing from Temmy’s vocalisations during the playbacks. However, based on the temporal structure, these calls are rather similar. Temmy’s vocalisations might be a variation on the complex call that lacks the high burst and where the emphasis is mainly on the low-frequency grunts. Based on the frequency range, there is more similarity with the grumble, a call with a quick repetition of short and low pulses. These same pulses seem to occur in Temmy’s vocalisations. However, Temmy’s are higher in frequency (250 instead of 150 Hz) and occur in more tightly knitted sets. The lork call seems to be most similar to Temmy’s vocalisations. Both are relatively long calls (>60 s) with low frequency pulses. Nevertheless, the lork calls are much slower than Temmy’s vocalisations. Both the separate pulses as well as the pauses between these pulses are about double the length in the lork calls than in Temmy’s vocalisations. This means that these vocalisations of Temmy in response to the male long call share features of some of the known calls from the orang-utan repertoire but bear no complete similarity to any of them.
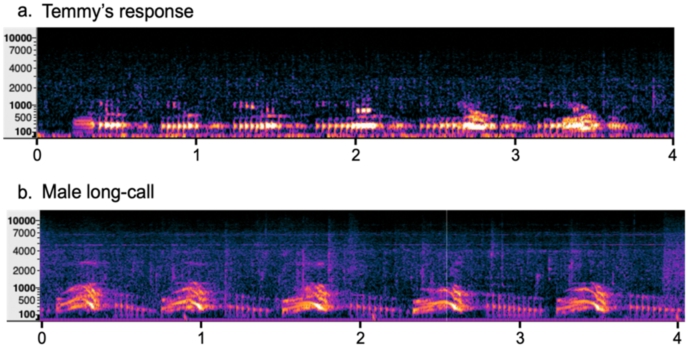
Temmy’s vocalisations seem to be most similar to the long calls of male orang-utans (Figure 2). Male long calls also consist of bark-like pulses that average around 800 ms each (average IOI of 1.5 s) with a fundamental frequency of around 320 Hz. An analysis of the spectral information also shows that most of the power in these calls occurs in the same spectrum, between 200 and 350 Hz, with little to no power in the higher frequencies. Regarding the temporal structure, these calls bear strong resemblance in the sound onset timing. In addition, the duration of each pulse is similar between Temmy’s vocalisations and male long calls. Temmy’s pulses have inter-onset-intervals (IOIs) of approximately 740 ms (SD = 61 ms), whilst the 12 male long calls that we analysed have IOIs of approximately 800 ms (SD = 54 ms). The clearest difference between Temmy’s vocalisations and the male long calls lies in the overall temporal structure. Long calls consist of rather regular repetitions of pulses that can last up to 3 min with steady IOIs. Temmy’s calls could also last up until 3 min in total, but with concatenations of bouts of 5–6 pulses, followed by a longer pause (±2.5 s) before there was another pulse-bout. The IOIs within these bouts are similar to male long calls, but the pauses between bouts are not typical for long calls. To conclude, Temmy’s call seems to be most similar in temporal structure and frequency distribution to the male long calls and less so to other orang-utan vocalisations.
Male long call spectrogram in comparison with Temmy’s vocal response. Male long calls have a longer total duration than Temmy’s vocalisations, but the pulses bear many resemblances in spectral and temporal pattern.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10273
3.1.2. Eloise
A second female, Eloise, proactively produced calls that also bear similarities with male long calls (Audio 2 at https://stream.cadmore.media/player/3852edf8-260e-4d33-87d2-42db6a7c073c; Figure 3). Two good-quality recordings of these calls were collected in 2017. Rather than the reactive calls of Temmy to playbacks, this female produced long call-like calls spontaneously. Eloise’s calls mainly differ from Temmy’s with respect to timing. Each pulse is much longer, with an average of 2.237 s per pulse (average IOI 2.467 s). This makes her pulses also longer than those of the male long call. However, unlike the concatenated pulse-bouts in Temmy’s calls, Eloise’s full calls retained a regular IOI pattern for the whole duration. One call could continue for up to 60 s with steady IOIs, which bears more similarity to the long stable duration of the long calls which generally last between 60 and 120 s. With respect to the fundamental and peak frequency, Eloise’s calls were closer to the male long call compared to Temmy’s (average Eloise fundamental frequency: 300 Hz, male long call around 350 Hz, Figure 3).
Eloise’s vocalisations in comparison to the male long call. The pulses in Eloise’s call are longer than those of the male-long call. However, the overall call structure and total call length are similar. The spectral information of the call also bears many similarities to the male long call.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10273
These calls seem to bear little resemblance to known orang-utan calls. It might come closest to the complex call which has also relatively long and low call units. However, the units in the complex call are produced as several short, consecutive bursts, whilst the long call-like call of Eloise is one long burst on an exhale, followed by an audible inhale. Especially the long duration of the pulses and the total call duration of Eloise’s vocalisations make them incomparable to any of the known orangutan calls.
3.2. Behavioural context
3.2.1. Temmy
Temmy was housed in the outside enclosure while we were performing a playback experiment in the indoor enclosure with the four other orang-utans housed in Allwetter Zoo. On the first day of this experiment, we noticed that Temmy showed a strong behavioural reaction to the long calls, but not the controls, that we used in the playback experiment, because she would come and sit in front of a semi-transparent door that connected the indoor and outdoor enclosure. Because of her strong reaction to the long call stimuli, we asked permission to opportunistically expose her to some stimuli in the outside enclosure. We started doing this on the afternoon of the first day.
On the afternoon after we had finished with the indoor trials for the playback study, we played two long calls to Temmy, in the outer enclosure (one flanged male call, one transitioning male call). In both cases she showed a strong behavioural response (Video 1 at https://stream.cadmore.media/player/9db25086-79e4-4f48-af5d-318fbb170ad2 and Video 2 at https://stream.cadmore.media/player/14a82a1e-6556-44b7-9d46-8803cc1e14f3). Her response to the long calls was characterised by fast movement in the direction of the speaker, climbing into climbing structures, and standing/sitting while looking in the direction of the speaker. Furthermore, Temmy started vocalising for multiple minutes in a row. After this, she and Mr. Miyagi were moved to the night enclosures. On the morning of day 2, we resumed playing long calls to Temmy in the outside enclosure. However, we now also included control sounds to determine whether she would show the same behavioural response to all sounds. In total, we played three long calls and four control sounds. Temmy showed responses similar to the day before when we played long calls, including vocalisations. However, she showed no discernible behavioural response when we played a control sound, although she vocalised very shortly in response to one control trial that started after a long call trial. Crucially, she quickly approached the source of the stimuli in response to long calls, but not to control sounds (Video 3 at https://stream.cadmore.media/player/36a876d8-b519-4518-bc63-797357592b69 and Video 4 at https://stream.cadmore.media/player/d140ee06-de8a-4c5c-bff2-a1007c7f03a1).
On the afternoon of day 2, we wanted to continue the experiment by playing more stimuli to Temmy. In this case, we played two long calls and two control stimuli. From this point onwards, she did not respond to both the control sounds and the long call stimuli anymore. In a second study period, 66 days later, we again presented her with both control sounds and long calls, both of which did not result in any apparent responses.
3.2.2. Eloise
Eloise, a Bornean orang-utan female living in Los Angeles Zoo, had been producing unique vocalisation since she became of reproductive age. Although no consistent records exist with regard to this specific behaviour at earlier stages of her life, one of the authors (MF) has worked as her main caregiver since 1999. The observations below are based on her experiences with Eloise, who consistently produced vocalisation until approximately 2018. The vocalisations consistently co-occurred with her peri-ovulatory period, as evidenced by the fact that Eloise consistently produced the vocalisations approximately two weeks after onset of her menstruation on a monthly basis. Furthermore, her vocalisations coincided with increased proceptivity towards the flanged male that she was housed with, reflected in following the male and actively soliciting him.
During the monthly vocalisation time window, Eloise would actively pursue the male, while outside of her ovulation timeframe she would not actively seek him out. This would include her following the male throughout the exhibit space when he would move areas. She would also position herself for copulation, trying to solicit a response from the male. The male would copulate with Eloise, and would occasionally want to be housed with her overnight. Given Eloise’s condition, she was subordinate to the other females in the group. If any of the other females were ovulating at the same time as Eloise, she would be supplanted and pushed away by the females. Otherwise, these females would not intervene. Her ovulation lasted approximately 3–4 days, with the vocalisations being most intense during the first day or two of the cycle and slowly tapering off.
Eloise has stopped producing her vocalisations since she transitioned towards her post-reproductive period. When her menstrual cycle started to become less regular, her calls became less frequent and eventually disappeared.
4. Discussion
In this anecdotal report, we describe two cases of female long call-like vocalisation in zoo-housed Bornean orang-utans. First, we described the case of Temmy, who showed a vocal response when we opportunistically played long call recordings to her, which disappeared after two days. In a short follow-up experiment, she did not show the same vocal response either. Second, we described the case of Eloise, a female who consistently produced specific vocalisations, presumably during ovulation. When she reached post-reproductive age, she ceased producing this vocalisation. Our observations suggest that the vocal repertoire of female orang-utans might be more elaborate than previously thought (Hardus et al., 2008), perhaps owing to the increasingly recognized flexibility in orangutan vocalisations (Lameira et al., 2022). While Temmy’s vocalisations seemed to travel a relatively short distance, Eloise’s vocalisations seemed much louder, and could be considered middle- or even long-distance calls (sensu Hardus et al., 2008). Even though we want to emphasize that anecdotal reports should be interpreted with caution (Sándor & Miklósi, 2020), we try to suggest functional explanations for our observations and provide suggestions on how to further study these explanations.
With regard to the calls produced by Temmy and her behaviour during the playbacks, there are two explanations that strike us as most likely. First, it could be the case that her vocalisations are reflective of negative arousal in response to an unexpected stimulus. After habituating to the long call stimuli, we would then expect a reduction in her response, exactly as we observed. We tried to somewhat control for habituation by playing her the same recording two months later, hoping that this would be a long enough period for spontaneous recovery to occur (Rankin et al., 2009). This would indeed suggest that habituation had occurred during our first study period. However, we saw no more vocal responses by Temmy during the second study period, nor any conspicuous behavioural responses, which is not in line with the habituation hypothesis. Nevertheless, it is important to note that the social structure of the group had also changed between our first and second study period, because a new flanged male was present in the zoo. Although he was not housed with Temmy yet, they were housed in the same building, meaning that she could have been exposed to his long calls, which resulted in further habituation to long call stimuli.
An alternative explanation would be that Temmy’s vocal response and behavioural response was driven by her peri-ovulatory status. Previous work has suggested that male long calls may function to attract sexually active females (Mitra Setia & van Schaik, 2007), and in this case it could be that Temmy showed a positive response to the long call stimuli because of her ovulatory status. It is important to emphasize that we lack hormonal evidence of ovulation. Nevertheless, we have two reasons to assume that Temmy could be ovulating during our first observation period. Firstly, Temmy was not on contraceptives. Secondly, according to the curator and the orang-utan caretakers, Temmy started the peri-ovulatory period of her menstrual cycle right before the first study period, evidenced by a cooperative mating event between Mr. Miyagi and Temmy two days prior to the start of our study. Such matings are known to be more common during the peri-ovulatory period (Durgavich et al., 2023). In orang-utans, this part of the cycle lasts approximately 5 days, similar to humans (Kubátová & Fedorova, 2016). This would explain why her vocal and behavioural response to the long call stimuli decreased after 2 days. Furthermore, the fact that Temmy did not show a similar response during the second study period would also fit with this explanation. An essential step to establish a link between ovulation and Temmy’s vocal response would be to record her response to an intraspecific control, such as a female call. However, we did not have such intraspecific calls available during the study period. We tried to play her a transitioning male call on the first day, thinking it might be less arousing, but she responded in a similar way when compared to her response to flanged male long calls.
In the case of Eloise, the evidence for a link between her peri-ovulatory status and her vocalisations is much stronger, since her vocalisations followed approximately two weeks after her menstruation and were also accompanied by proceptive behaviour towards the male she was housed with. This is in line with results from restricted-access tests (Maple et al., 1979; Nadler, 1995): when orang-utan males have free access to females, mating events are quite evenly spread across the menstrual cycle. However, when females can control access, they specifically approach males during ovulation. This is also supported by the fact that wild orang-utan females selectively mate with flanged males during their peri-ovulatory period (Knott et al., 2009). Thus, potentially Eloise’s vocalisations could be considered proceptive behaviour towards the male.
Potentially, such peri-ovulatory vocalisations could function to coordinate movement. Male long calls have such a coordinative function (Mitani, 1985; Delgado et al., 2008), but it could well be that female vocalisations have a similar function. Although vocalisations that indicate proceptivity seem to be rare in primates (Dixson, 2012), females of at least three other relatively solitary primate species seem to produce proceptive vocalisations that can travel relatively long distances (aye-aye, Daubentonia madagascariensis: Sterling & Richard, 1995; grey mouse lemur, Microcebus murinus: Buesching et al., 1998; lion-tailed macaque, Macaca silenus: Lindburg, 1990). Given that Eloise’s vocalisation was relatively loud, the mate attraction hypothesis would be a likely candidate to explain her vocalisations. However, Temmy’s call seems to travel relatively short distance. Potentially, short-distance vocalisations like Temmy’s communicate mating motivation or induce arousal in males. Such vocalisations have also been described in other primate species (Dixson, 2012), such as pre-copulatory vocalisations in gelada (Theropithecus gelada) females that are especially produced during ovulation (Roberts et al., 2017) and peri-ovulatory mounting vocalisations in tufted capuchins (Cebus apella; Carosi & Visalberghi, 2002). Thus, female vocalisations that indicate proceptivity have previously been described in other primate species, and might be present in orang-utans, although this requires further investigation.
It is important to note, though, that this functional account is in conflict with an important aspect of orang-utan sexuality, namely concealed ovulation in females. Orang-utan females show no outward indicators of ovulation (Knott & Kahlenberg, 2011). It has been suggested that concealment of ovulation reduces infanticide risk through paternity confusion in orang-utans (Kunz et al., 2022). If concealing ovulation is indeed beneficial for females, peri-ovulatory vocalisations could be detrimental. However, our cases concern two females from the zoo-housed population, and such housing is often associated with changes in selective pressures (Williams & Hoffman, 2009). Specifically, orang-utan females are often housed with only one breeding male in zoos, reducing the likelihood of infanticide, while wild orang-utan females need to navigate a more complex social environment, with multiple males that might pose a threat. Therefore, peri-ovulatory vocalisations might occasionally be observed in zoo-housed females, despite the fact that they may be under strong negative selection in wild orang-utan populations.
Given the anecdotal nature of our observations, we cannot conclude that peri-ovulatory vocalisations exist in orang-utans. However, our report could be a starting point for a well-controlled playback experiment in the future that might probe reactive calls from females in a peri-ovulatory state, as might have been the case for Temmy. By exposing females over a longer time period to playbacks of long calls, other conspecific calls as intraspecific control, and ambient controls, it would be possible to disentangle a general habituation explanation from a specific peri-ovulatory vocal response to male long calls. However, it will probably still be challenging to perform such an experiment with zoo-housed animals, since they are often either on contraceptives or carrying a dependent infant, which may modulate their behavioural response to such playbacks. Therefore, it will remain difficult to obtain a large enough sample size in such designs.
In conclusion, we described two long call-like calls produced by zoo-housed female Bornean orang-utans, suggesting a more extensive repertoire of orang-utan female vocalisations, possibly resulting from high degrees of vocal flexibility. Our observations open up possibilities that the orang-utan female vocal repertoire might include vocalisations that indicate proceptivity, although careful experimental approaches will be necessary to substantiate this hypothesis.
Corresponding author’s e-mail address: t.s.roth@uu.nl
Acknowledgements
We want to thank curator Miriam Göbel and all the orang-utan caretakers at Allwetter Zoo for supporting our research. In addition, we want to thank Evy van Berlo for helping with data collection in the second study period in Allwetter Zoo. Furthermore, we want to thank Cathleen Cox (Los Angeles Zoo) for permission to share the audio recordings of Eloise.
References
Askew, J.A. (2019). Geographic variation in the feeding ecology and long-distance vocalizations of orangutans. — University of Southern California, Los Angeles, CA.
Buesching, C.D., Heistermann, M., Hodges, J.K. & Zimmermann, E. (1998). Multimodal oestrus advertisement in a small nocturnal prosimian, Microcebus murinus. — Folia Primatol. 69: 295-308.
Carosi, M. & Visalberghi, E. (2002). Analysis of tufted capuchin (Cebus apella) courtship and sexual behavior repertoire: Changes throughout the female cycle and female interindividual differences. — Am. J. Phys. Anthropol. 118: 11-24.
Delgado, R.A. (2006). Sexual selection in the loud calls of male primates: signal content and function. — Int. J. Primatol. 27: 5-25.
Delgado, R.A., Lameira, A.R., Davila Ross, M., Husson, S.J., Morrogh-Bernard, H.C. & Wich, S.A. (2008). Geographical variation in orangutan long calls. — In: Orangutans: Geographic variation in behavioral ecology and conservation (Wich, S.A., Utami Atmoko, S.S., Setia, T.M. & van Schaik, C.P., eds). Oxford University Press, Oxford, p. 215-224.
Dixson, A.F. (2012). Primate sexuality: Comparative studies of the prosimians, monkeys, apes, and humans. — Oxford University Press, Oxford.
Durgavich, L.S., Harwell, F.S. & Knott, C.D. (2023). A composite menstrual cycle of captive orangutans, with associated hormonal and behavioral variability. — Am. J. Primatol. 85: e23420.
Egnor, S.E.R. & Hauser, M.D. (2004). A paradox in the evolution of primate vocal learning. — Trends Neurosci. 27: 649-654.
Fischer, J., Noser, R. & Hammerschmidt, K. (2013). Bioacoustic field research: A primer to acoustic analyses and playback experiments with primates. — Am. J. Primatol. 75: 643-663.
Fischer, J. & Price, T. (2017). Meaning, intention, and inference in primate vocal communication. — Neurosci. Biobehav. Rev. 82: 22-31.
Hardus, M.E., Lameira, A.R., Singleton, I., Morrogh-Bernard, H.C., Knott, C.D., Ancrenaz, M., Utami Atmoko, S.S. & Wich, S.A. (2008). A description of the orangutan’s vocal and sound repertoire, with a focus on geographic variation. — In: Orangutans: Geographic variation in behavioral ecology and conservation (Wich, S.A., Utami Atmoko, S.S., Setia, T.M. & van Schaik, C.P., eds). Oxford University Press, Oxford, p. 49-64.
Knott, C.D. & Kahlenberg, S.M. (2011). Orangutans: Understanding forced copulations. — In: Primates in perspective, 2nd edn. (Campbell, C.C., MacKinnon, K.C., Bearder, S.K., Panger, M.A. & Fuentes, A., eds). Oxford University Press, Oxford, p. 313-326.
Knott, C.D., Thompson, M., Stumpf, R.M. & McIntyre, M.H. (2009). Female reproductive strategies in orangutans, evidence for female choice and counterstrategies to infanticide in a species with frequent sexual coercion. — Proc. Roy. Soc. Lond. B: Biol. Sci. 277: 105-113.
Kubátová, A. & Fedorova, T. (2016). Saliva crystallization occurs in female Bornean orangutans (Pongo pygmaeus): Could it be a new option for monitoring of menstrual cycle in captive great apes? — PLoS ONE 11: e0159960.
Kunz, J.A., van Noordwijk, M.A. & van Schaik, C.P. (2022). Orangutan sexual behavior. — In: The Cambridge handbook of evolutionary perspectives on sexual psychology Volume 4: Controversies, applications, and nonhuman primate extensions (Shackelford, T., ed.). Cambridge University Press, Cambridge, p. 401-425.
Lameira, A.R. & Hardus, M.E. (2023). Wild orangutans can simultaneously use two independent vocal sound sources similarly to songbirds and human beatboxers. — Proc. Natl. Acad. Sci. USA Nexus 2: pgad182.
Lameira, A.R., Hardus, M.E., Mielke, A., Wich, S.A. & Shumaker, R.W. (2016). Vocal fold control beyond the species-specific repertoire in an orang-utan. — Sci. Rep. 6: 30315.
Lameira, A.R., Santamaría-Bonfil, G., Galeone, D., Gamba, M., Hardus, M.E., Knott, C.D., Morrogh-Bernard, H., Nowak, M.G., Campbell-Smith, G. & Wich, S.A. (2022). Sociality predicts orangutan vocal phenotype. — Nature Ecol. Evol. 6: 644-652.
Lameira, A.R. & Wich, S.A. (2008). Orangutan long call degradation and individuality over distance: A playback approach. — Int. J. Primatol. 29: 615-625.
Lindburg, D.G. (1990). Proceptive calling by female lion-tailed macaques. — Zoo Biol. 9: 437-446.
Maple, T.L., Zucker, E.L. & Dennon, M.B. (1979). Cyclic proceptivity in a captive female orang-utan (Pongo pygmaeus abelii). — Behav Process. 4: 53-59.
Mitani, J.C. (1985). Sexual selection and adult male orangutan long calls. — Anim. Behav. 33: 272-283.
Mitra Setia, T. & van Schaik, C.P. (2007). The response of adult orang-utans to flanged male long calls: inferences about their function. — Folia Primatol. 78: 215-226.
Nadler, R.D. (1995). Sexual Behavior of Orangutans (Pongo pygmaeus). — In: The Neglected Ape (Nadler, R.D., Galdikas, B.F.M., Sheeran, L.K. & Rosen, N., eds). Springer, Boston, MA, p. 223-237.
Rankin, C.H., Abrams, T., Barry, R.J., Bhatnagar, S., Clayton, D.F., Colombo, J., Coppola, G., Geyer, M.A., Glanzman, D.L., Marsland, S., McSweeney, F.K., Wilson, D.A., Wu, C.-F. & Thompson, R.F. (2009). Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. — Neurobiol. Learn. Memor. 92: 135-138.
Rijksen, H.D. (1978). A field study on Sumatran orang utans (Pongo pygmaeus abelii Lesson 1827): ecology, behaviour and conservation. — Phd thesis, Landbouwhogeschool Wageningen.
Roberts, E.K., Lu, A., Bergman, T.J. & Beehner, J.C. (2017). Female reproductive parameters in wild geladas (Theropithecus gelada). — Int. J. Primatol. 38: 1-20.
Roth, T.S., Rianti, P., Fredriksson, G.M., Wich, S.A. & Nowak, M.G. (2020). Grouping behavior of Sumatran orangutans (Pongo abelii) and Tapanuli orangutans (Pongo tapanuliensis) living in forest with low fruit abundance. — Am. J. Primatol. 82: e23123.
Sándor, K. & Miklósi, Á. (2020). How to report anecdotal observations? A new approach based on a lesson from “puffin tool use”. — Front. Psychol. 11: 555487.
Sterling, E. & Richard, A. (1995). Social organization in the aye-aye (Daubentonia madagascariensis) and the perceived distinctiveness of nocturnal primates. — In: Creatures of the dark (Alterman, L., Doyle, G.A. & Izard, M.K., eds). Springer, Boston, MA, p. 439-451.
van Schaik, C.P. (1999). The socioecology of fission–fusion sociality in orangutans. — Primates 40: 69-86.
van Schaik, C.P. & van Hooff, J.A.R.A.M. (1996). Toward an understanding of the orangutan’s social system. — In: Great ape societies (Marchant, L.F., Nishida, T. & McGrew, W.C., eds). Cambridge University Press, Cambridge, p. 3-15.
Wich, S.A., Krützen, M., Lameira, A.R., Nater, A., Arora, N., Bastian, M.L., Meulman, E., Morrogh-Bernard, H.C., Atmoko, S.S.U., Pamungkas, J., Perwitasari-Farajallah, D., Hardus, M.E., van Noordwijk, M. & van Schaik, C.P. (2012). Call cultures in orang-utans? — PLoS ONE 7: e36180.
Williams, S.E. & Hoffman, E.A. (2009). Minimizing genetic adaptation in captive breeding programs: A review. — Biol. Conserv. 142: 2388-2400.
Zimmermann, E. (2017). Evolutionary origins of primate vocal communication: diversity, flexibility, and complexity of vocalizations in basal primates. — In: Primate Hearing and Communication (Quam, R.M., Ramsier, M.A., Fay, R.R. & Popper, A.N., eds). Springer International, Cham, p. 109-140.
Orang-utans housed at Allwetter Zoo during the study periods.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10273
Details of the playback trials performed with Temmy on the afternoon of the first test day and the morning of the second test day.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10273
Supplementary material
Audio 1. This file can be accessed at https://stream.cadmore.media/player/397a06c8-afa9-4818-b2c5-55ae18034cb7.
Audio 2. This file can be accessed at https://stream.cadmore.media/player/3852edf8-260e-4d33-87d2-42db6a7c073c.
Video 1. This file can be accessed at https://stream.cadmore.media/player/9db25086-79e4-4f48-af5d-318fbb170ad2.
Video 2. This file can be accessed at https://stream.cadmore.media/player/14a82a1e-6556-44b7-9d46-8803cc1e14f3.
Video 3. This file can be accessed at https://stream.cadmore.media/player/36a876d8-b519-4518-bc63-797357592b69.
Video 4. This file can be accessed at https://stream.cadmore.media/player/d140ee06-de8a-4c5c-bff2-a1007c7f03a1.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}