Abstract
Acoustic signalling mediates key animal behaviours, but the factors driving inter- and intra-specific variation remain elusive, especially in non-learning suboscine birds. Acoustic traits may evolve to signal an individual’s quality through a process termed honest signalling. Most studies on bird song evolution via honest signalling focus on oscine bird species (song learners) while fewer evaluate honest signalling in suboscines. We tested whether song was an honest signal of male quality in southwestern willow flycatchers (Empidonax traillii extimus), a suboscine, by monitoring nest success and recording male advertisement songs. Based on song characteristics known to vary among individuals, we found higher minimum song frequency, but not temporal components, was positively correlated with the number of fledglings and nest success, after accounting for first egg date, the number of eggs, and clutch number. Our study therefore offers evidence that suboscine song may have evolved as an honest signal of reproductive success.
1. Introduction
Song is used by birds to attract mates (Collins, 2004), but the reasons why particular songs are selected in mating decisions, and the evolutionary mechanisms mediating preferred song traits, remain unresolved (Andersson, 1994). Song characteristics may evolve through genetic drift, natural selection, sexual selection, cultural selection, arbitrary mate choice, or a combination of these factors (Prum, 2010). Under the sexual selection model, mating decisions are based on preferred traits, and numerous studies suggest song is an honest signal of reproductive success, that is, song indicates quality to potential mates (reviewed in Soma & Garamszegi, 2011, but see Garamszegi & Møller, 2004; Byers & Kroodsma, 2009). Mates may choose songs that signal higher quality (Baker et al., 1986), as this increases the mate’s own fitness directly and/or indirectly (Andersson, 1994; Johnstone, 1995; Kokko et al., 2003). If songs signal quality, song traits should then be correlated with an individual’s quality such as reproductive output (McGregor et al., 1981; Kokko et al., 2002; Safran et al., 2013).
A meta-analysis assessing the relationship between song and genetic and social (apparent) reproductive success found song predicted the number of mates and offspring; however, this was biased towards oscine songbirds (Soma & Garamszegi, 2011). Studies of suboscines found nest initiation, participation during dawn chorus (Sexton et al., 2007; Murphy et al., 2008; Bruni & Foote, 2014), and song structure (Garamszegi, 2005; Araya-Ajoy et al., 2009) predicted mate quality, reproductive and/or mating success, and brood numbers. Generally, however, whether suboscine song characteristics signal reproductive success is less understood compared to song learners (but see Araya-Ajoy et al., 2009). Song characteristics such as frequency and temporal aspects may be selected to optimize transmission given the ecological context (Marten & Marler, 1977; Marten et al., 1977) or because they signal mate quality, or both, or they may be arbitrarily selected (Prum, 2010). Suboscines are a useful group in which to explore the role of sexual selection in shaping song characteristics because song variation is not confounded by other sources of variation such as cultural learning (Tobias et al., 2012). Therefore, if song is related to reproductive success, this may be strong evidence that song is an honest signal of mate quality (Safran et al., 2013).

Southwestern willow flycatcher (Empidonax traillii extimus) ‘fitz-bew’ songs are their main advertising vocalization. Individuals that sang lower frequency songs (left panel) produced fewer offspring than individuals that sang higher frequency songs (right panel,
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275
The southwestern willow flycatcher (Empidonax traillii extimus) is a subspecies of the willow flycatcher that occurs in the Southwestern United States. Willow flycatchers are suboscines with no evidence of song learning (Kroodsma, 1984). Their main advertising vocalization is the ‘fitz-bew’ song (Figure 1), which varies among and is unique to individuals (Fernández-Juricic et al., 2009). In studies assessing geographic song variation in willow flycatchers, minimum song frequencies and frequency modulations of the song’s final notes explained the most variation among individuals (Figure 1; Sedgwick, 2001; Mahoney et al., 2020), but the factors mediating individual variation remain unknown. Larger bodied tyrannid species with heftier bills sing lower frequency and slower paced songs (Schoen et al., 2023), however body size (estimated by wing chord) and bill morphology (length and width) do not vary widely among willow flycatcher individuals (male museum specimens vary on average 4% for wing chord, 3% for bill width, and 5% for nares length; Unitt, 1987), suggesting an alternative factor is shaping song in this species. Sex ratios in willow flycatchers range from 40% female and 60% male to 71% female and 29% male (Paxton et al., 2002), indicating sexual selection may be acting within some populations. Predicting reproductive success acoustically would be important from an applied perspective as well, because practitioners could remotely identify fecund pairs. We tested the honest signalling hypothesis in southwestern willow flycatchers and specifically assessed if minimum song frequencies and frequency modulations (shown to vary among willow flycatcher individuals, Mahoney et al., 2020) were correlated with reproductive success.
2. Methods
2.1. Nest monitoring
We monitored southwestern willow flycatcher nests (
2.2. Song recordings and analyses
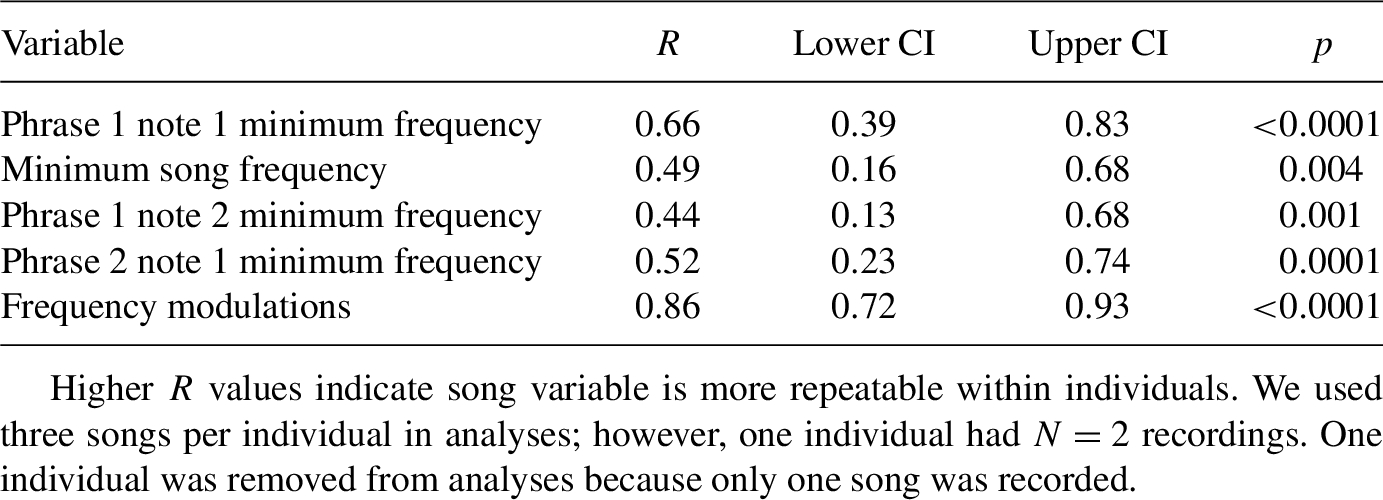
We recorded singing male southwestern willow flycatchers (from 10 m distance) in each focal territory with a Sennheiser ME66 shotgun microphone (mono-line) with Rycote handgrip and Rycote Softie windshield and a Marantz PMD661 MKII solid-state recorder (44.1 kHz sampling rate). To avoid recording the same individual twice, we only recorded individuals separated by at least 200 m. We identified an individual’s sex based on behaviour; while females are reported to sing (Seutin, 1987; Yard & Brown, 2003), males can be behaviourally identified because they advertise from conspicuous territorial perches and females sing less often and typically in response to territory intrusions close to their nest (M.A. McLeod, pers. commun.). Therefore, we restricted our recordings to songs that were unsolicited (i.e., playback was not used to elicit a song) and from individuals displaying on conspicuous perches (Mahoney et al., 2020, 2021). We then quantified frequency (Hz) and temporal song elements identified by Mahoney et al. (2020) to be variable among individuals. In that study, the authors found the following five song elements explained the most variation from a principal components analysis (PCA): minimum overall frequency; low frequencies in phrase 1 note 1, phrase 1 note 2, and phrase 2 note 1; and the minimum number of frequency modulations in the terminal portion of the song. Therefore, to test the relationship between male quality and song, we quantified the frequency and temporal song components as above (Figure 1). We digitized song recordings using Raven Pro (Cornell Lab of Ornithology) using the Hann Window (spectrogram visualization parameters: size = 256 samples, 50% window overlap, DFT = 256 samples; Mahoney et al., 2020) with an unsmoothed view. To minimize background noise, we used a high- (at 1 kHz) and low-pass (at 7.5 kHz) filter, and we only used recordings with high signal-to-noise ratios. We selected the first high signal-to-noise song in the recording, and we used one song from each individual (as in Sedgwick, 2001; Mahoney et al., 2020; Schoen et al., 2023), because intra-individual song variation in southwestern willow flycatchers is low and repeatable (Fernández-Juricic et al., 2009). To confirm songs in our study were repeatable, we quantified three songs per individual as above, and conducted a repeatability analysis using the R (R Core Development Team, 2023) package rptR (Stoffel et al., 2017). One individual from Alamo Lake had two recordings. Another individual from Seegmiller had one high signal-to-noise song recording, thus that individual was removed from the rptR analysis. We found all song variables were repeatable (Table A1 in the Appendix).
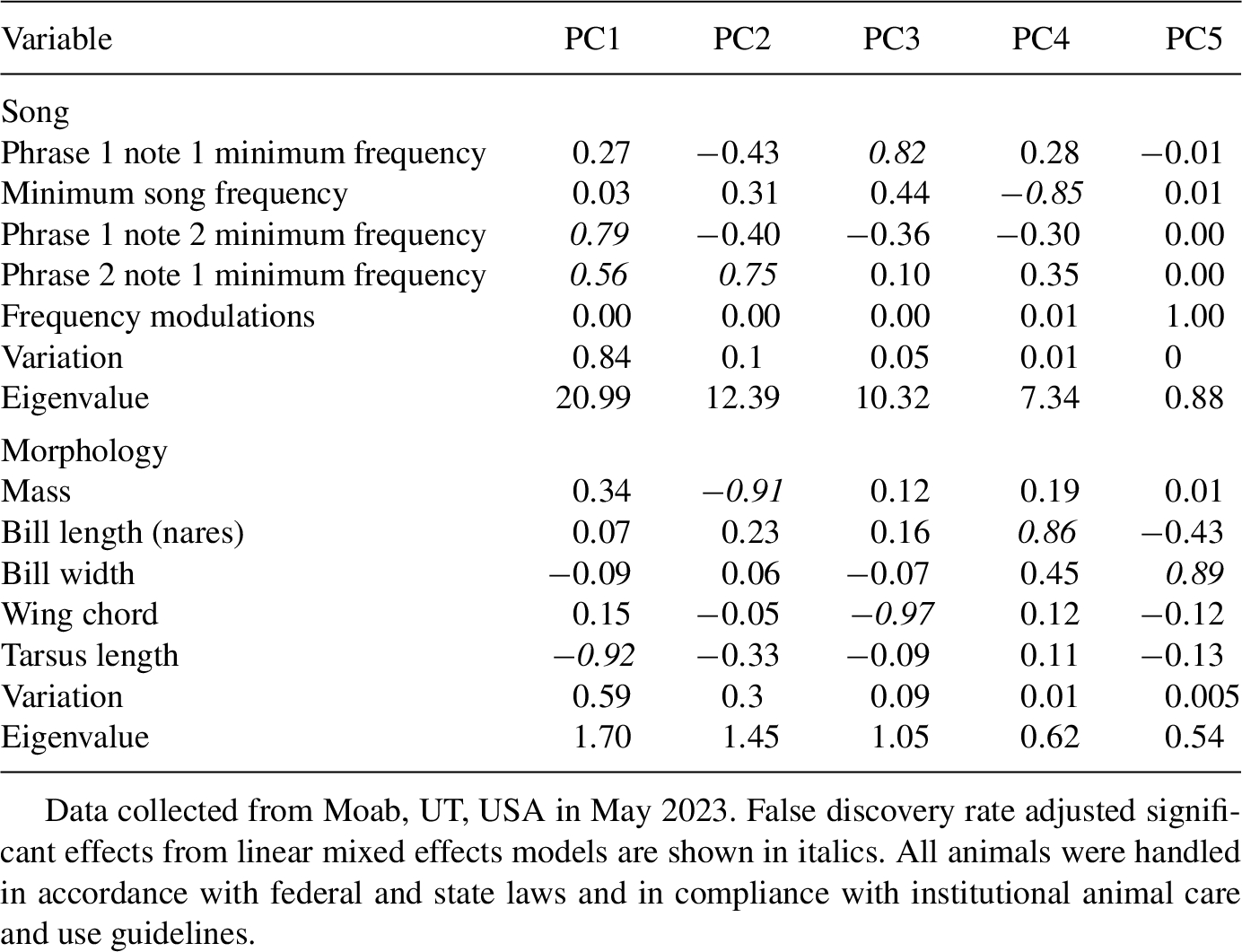
While morphology can influence song frequency and temporal elements in some Tyrannidae species (Schoen et al., 2023), our song characteristics are not corrected for body size. Willow flycatcher morphology, specifically body size and bill morphology, do not vary widely among individuals (Unitt, 1987), and we assumed morphology does not influence song characteristics. To confirm this assumption, we tested the relationship between song and morphology using an unpublished dataset of willow flycatcher song and morphology. As part of an ongoing project, we recorded songs from banded individuals near Moab, UT, USA (
2.3. Data analysis
To assess the relationship between song and apparent reproductive success in southwestern willow flycatchers, we summarized the song frequency and temporal elements from above using a PCA and selected principal components (PCs) with eigenvalues >1. We then constructed linear mixed effects models using the lme4 package in R (Bates et al., 2015) testing the relationship between the total number of fledglings as the response variable, song characteristics (summarized with PCA) and the first egg laying date (Julian day of year) as main effects. In six cases, the date of the first laid egg was unknown and we estimated the egg laying date by back calculating from the last observed phase of the nest based on the duration of each nesting phase (two days for egg laying; 12 days for incubation; 13 days for nestling; a total of 27 days to fledge) estimated from 8 years of extensive nest monitoring on the Lower Colorado River, including at our Alamo Lake site (McLeod & Pelligrini, 2011). Of the six cases with unknown first egg dates, four nests fledged young, and we estimated the first egg date by subtracting 27 days from the fledge date. In one instance, the last observed phase was incubating, so we subtracted 14 days. For the final nest, the last observed nest phase was egg laying and we subtracted two days to estimate first egg date. Similar methods have been used in our study areas to calculate first egg laying dates (Dobbs et al., 2012).
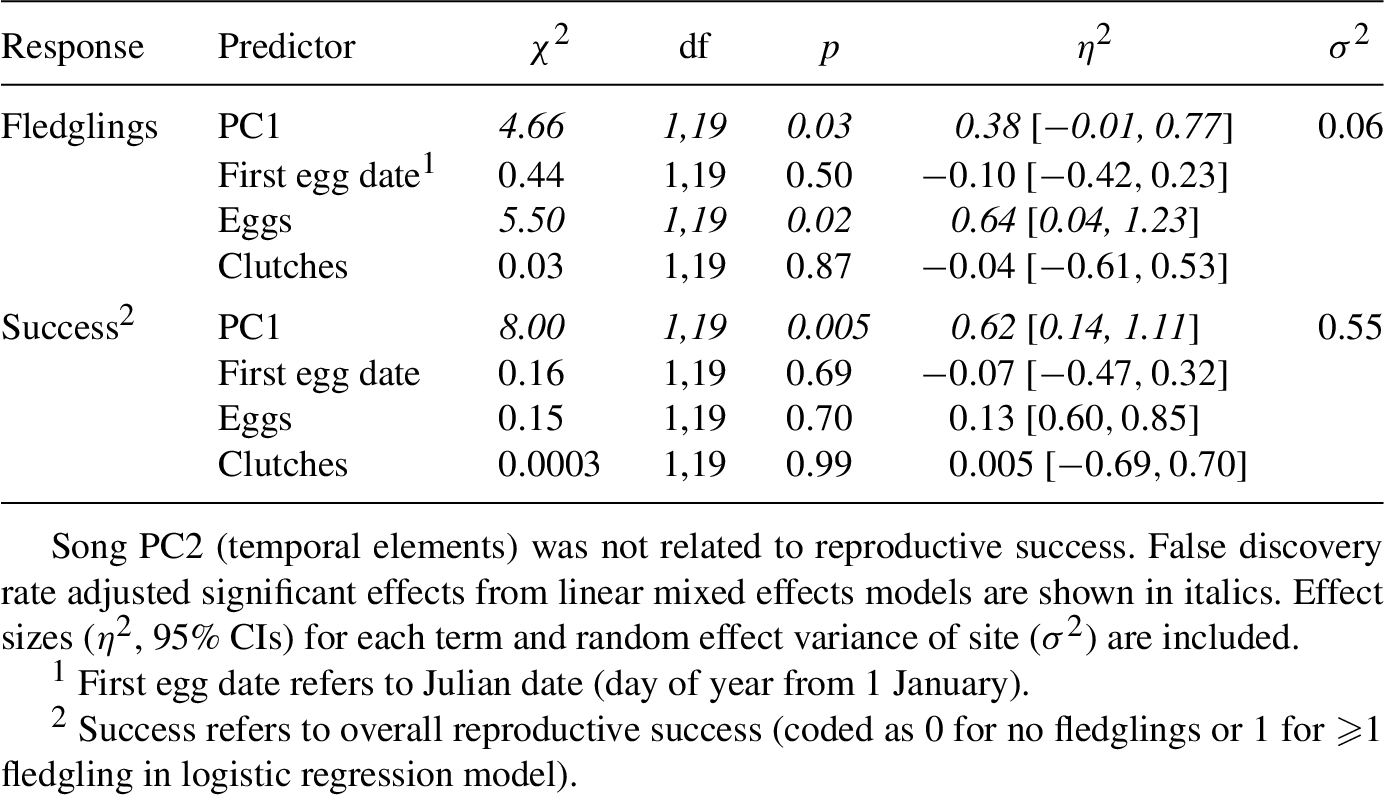
There may be a positive relationship between the number of nesting attempts and reproductive success in willow flycatchers as pairs will renest following failed attempts (Sedgwick, 2004). Indeed, we found positive relationships between the number fledglings and eggs, nestlings, and clutches (eggs:
In separate models, we tested if song characteristics (from PCA) predicted overall reproductive success (i.e. nests that fledged or failed) using a logistic regression model. Nest success was included as the response variable (coded as 0 for a nest that did not fledge young and 1 for a nest that fledged n ⩾ 1 young, see Nest Monitoring methods), and main and random effects were included as above. The number of eggs, nestlings, and fledglings did not vary annually (ANOVA
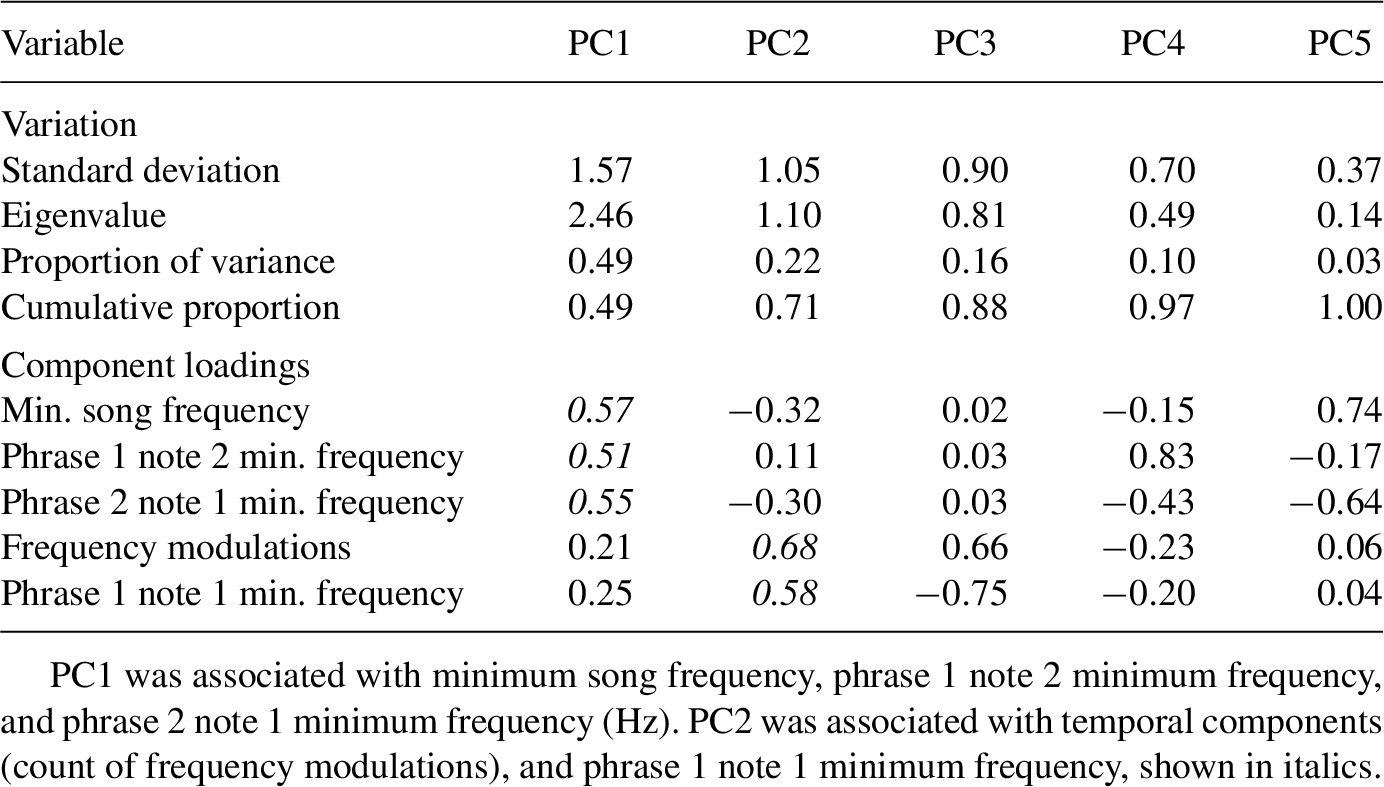
Principal components analysis results summarizing southwestern willow flycatcher (Empidonax traillii extimus) song variation.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275
2.4. Ethical notes
We conducted all field work in accordance with federal and state laws and in compliance with institutional animal care and use guidelines.
3. Results
From our song PCA, PC1 explained 49% of the variation in song structure (eigenvalue = 2.46), and song PC2 explained 22% of the total variation (eigenvalue = 1.10). PC1 and PC2 had eigenvalues >1, so we used them in subsequent analyses (Table 1). Song PC1 was positively correlated with minimum song frequency, and to a lesser extent, positively related to minimum frequency in phrase 1 note 2 and phrase 2 note 1 (Table 1). PC2 was most strongly and positively correlated with frequency modulations, but also to minimum frequencies in phrase 1 note 1 (Table 1). Therefore, higher PC1 and PC2 scores represented songs with higher minimum frequencies and with more frequency modulations in the terminal portion of the song.
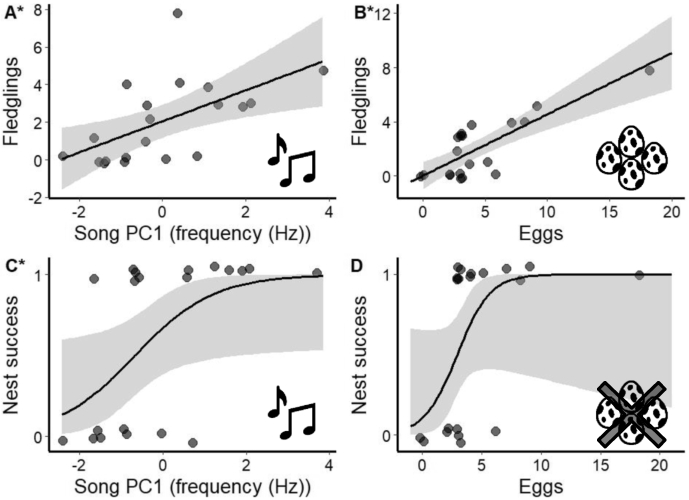
The total number of fledglings produced was positively related to song PC1 (Figure 2A;
Southwestern willow flycatchers (Empidonax traillii extimus) songs predicted the number of offspring. Individuals that (A) sang higher minimum song frequencies (Hz, summarized by PC1, 49%, eigenvalue = 2.46) and (B) had mates that laid more eggs, produced more fledglings. (C) Individuals that sang higher frequency songs also had more successful nests (fledged ⩾1 offspring); however, (D) eggs did not predict whether nests were successful. Schematics refer to dependent variables. Asterisks indicate significant effects after false discovery rate adjustments. Larger song PC1 values represent songs with higher minimum frequencies. Raw data are plotted and points are jittered to help with presentation.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275
Overall reproductive success was positively related to song PC1 (Figure 2C;
Southwestern willow flycatcher (Empidonax traillii extimus) males that sang advertisement songs with higher minimum frequencies (song PC1) produced more fledglings and had higher reproductive success, after accounting for nest initiation date, number of eggs laid, and number of clutches.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275
4. Discussion
Song is used to attract mates, and therefore is often thought to have evolved to signal quality via honest signalling (Soma & Garamszegi, 2011). Under the honest signal model of song evolution, individuals of higher quality exhibit song characteristics that are correlated with measures of quality, such as the number of young produced (Safran et al., 2013). Studies evaluating song and quality often focus on oscine songbirds, where song is learned and therefore thought to be relatively more plastic than that of suboscines, where song is innate (Raposo & Höfling, 2003). In our study, we found minimum song frequencies, a labile song characteristic among suboscine southwestern willow flycatcher individuals (Mahoney et al., 2020), predicted the apparent reproductive success of males after controlling for the first egg date, the number of eggs laid, and the number of clutches.
A logical alternative hypothesis to explain reproductive success is that pairs which lay more eggs will produce more fledglings, but there may be a trade-off between egg quantity and quality (Smith & Fretwell, 1974). Under experimentally increased clutch sizes, nests with more eggs experienced lower nest success, thus laying more eggs does not necessarily increase reproductive success (Nager et al., 2000). Indeed, we found mixed evidence that eggs correlated with reproductive success. While the number of eggs laid predicted the number of fledged offspring, when assessing if the nest was successful or not (whether a nest fledged ⩾1 young), eggs no longer predicted success, but song PC1 did. Additionally, the effect of eggs was dependent on the relatively high number of eggs from polygynous individuals, whereas the song PC1 effect was consistent regardless of whether polygynous individuals were included in the analysis.
Other aspects of suboscine song vary with individual quality including participation in dawn chorus (Sexton et al., 2007; Murphy et al., 2008) and duetting (Diniz et al., 2019), however far fewer studies found a relationship between song elements (such as frequency characteristics) and reproductive output (Araya-Ajoy et al., 2009). An important caveat of our study is that our estimates of reproductive success do not account for extra-pair paternity; therefore, reproductive success may be overestimated for some individuals and underestimated for others. Southwestern willow flycatcher extra-pair paternity rates were estimated to be 14% (range = 0.03–0.32) in a population on the Kern River, CA, USA (Pearson et al., 2006). However, polygynous males sired more young than behaviourally monogamous males, and this was not offset by differences in extra-pair paternity (Pearson et al., 2006), suggesting behavioural estimates of reproductive success, as we used, adequately capture reproductive success.
Southwestern willow flycatchers are riparian obligates, often nesting in willows (Salix spp.) and tamarisk (Tamarix spp.), and the vegetation in their environment may impact the transmission of their songs (Morton, 1975; Seddon, 2005). Individuals that produced one or more fledglings exhibited minimum song frequencies above 1526 Hz on average, and we hypothesize this song frequency transmits optimally for receiver detection within riparian environments; however, attenuation playback experiments are needed to confirm this. Alternatively, individuals with successful nests may have been singing louder to increase receiver detection, which can increase song frequency (Nemeth et al., 2013). The highest frequency song in our dataset was from the Seegmiller Marsh in St. George, UT, USA and the individual was adjacent to a road, while the lowest frequency song was from a more rural site at the Gila River near Safford, AZ, USA. It is possible that anthropogenic noise may have impacted our results. If birds were responding to variation in anthropogenic noise, it follows that average song frequencies would be highest at the more urban Seegmiller site and lowest at the rural Gila River and Alamo Lake sites. In contrast to this notion song PC1 did not vary among sites (ANOVA
In other species, song communicates territory quality to potential mates (Buchanan & Catchpole, 1997; Scales et al., 2013; Manica et al., 2014). The reasons behind this relationship include higher quality territories may hold better resources to support nesting females, the vegetation may be such that nests are more likely to be concealed from predators, and/or territory holders may be more aggressive and territorial (Phillips et al., 2020). Thus, there is selection pressure to sing unique songs so competitors can avoid costly agonistic interactions and females can easily identify individuals holding high quality territories (Phillips et al., 2020). Further, signalling territory quality may be one mechanism to prevent cheating. While signalling theory predicts that lower quality individuals should deceive potential mates and competitors by cheating (Maynard Smith, 1982), deception can result in costly territorial fights (Molles & Vehrencamp, 2001). Therefore, signals evolve to maintain honesty (Maynard Smith & Harper, 1988). Although we did not monitor arrival dates or initial pairing dates, individuals from our study singing higher frequencies songs may have held higher quality territories. The primary objective of our study was to assess the connection between flycatcher song and a direct male quality measure (i.e. number of young produced) and future studies should assess the specific information encoded in southwestern willow flycatcher song that is signalling reproductive success.
There is some evidence that singing is a physiologically demanding behaviour, and there may be a tradeoff between the energy required to sing given an animal’s morphology (Podos et al., 2004). Body mass and bill morphology can influence song characteristics in birds generally (Mikula et al., 2021), and tyrant flycatchers specifically (Schoen et al., 2023). Based on willow flycatcher individuals from Moab, UT, morphology was not related to song characteristics, however we did not have morphology for all individuals in our study with reproductive success estimates. Although it remains untested in southwestern willow flycatcher individuals, if bill and body size constrain vocal characteristics, our results suggest singing at higher minimum frequencies represents a performance challenge that is under sexual selection (Zahavi, 1975; Podos, 1997; Luther et al., 2016). Further, the interaction between bill morphology and song structure could signal foraging capabilities such as in Galapagos finches (Christensen et al., 2006). Alternatively, the higher frequency songs may be preferred by females through arbitrary mate choice (Prum, 2010). Future work should assess the role of morphology in song variation in willow flycatchers to disentangle the drivers of song evolution in this species.
Older birds may be more successful breeders, and age may be encoded in song signals (Kipper & Kiefer, 2010). A comprehensive study of song sparrows (Melospiza melodia) found repertoire size was positively correlated with the number of offspring and grand offspring; however, older individuals exhibited songs with larger repertoires (Reid et al., 2005). Similarly, older individuals can sing lower frequency songs (Garamszegi et al., 2005). Future studies could assess this hypothesis as we were not able to determine ages of individuals, but it is possible that song signals age, which can be correlated with reproductive behaviour (Kipper & Kiefer, 2010). Long term monitoring of southwestern willow flycatcher populations and their vocal behaviour will be needed to assess the relationship between age, song, and reproductive success.
Acoustic monitoring is a useful tool in conservation as it may allow researchers to remotely study key behaviours mediating the life histories of endangered animals (Teixeira et al., 2019). An on-going management challenge in the U.S. is the conservation of southwestern willow flycatcher populations (Ruegg et al., 2018). Threats to the existence of willow flycatchers in the southwest are numerous and complex but are mostly attributed to riparian habitat loss (FR, 1995; 60 FR 10694-10715). Given the results of our study, we recommend acoustic monitoring of southwestern willow flycatcher populations accompany traditional nest monitoring surveys to evaluate the reproductive success of breeding pairs.
Song is an important behaviour in birds, but the evolutionary factors involved in suboscine song variation are unresolved (Tobias et al., 2012). While it is thought that suboscine song plasticity is relatively low, there is some evidence that individual singing behaviour is an honest signal (e.g. dawn chorus participation (Murphy et al., 2008) and duetting (Diniz et al., 2019)). We offer evidence that suboscine song structure is an honest signal of reproductive success, because minimum song frequencies, which are variable among individuals, predicted the number of fledglings. While the specific mechanism remains to be studied, our study offers unique evidence of the role of honest signalling in suboscine song evolution.
Corresponding author’s e-mail address: seanmahoney@arizona.edu
Acknowledgements
After the first author, names appear alphabetically. We thank T.C. Theimer and B. Pasch for reviewing early drafts of the manuscript. A. Songer assisted in collecting morphology data. Comments from two anonymous reviewers and E. Clotfelter greatly improved the paper. Finally, preliminary results were presented at several RiversEdge West workshops and discussions were helpful in the development of the paper.
References
Andersson, M. (1994). Sexual selection. — Princeton University Press, Princeton, NJ.
Araya-Ajoy, Y.M., Chaves-Campos, J., Kalko, E.K. & DeWoody, J.A. (2009). High-pitched notes during vocal contests signal genetic diversity in ocellated antbirds. — PLoS ONE 4: e8137.
Baker, M.C., Bjerke, T.K., Lampe, H. & Espmark, Y. (1986). Sexual response of female great tits to variation in size of males’ song repertoires. — Am. Nat. 128: 491-498.
Bates, D., Maechler, M., Bolker, B., Walker, S., Christensen, R.H.B., Singmann, H., Dai, B., Grothendieck, G., Green, P. & Bolker, M.B. (2015). Package ‘lme4’. — Convergence 12: 2.
Ben-Shachar, M.S., Lüdecke, D. & Makowski, D. (2020). Effectsize: Estimation of effect size indices and standardized parameters. — J. Open Source Softw. 5: 2815.
Bruni, A. & Foote, J.R. (2014). Dawn singing of Eastern phoebes varies with breeding stage and brood number. — Wilson J. Ornithol. 126: 500-507.
Brunton, D.H., Roper, M.M. & Harmer, A.M. (2016). Female song rate and structure predict reproductive success in a socially monogamous bird. — Front. Ecol. Evol. 4: 13.
Buchanan, K.L. & Catchpole, C.K. (1997). Female choice in the sedge warbler Acrocephalus schoenobaenus multiple cues from song and territory quality. — Proc. Roy. Soc. Lond. B: Biol. Sci. 264: 521-526.
Byers, B.E. & Kroodsma, D.E. (2009). Female mate choice and songbird song repertoires. — Anim. Behav. 77: 13-22.
Christensen, R., Kleindorfer, S. & Robertson, J. (2006). Song is a reliable signal of bill morphology in Darwin’s small tree finch Camarhynchus parvulus, and vocal performance predicts male pairing success. — J. Avian Biol. 37: 617-624.
Collins, S. (2004). Vocal fighting and flirting: the functions of birdsong. — In: Nature’s music: the science of birdsong (Marler, P. & Slabbekoorn, H., eds). Elsevier Academic Press, San Diego, CA, p. 39-79.
Diniz, P., Macedo, R.H. & Webster, M.S. (2019). Duetting correlates with territory quality and reproductive success in a suboscine bird with low extra-pair paternity. — Auk: Ornithol. Adv. 136: uky004.
Dobbs, R.C., Huizinga, M., Edwards, C.N. & Fridell, R.A. (2012). Status, reproductive success, and habitat use of Southwestern willow flycatchers on the Virgin River, Utah, 2008–2011. — Publication Number 12-36, Utah Division of Wildlife Resources, Salt Lake City, UT.
English, H.C., Graber, A.E., Stump, S.D., Telle, H.E. & Ellis, L.A. (2006). Southwestern willow flycatcher 2005 survey and nest monitoring report. — Nongame and endangered wildlife program technical report 248, Arizona Game and Fish Department, Phoenix, AZ.
Fernández-Juricic, E., del Nevo, A.J. & Poston, R. (2009). Identification of individual and population-level variation in vocalizations of the endangered Southwestern willow flycatcher (Empidonax traillii extimus). — Auk 126: 89-99.
Fox, J., Weisberg, S., Adler, D., Bates, D., Baud-Bovy, G., Ellison, S. & Heiberger, R. (2012). Package ‘car’. — R Foundation for Statistical Computing, Vienna.
FR (1995). — SWFL listing. Available online at https://www.govinfo.gov/content/pkg/FR-1995-02-27/pdf/95-4531.pdf.
Garamszegi, L.Z. (2005). Bird song and parasites. — Behav. Ecol. Sociobiol. 59: 167-180.
Garamszegi, L.Z. & Møller, A.P. (2004). Extrapair paternity and the evolution of bird song. — Behav. Ecol. 15: 508-519.
Garamszegi, L.Z., Heylen, D., Møller, A.P., Eens, M. & de Lope, F. (2005). Age-dependent health status and song characteristics in the barn swallow. — Behav. Ecol. 16: 580-591.
Gentry, K.E., McKenna, M.F. & Luther, D.A. (2018). Evidence of suboscine song plasticity in response to traffic noise fluctuations and temporary road closures. — Bioacoustics 27: 165-181.
Johnstone, R.A. (1995). Honest advertisement of multiple qualities using multiple signals. — J. Theor. Biol. 177: 87-94.
Kipper, S. & Kiefer, S. (2010). Age-related changes in birds’ singing styles: on fresh tunes and fading voices? — In: Advances in the Study of Behavior (Slater, P.J.B., Snowdon, C.T., Rosenblatt, J.S. & Milinski, M., eds). Academic Press, Cambridge, MA, p. 77-118.
Kokko, H., Brooks, R., McNamara, J.M. & Houston, A.I. (2002). The sexual selection continuum. — Proc. Roy Soc. Lond. B: Biol. Sci. 269: 1331-1340.
Kokko, H., Brooks, R., Jennions, M.D. & Morley, J. (2003). The evolution of mate choice and mating biases. — Proc. Roy Soc. Lond. B: Biol. Sci. 270: 653-664.
Koronkiewicz, T.J., Sogge, M.K., Van Riper III, C. & Paxton, E.H. (2006). Territoriality, site fidelity, and survivorship of willow flycatchers wintering in Costa Rica. — Condor 108: 558-570.
Kroodsma, D.E. (1984). Songs of the alder flycatcher (Empidonax alnorum) and willow flycatcher (Empidonax traillii) are innate. — Auk 101: 13-24.
Luther, D.A., Phillips, J. & Derryberry, E.P. (2016). Not so sexy in the city: urban birds adjust songs to noise but compromise vocal performance. — Behav. Ecol. 27: 332-340.
Mahoney, S.M., Reudink, M.W., Pasch, B. & Theimer, T.C. (2020). Song but not plumage varies geographically among willow flycatcher Empidonax traillii subspecies. — J. Avian Biol. 51: jav.02621.
Mahoney, S.M., Pasch, B. & Theimer, T.C. (2021). Subspecies discrimination based on song structure by willow flycatchers. — J. Field Ornithol. 92: 173-183. DOI:10.1111/jofo.12366.
Manica, L.T., Maia, R., Dias, A., Podos, J. & Macedo, R.H. (2014). Vocal output predicts territory quality in a Neotropical songbird. — Behav. Proc. 109: 21-26.
Marten, K. & Marler, P. (1977). Sound transmission and its significance for animal vocalization: II. Tropical forest habitats. — Behav. Ecol. Sociobiol. 2: 291-302.
Marten, K., Quine, D. & Marler, P. (1977). Sound transmission and its significance for animal vocalization: I. Temperate habitats. — Behav. Ecol. Sociobiol. 2: 271-290.
Maynard Smith, J. (1982). Do animals convey information about their intentions? — J. Theor. Biol. 97: 1-5.
Maynard Smith, J. & Harper, D.G.C. (1988). The evolution of aggression: can selection generate variability? — Phil. Trans. Roy. Soc. Lond. B: Biol. Sci. 319: 557-570.
McGregor, P.K., Krebs, J.R. & Perrins, C.M. (1981). Song repertoires and lifetime reproductive success in the great tit (Parus major). — Am. Nat. 118: 149-159.
McLeod, M.A. & Pellegrini, A.R. (2011). Southwestern willow flycatcher surveys, demography, and ecology along the lower Colorado River and tributaries, 2010. — Annual report submitted to U.S. Bureau of Reclamation, Boulder City, NV, by SWCA Environmental Consultants, Flagstaff, AZ.
Mikula, P., Valcu, M., Brumm, H., Bulla, M., Forstmeier, W., Petrusková, T., Kempenaers, B. & Albrecht, T. (2021). A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. — Ecol. Lett. 24: 477-486.
Molles, L.E. & Vehrencamp, S.L. (2001). Songbird cheaters pay a retaliation cost: evidence for auditory conventional signals. — Phil. Trans. Roy. Soc. Lond. B: Biol. Sci. 268: 2013-2019.
Morton, E.S. (1975). Ecological sources of selection on avian sounds. — Am. Nat. 109: 17-34.
Murphy, M.T., Sexton, K., Dolan, A.C. & Redmond, L.J. (2008). Dawn song of the eastern kingbird: an honest signal of male quality? — Anim. Behav. 75: 1075-1084.
Nager, R.G., Monaghan, P. & Houston, D.C. (2000). Within-clutch trade-offs between the number and quality of eggs: experimental manipulations in gulls. — Ecology 81: 1339-1350.
Nemeth, E., Pieretti, N., Zollinger, S.A., Geberzahn, N., Partecke, J., Miranda, A.C. & Brumm, H. (2013). Bird song and anthropogenic noise: vocal constraints may explain why birds sing higher-frequency songs in cities. — Proc. Roy. Soc. Lond. B: Biol. Sci. 280: 20122798.
Paxton, E.H., Sogge, M.K., McCarthey, T.D. & Keim, P. (2002). Nestling sex ratio in the Southwestern willow flycatcher. — Condor 104: 877-881.
Paxton, E.H., Sogge, M.K., Durst, S.L., Theimer, T.C. & Hatten, J.R. (2007). The ecology of the southwestern willow flycatcher in central Arizona — a 10-year synthesis report. — Open-File Repirt 2007-1381, U.S. Geological Survey, Reston, VA.
Pearson, T., Whitfield, M.J., Theimer, T.C. & Keim, P. (2006). Polygyny and extra-pair paternity in a population of Southwestern willow flycatchers. — Condor 108: 571-578.
Phillips, J.N., Cooper, W.J., Luther, D.A. & Derryberry, E.P. (2020). Territory quality predicts avian vocal performance across an urban–rural gradient. — Front. Ecol. Evol. 8: 587120.
Podos, J. (1997). A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes: Emberizidae). — Evolution 51: 537-551.
Podos, J., Huber, S.K. & Taft, B. (2004). Bird song: the interface of evolution and mechanism. — Annu. Rev. Ecol. Evol. Syst. 35: 55-87.
Prum, R.O. (2010). The Lande–Kirkpatrick mechanism is the null model of evolution by intersexual selection: implications for meaning, honesty, and design in intersexual signals. — Evolution 64: 3085-3100.
R Core Development Team (2023). RA language and environment for statistical computing. — R Foundation for Statistical Computing, Vienna.
Raposo, M.A. & Höfling, E. (2003). Overestimation of vocal characters in Suboscine taxonomy (Aves: Passeriformes: Tyranni): causes and implications. — Lundiana 4: 35-42.
Reid, J.M., Arcese, P., Cassidy, A.L., Hiebert, S.M., Smith, J.N., Stoddard, P.K., Marr, A.B. & Keller, L.F. (2005). Fitness correlates of song repertoire size in free-living song sparrows (Melospiza melodia). — Am. Nat. 165: 299-310.
Rourke, J.W., McCarthey, T.D., Davidson, R.F. & Santaniello, A.M. (1999). Southwestern willow flycatcher nest monitoring protocol. — Nongame and Endangered Wildlife Program, Technical Report 144, AGFD, Phoenix, AZ.
Ruegg, K., Bay, R.A., Anderson, E.C., Saracco, J.F., Harrigan, R.J., Whitfield, M., Paxton, E.H. & Smith, T.B. (2018). Ecological genomics predicts climate vulnerability in an endangered southwestern songbird. — Ecol. Lett. 21: 1085-1096.
Safran, R.J., Scordato, E.S., Symes, L.B., Rodríguez, R.L. & Mendelson, T.C. (2013). Contributions of natural and sexual selection to the evolution of premating reproductive isolation: a research agenda. — Trends Ecol. Evol. 28: 643-650.
Scales, J., Hyman, J. & Hughes, M. (2013). Fortune favours the aggressive: territory quality and behavioural syndromes in song sparrows, Melospiza melodia. — Anim. Behav. 85: 441-451.
Schoen, J., Miller, S.M., Jennings, Q., Reudink, M.W. & Mahoney, S.M. (2023). Disentangling the mechanisms of signal evolution in Tyrannidae flycatchers Part I: Song is constrained by morphology and covaries with ecological factors. — J. Field Ornithol. 94: 6.
Seddon, N. (2005). Ecological adaptation and species recognition drives vocal evolution in neotropical suboscine birds. — Evolution 59: 200-215.
Sedgwick, J.A. (2001). Geographic variation in the song of willow flycatchers: differentiation between Empidonax traillii adastus and E. t. extimus. — Auk 118: 366-379.
Sedgwick, J.A. (2004). Site fidelity, territory fidelity, and natal philopatry in willow flycatchers (Empidonax traillii). — Auk 121: 1103-1121.
Seutin, G. (1987). Female song in Willow Flycatchers. — Auk 104: 329-330.
Sexton, K., Murphy, M., Redmond, L. & Dolan, A. (2007). Dawn song of eastern kingbirds: intrapopulation variability and sociobiological correlates. — Behaviour 144: 1273-1295.
Smith, C.C. & Fretwell, S.D. (1974). The optimal balance between size and number of offspring. — Am. Nat. 108: 499-506.
Sogge, M.K., Marshall, R.M., Sferra, S.J. & Tibbitts, T.J. (1997). A southwestern willow flycatcher natural history summary and survey protocol. — Technical Report NPS/NAUCPRS/NRTR-97/12, USGS Colorado Plateau Research Station, Northern Arizona University, Flagstaff, AZ.
Soma, M. & Garamszegi, L.Z. (2011). Rethinking birdsong evolution: meta-analysis of the relationship between song complexity and reproductive success. — Behav. Ecol. 22: 363-371.
Stoffel, M.A., Nakagawa, S. & Schielzeth, H. (2017). rptR: Repeatability estimation and variance decomposition by generalized linear mixed-effects models. — Methods Ecol. Evol. 8: 1639-1644.
Teixeira, D., Maron, M. & van Rensburg, B.J. (2019). Bioacoustic monitoring of animal vocal behavior for conservation. — Conserv. Sci. Pract. 1: e72.
Tobias, J.A., Brawn, J.D., Brumfield, R., Derryberry, E.P., Kirschel, A.N. & Seddon, N. (2012). The importance of suboscine birds as study systems in ecology and evolution. — Ornithol. Neot. 23: 261-274.
Unitt, P. (1987). Empidonax traillii extimus: an endangered subspecies. — Western Birds 18: 137-162.
Woodgate, J.L., Mariette, M.M., Bennett, A.T., Griffith, S.C. & Buchanan, K.L. (2012). Male song structure predicts reproductive success in a wild zebra finch population. — Anim. Behav. 83: 773-781.
Yard, H.K. & Brown, B.T. (2003). Singing behavior of Southwestern Willow Flycatchers. — Stud. Avian Biol. 26: 125-130.
Zahavi, A. (1975). Mate selection — A selection for a handicap. — J. Theor. Biol. 53: 205-214.
Appendix
Southwestern willow flycatcher songs are significantly repeatable, based on repeatability analyses.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275
Principal components analysis loadings of willow flycatcher (Empidonax traillii) song from banded individuals with morphology measurements.
Citation: Behaviour 161, 7 (2024) ; 10.1163/1568539X-bja10275


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}