Abstract
Reptiles are a vastly understudied taxon, leaving large gaps in comparative cognition and behaviour. They are more similar, cognitively, to mammals and birds than previously thought, but are difficult to test in cognitive tasks. The difficulty comes in effectively motivating reptiles to perform behaviours that allow for cognitive and behavioural comparison to mammals and birds. Painted turtles are a good model for comparison due to their relatively common use in scientific studies, compared to other reptiles, allowing them to be widely compared with mammals and birds. Methyl anthranilate (MA) is a potential short-term aversive stimulus in reptiles, similar to capsaicin in mammals, that could aid in motivating reptiles to perform cognitive tasks for comparison with mammals and birds. In this study, we tested MA on painted turtles to determine if it is an effective short-term aversive stimulus for use in behavioural and cognitive studies. We found that MA produces a short-term aversive reaction in painted turtles, exemplified by a decline in willingness to consume food treated with MA. Future comparative studies could benefit from using MA as an aversive stimulus in cognitive and behavioural tests on reptiles.
1. Introduction
In the study of evolutionary cognition, the comparative approach is a powerful tool to examine patterns of behaviour across taxa, and to provide insight into the potential processes that drive the evolution of traits (Miles & Dunham, 1993; Huey et al., 2019). To maintain strong validity in comparative studies, a variety of taxa must be considered, especially those previously underrepresented. However, comparing traits across a diverse array of taxa can be difficult, as it requires balancing consistency of testing with accounting for diverse taxa’s unique evolutionary background, selection pressures, and taxon-specific gestalt (Burghardt, 1977; Shaw & Schmelz, 2017).
Reptiles, a historically understudied and underrepresented taxa, are being increasingly included in comparative cognition studies as a root taxonomic group for comparisons between mammals and birds, leading to additions to our understanding of the evolution of cognition in vertebrates (Roth et al., 2019; Szabo et al., 2021; Kundey, 2024). For example, we know that reptiles are capable of learning and performing a number of complex cognitive tasks (Burghardt, 1977; Powers et al., 2009; Burghardt, 2020), and even demonstrate similar spatial learning and memory capabilities to mammals and birds (Roth & Krochmal, 2016; Kundey, 2024). The overlaps in cognitive capabilities are likely due to their functionally analogous brain regions used in spatial learning (López et al., 2000; López et al., 2003). Taxa with analogous brain regions are likely to possess similar cognitive mechanisms and thus similar learning abilities (Fischer et al., 2019). Differences in neural mechanisms underlying cognition between endotherms and ectotherms have also been identified, highlighting the evolutionary progress of certain cognitive systems (Powers et al., 2009). For example, scopolamine seems to affect reptiles’ and mammals’ negative patterning learning similarly, implying an ancestral origin of the effect (Powers et al., 2009). Thus, we are continuing to expand our understanding of the neural mechanisms underlying cognition in amniotic vertebrates and the factors that may be responsible for driving evolutionary differences and similarities (Roth et al., 2019). In doing so, comparing the cognitive abilities of these taxa could reveal even more new information on the evolution of learning abilities and cognition (Roth et al., 2019; De Meester & Baeckens, 2021; Szabo et al., 2021; Roth & Krochmal, 2024).
Despite the benefits of studying diverse taxa from an integrative standpoint, reptiles remain an underrepresented taxon in much of cognitive science (Matsubara et al., 2017; De Meester & Baeckens, 2021; Kundey, 2024). A key limiting factor in comparing the cognition of mammals and birds with reptiles is the nature of motivating an animal to perform a task (Dayan & Balleine, 2002; Yee & Braver, 2018). When an individual is found not to perform a certain task, determining whether it is unable to perform the task or whether it lacks the proper motivation to do so is challenging (van Horick & Madden, 2016). This challenge becomes particularly difficult when comparing species with different umwelts, or worldviews (Varholick, 2022). Designing a task that adheres to both species’ ecological contexts and species-specific motivations while remaining scientifically reliable and relevant can be impossible in some cases. Thus, when species have different motivations, ensuring control, reliability, and effective comparison within the context of an experiment becomes difficult (Burghardt, 1977).
To test hypotheses about the factors that influence behavioural or cognitive processes, researchers have at their disposal a wide variety of both positive and negative stimuli to motivate animals to complete certain tasks or perform certain behaviours (Dayan & Balleine, 2002). Positive stimuli are rewards that encourage the performance of certain tasks or behaviour (positive reinforcement). Negative stimuli, on the other hand, discourage certain tasks or behaviours through punishment (Michael, 1975; Dayan & Balleine, 2002; Kuczaj & Xitgo, 2002). Implementing a punishment to discourage an undesired behaviour involves presenting an aversive stimulus after the undesired behaviour is performed, thereby encouraging the individual to display a different, desired behaviour (Wergård et al., 2015).
Both positive and negative stimuli are often tied to homeostatic functions, such as accessing food or water, sleep, avoiding pain or potential predators, or thermoregulation (Toates, 1995; Mota-Rojas et al., 2021). Birds and mammals are easy to motivate using access to food and water as positive reinforcement. These taxa, in part due to their high metabolisms that require them to eat frequently (McNab, 2019; Alarcón, 2022), become more food-driven and spend more time searching for food sources and learning to complete tasks resulting in access to food when food deprived prior to testing (Kuczaj & Xitco, 2002; Padmala & Pessoa, 2010; Anselme & Güntürkün, 2019; Szabo et al., 2022). Negative stimuli and punishments often utilize aversive experiences, such as pain, to reduce unwanted behaviours. Different motivators can be used as positive and negative reinforcement, and punishment as ways to motivate animals to learn certain behaviours and tasks.
While birds and mammals can be motivated using relatively simple methods and tools, the same cannot be said for most reptiles (Davis & Burghardt, 2011; Wilkinson & Glass, 2018; Roth et al., 2024). Encouraging reptiles to exhibit a desired behaviour to gain access to previously withheld resources does not ensure they will be motivated enough to change their behaviour in the desired way (DeNardo, 2021). As ectotherms, reptiles have slow metabolisms and need to feed relatively infrequently. Thus, with some exceptions (i.e., tortoises, which can be quite food driven; Tomonaga et al., 2023), reptiles need to be food deprived for an extended period of time to be motivated enough to learn and complete a task for food, which is impractical for testing (Legendre & Davesne, 2020; Tomonaga et al., 2023). While motivating reptiles with high-reward food is possible, doing so typically requires many trials, and may not be successful in all individuals, making it difficult or impractical for studies with small sample sizes. While food can be used to motivate turtles as a positive reinforcer (e.g., Powers et al., 2009), motivating behaviour with aversive stimuli can often be faster and lead to more efficient learning (van der Schaaf et al., 2022).
One classic negative reinforcer used in mammal and bird studies is thermal stress. However, this approach is problematic for reptiles. As ectotherms, reptiles respond to low temperatures by conserving energy and reducing activity rather than increasing behaviour (Prassack et al., 2001; Krochmal & Bakken, 2003; Tattersall, 2016; Pettersen, 2020; Bodensteiner et al., 2021; Giacometti et al., 2021; Bernich et al., 2022), and high temperatures by relocating to a cooler location. While extreme heat successfully motivated rattlesnakes to complete learning tasks in order to locate cooler temperatures, it failed with pit vipers and required specialized apparatus (Krochmal et al., 2018) that could be impractical and complex. This strategy would likely prove equally ineffective for semi-aquatic reptiles like turtles, which physiologically adapt to abnormal water temperatures (Prossack et al., 2001; Pettersen, 2020; Mednikova & Kalabushev, 2023).
While motivators can vary between species, semi-aquatic turtles are in alignment with most other reptiles, with temperature and food being ineffective and unreliable positive stimuli (Krochmal et al., 2018; Legendre & Davesne, 2020). Given these limitations, an alternative means of motivating reptiles in a controlled experimental context that is more practical than food as a positive stimulus or thermal manipulation is needed. An aversive stimulus that motivates semi-aquatic reptiles, such as painted turtles (Chrysemys picta), could also motivate a wide breadth of reptiles (Roth et al., 2019; Szabo et al., 2021), leading to increased comparison and knowledge of the cognitive and behavioural abilities of reptiles (Kundey, 2024). Painted turtles are a fitting model for studying aversive stimuli because they are motivationally in alignment with other reptiles, and they are a commonly studied species in comparative cognition, including at the cognitive and behavioural levels, relative to other reptiles (Valenzuela, 2009; De Meester & Baeckens, 2021; Roth et al., 2021).
We propose methyl anthranilate (MA) as a potential novel aversive stimulus for behavioural manipulation and motivation in reptilian studies. Access to a simple aversive stimulus, as opposed to an appetitive stimulus, for behavioural studies could lead to greater cross-species comparison, and increased clarity on the evolution of reptilian cognition. MA is a chemical naturally found in Concord grapes and artificially made as a petroleum-based compound derived from anthranilic acid. It is used as artificial grape flavouring and scent, and is a common bird repellent for crop protection, and a motivator in a variety of avian studies (Werner & Avery, 2017; Krishnatreyya et al., 2018; Luo et al., 2019). MA activates Transient Receptor Potential Ankyrin 1(TRPA1) receptors in birds, causing a negative sensation and immediate aversive effect, making it an efficient bird repellent (Werner & Avery, 2017; Zhang et al., 2022), sometimes teaching birds to avoid the novel stimulus after one exposure (Clark et al., 2014b; Ahmad et al., 2016; Werner et al., 2017; Zhang et al., 2022; Chamorro et al., 2023; DeLiberto & Werner, 2024). MA is a non-invasive, low-impact aversive stimulus functionally equivalent to capsaicin in mammals — both produce short-lived negative sensations causing no tissue damage (Krishnatreyya et al., 2018), with capsaicin activating TRPV1 receptors that are functionally equivalent to avian TRPA1 receptors (Fernandes et al., 2012). It is possible that the TRPA1 receptor and its response to MA is a conserved trait among avian and non-avian reptiles (Saito et al., 2012; Saito & Tominaga, 2017; Zhong et al., 2017; Zhang et al., 2022; Saito & Saito, 2025), as both birds and reptiles respond similarly to some bitter substances that seem to activate TRPA1, meaning they spit out the aversive food and show less interest in consuming it (Rowland et al., 2015; Saito & Tominaga, 2017; Griffen et al., 2022). Because of these shared receptors, reptiles may react to MA similarly to birds.
Thus, we tested the hypothesis that Eastern painted turtles (Chrysemys picta) have a short-lived, aversive response to MA, such that it could be potentially used as a negative stimulus in future behavioural experiments. We predicted that if MA is perceived as aversive, we would observe an increase in distaste-related behaviours and behaviours of avoidance, such as an increased reluctance to fully consume the food, an increased number of spit outs, and consequently, an increased number of bites and strikes, as the turtles repeatedly bite and spit out the same piece of cod (Moldowan et al., 2015).
2. Methods
2.1. Species description
We tested 16 adult, long-term captive eastern painted turtles (Chrysemys picta). Fourteen of these turtles were wild-caught (7 originated from Chesapeake Farms in Kent County, MD; 7 from Chester River Field Station in Queen Anne’s County, MD, USA). The remaining two turtles were purchased from a commercial supplier (ReptileCity, Lexington, SC, USA). All turtles used in testing have lived in the Franklin and Marshall College Vivarium since 2013. All wild-caught turtles were held in two 1817 l artificial ponds, while the captive-raised turtles were held in two 37.8 l tanks. Before the experiment, all of the turtles were fed ReptoMin turtle pellets daily and fresh vegetables (e.g., lettuce and squash) 3–4 times per week. The turtles were allowed to freely graze on the food. Beginning six months prior to, and continuing throughout testing, we withheld all foods other than ReptoMin turtle pellets three times per week. We allowed the turtles to eat to satiation, but removed any leftover food after 30 min to prevent the turtles from grazing and to maintain cleanliness.
2.2. Experimental design
We habituated the turtles to the testing apparatus (described below) by running them in one 10-min habituation trial every day for 12 days. In order to meet the criterion for habituation, the turtles needed to eat the presented cod during any seven out of eight consecutive habituation trials, over the course of the 12 habituation trials. If a turtle did not meet criterion, they were not included in the study. If a turtle met the criterion sooner than day 12, we tested them the day after reaching criterion, and conducted the final control trial the day after testing, following an ABA design, in which a baseline test is run (A), followed by the treatment (B), and finally, another baseline (control) test (A). No additional tests were conducted after the final control test. We tested each turtle a maximum of once per day.
2.3. Details of testing apparatus
We conducted all trials in a 40.64 cm × 20.32 cm × 25.4 cm testing apparatus filled with 12–15 cm of water, which was enough for the turtles to be fully submerged and comfortably swim around. The apparatus was opaque to prevent turtles from seeing into the testing room. We placed a GoPro Hero 5 Black on a tripod approximately 40 cm directly above the testing apparatus to film all trials for coding and analysis.
2.4. Details of test/procedure
Each turtle was tested individually in a semi-arbitrary order. The turtle was placed in the testing apparatus and allowed 10 min of habituation prior to feeding to reduce handling stress. After 10 min, we placed a 2 cm3 piece of thawed cod (obtained from a local grocery store) approximately eight cm in front of the turtle. The turtles were given 10 min to eat the cod, after which, we returned them to their enclosures and removed any remaining cod from the apparatus before beginning the next turtle’s trial. Control/habituation cod was soaked in 200-proof ethanol (solvent for the treatment stimulus) for 10 min. The cod was then dried at room temperature for 15 min to allow the ethanol to evaporate entirely before being fed to the turtles. Once a turtle reached criterion (eating seven of eight consecutive days), the following day they were tested following the same procedure, but using the treatment cod. Treatment cod was soaked in MA (food grade; Sigma-Aldrich) dissolved in 75% (v/v) ethanol for 10 min, allowing the cod to dry and the alcohol to fully evaporate as before. The day following the treatment test, we ran the turtles in a final control trial using non-MA treated cod. We tested any turtle receiving treatment cod last in the day to prevent potentially contaminating the water for turtles not receiving treatment cod that day.
2.5. Video coding
All habituation and testing videos were filmed and coded following the same guidelines. All trials were recorded and scored by two independent reviewers (TRG and TCR) with high interobserver reliability (Cronbach’s Alpha = 0.974). We measured latency to first bite as the time from when the cod hit the bottom of the testing apparatus and the turtles’ heads were <90° to the fish (to ensure the turtle had seen the fish), to when the turtle took the first bite of cod. A strike was defined as when the turtle moved its head in and out of its shell quickly, while attempting to bite the cod. Strikes included any attempt to eat the cod, including ripping apart the fish without consuming any of it, using the front legs to rip apart the cod while it was being consumed, or the turtle rapidly biting at the cod without necessarily consuming anything. A spit out was when a turtle bit a piece of cod and then entirely ejected the cod from its mouth, without taking another bite, and when a turtle used its front legs to push cod out of its mouth without actually consuming any fish. We determined how long it took a turtle to finish the cod by measuring the time from the first bite to the time when all cod had been consumed, excluding small, incidental flakes that came off of the fish. We also recorded whether a turtle did not touch the cod at all, and if they finished eating the entire piece of cod.
2.6. Data analysis
We conducted a Friedman’s test to compare the turtles’ performance across trials (Habituation, Treatment, Control), specifically looking at the behaviours indicating aversion to the MA (number of strikes/bites, and spit outs, and the latencies to first bite and to finishing the cod). Four of the 16 turtles did not meet the habituation criterion and thus were not used in testing. All 12 turtles tested consumed fish in the habituation and post-treatment control tests. However, two turtles did not attempt to bite the fish at all during the treatment and were thus removed from the analyses of the number of strikes/bites and the number of spit-outs. We analysed the ten turtles that at least attempted to bite the fish for the number of strikes/bites and the number of spit-outs. We used a Cochran’s Q to compare the proportion of turtles that ate all of the cod to those who did not eat at all. We included all 12 turtles that were tested in the Cochran’s Q analysis. Pairwise analyses were conducted comparing results between Habituation and Control trials for all measures. All figures were made in SPSS.
3. Results
We hypothesized that, due to the shared evolutionary history between avian and non-avian reptilian species (Roth et al., 2019), as well as superficial similarities of TRPA1 between those groups (Rowland et al., 2015), painted turtles would react negatively to MA, indicating that it is an aversive stimulus. We predicted that the presence of MA in a desired food item would be consistent with behaviours of avoidance, such as an increased reluctance to fully consume the food, an increased number of spit outs, and consequently, an increased number of bites and strikes, as the turtles repeatedly bit and spat out the same piece of cod.
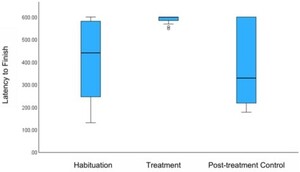
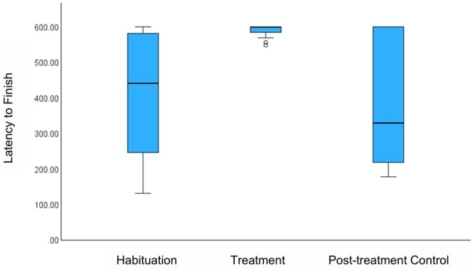
Consistent with our hypotheses, the painted turtles seemed to perceive MA as aversive, as evidenced by their reactions to consuming the treated cod, which largely supported our predictions. While the turtles took similar amounts of time to take the initial bite of the control and MA-treated cods (

The mean difference in latency, in seconds, between trials to finishing the cod. Error bars represent +/− 1 SE. There was a significant difference in the latency to finish the Control versus Treatment cod, using Friedman’s test (
Citation: Behaviour 163, 5 (2026) ; 10.1163/1568539X-bja10355
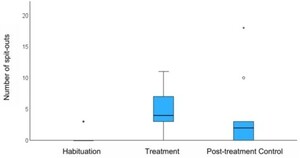
While eating, the turtles rejected (spat out) the MA-treated cod more frequently than the control, during either Habituation or Control trials. There were differences in the number of spit outs between all three trials (
The mean difference between trials in the number of spit outs. Error bars represent +/− 1 SE,
Citation: Behaviour 163, 5 (2026) ; 10.1163/1568539X-bja10355
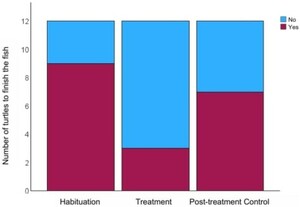
Importantly, several turtles did not finish the MA-treated cod (
The number of turtles to finish the cod during each trial. The legend is distinguishing whether the turtles consumed all of the cod they were presented with. Red represents the turtles that finished, Blue represents the turtles that did not finish all the cod. There was a significant difference in the number of turtles to finish the Control versus the Treatment cod (
Citation: Behaviour 163, 5 (2026) ; 10.1163/1568539X-bja10355
Pairwise comparisons between Habituation and Control trials showed no significant differences between the two trials for latency to bite (
4. Discussion
Our results suggest that methyl anthranilate (MA) is an immediate, but short-lasting, aversive stimulus for painted turtles. The turtles showed a significant decline in their willingness to consume the MA-treated cod compared to the control cod, supporting the idea that aversion in turtles is displayed by less enthusiastic feeding behaviours (Southwood et al., 2007; Swimmer et al., 2008). Significantly fewer turtles finished the MA-treated cod relative to the baseline trials, and they took significantly longer to engage with (first bite) the MA-treated cod. Those who did finish the MA-treated cod took significantly longer to do so and spat food out more frequently. The response to the MA was generally immediate, observable, and short-lasting. MA is a short-lasting aversive stimulus appearing to last on the order of minutes. The negative behaviours of turning the head, spitting out the food, and wiping the face all suggest agitation and some mild aversive response. However, those behaviours were never observed after the turtles were removed from the apparatus — the turtles continued to feed normally in their home enclosure. The thermal illusion of chemicals that bind to this suite of transient receptor proteins (TRPV1 and TRPA1) seems short-lived, consistent with the effect of MA in birds or capsaicin in mammals. While the turtles as a whole responded negatively when assessed behaviourally, the number of bites/strikes did not significantly increase during treatment, and the latency to bite did not increase after treatment.
In previous studies on birds, the tested individuals responded to MA similarly to the turtles tested here, although we used a lower concentration of MA than typical for birds (Werner & Avery, 2017; Mikiciuk et al., 2021). An aversive response was seen in many species of birds tested with MA (Werner & Avery, 2017), including starlings (Sturnus vulgaris) (Clark et al., 2014b), brown-headed cowbirds (Molothrus ater) (Marples & Roper, 1996), and mallards (Anas platyrhynchos) (DeLiberto & Werner, 2024). The response in these birds was consistent with the spit outs seen in our painted turtles. In addition to an aversive response, tested birds consumed less MA-treated food or water than control food or water (Werner & Avery, 2017; Ahmad et al., 2018), making it clear that birds and painted turtles have similar aversive responses.
While MA did not cause an overall increase in the number of bites and strikes taken at the treated cod, we observed the turtles striking repeatedly at the same few pieces of MA-treated cod without consuming it, although they had consumed the control cod. This means that the turtles consumed less MA-treated cod than control cod, despite the similar number of bites. It is also possible that the turtles lost some interest in the MA-treated cod after experiencing its aversive nature, as birds have done with MA and other aversive substances (Avery et al., 1997; Marples & Roper, 1997). This would explain why the turtles did not take significantly more bites and strikes with MA-treated cod. The turtles may also have detected the MA in the water before consuming the food. MA is aversive even when not directly consumed, but detected through the water, and birds are more sensitive to MA-tainted water than food, so this is possibly also the case for turtles (Crowe-Riddell & Lillywhite, 2023; DeLiberto & Werner, 2024), and would explain the higher proportion of turtles not consuming treated cod, as compared to untreated cod.
The turtles in our study consumed less MA-treated cod than untreated cod, but the pattern was not consistent across all animals, and some of our predictions did not hold, as some turtles responded more strongly to the MA than others. While individual variation is standard in the study of biological systems, it can be difficult to know concretely whether the turtles’ reactions to MA-treated cod was based on their individual states, the MA itself, or, most likely, a combination of both (Waters et al., 2017; Roth et al., 2020; Gharnit et al., 2022). It is possible that the turtles who ate the MA-treated cod more vigorously than others were relatively more motivated to eat (Waters et al., 2017). It is even possible that variation in pain thresholds or perception of MA among turtles could have caused those more strongly impacted by MA to have less desire to eat. Individual perception and pain thresholds of capsaicin have also been shown to vary genetically between mammalian individuals (Szolcsányi, 2015; Nolden et al., 2016), so the same could potentially be true for reptiles and MA. Despite the variation in individual responses among turtles in our study, it is clear that the turtles demonstrated an aversion to MA throughout testing, suggesting that MA is an aversive substance to painted turtles, regardless of individual differences.
While MA causes immediate behavioural effects in painted turtles, verifying its use as a potential aversive stimulus in future studies, it did not seem to cause aversion in the context of this study after a single exposure. Our goal was not to test the learning capacity of the turtles, per se. Turtles were offered MA only once and after many previous exposures to cod. Instead, we were setting up the conditions under which we could identify the clear behavioural response to the MA rather than demonstrating the ability to learn. The post-treatment condition was necessary to demonstrate a strong rebound effect. Future studies might consider using MA with a novel food stimulus to test the nature of the one-trial learning that is common in many avian studies (Marples & Roper, 1997; Ahmad et al., 2016; Chamorro et al., 2023; DeLiberto & Werner, 2024). However, in our context, as cod was not a novel stimulus for the painted turtles we should expect them to need multiple exposures to learn the aversion.
It is possible that we could have seen stronger individual responses with a higher concentration of MA. The turtles’ group-level behavioural responses are similar to those of avian test subjects. However, we used a lower concentration of MA in this study (i.e., 75% v/v with ethanol) based on guidance from our IACUC, as testing the pain response to MA had not been done before (to our knowledge) in this taxon. Had we used a higher concentration, as is common in avian studies (Werner & Avery, 2017; Mikiciuk et al., 2021), we might have seen a more prolonged response. It is also possible that birds and reptiles have differing densities of taste buds, as well as functional variation in TRPA1 receptors, both of which could lead to these taxa perceiving the same substance differently (Rowland et al., 2015; Saito & Saito, 2025). Thus, it is important to further compare and research reptile and bird gustatory systems to compare sensitivities and pain/taste thresholds.
Although we did find MA to be aversive to painted turtles, we did not identify the mechanisms that produced the aversion, as this was beyond the scope of our study. We use inference and suggest that, given the close phylogenetic relationship between turtles and birds (Roth et al., 2019), and the fact that MA had an aversive effect on painted turtles, it seems likely that that TRPA1 receptors of turtles could be sensitive to the same chemicals and produce the same response as in birds (Saito & Tominaga, 2017; Werner & Avery, 2017). However, there may be some other mechanism unique to turtles that is producing the aversion to MA that might better explain the differences between the responses to MA of birds in the literature and the turtles in our study. To better understand how MA impacts painted turtles, the mechanisms behind the effects need to be studied further. Additionally, future studies should consider the breadth of reptilian taxa that respond to MA. MA is an effective aversive stimulus in all bird species that have been tested, despite the amount of diversity within the avian taxa, so many reptilian species could react similarly to MA, despite their diversity (Terčič et al., 2020). However, we have only tested painted turtles, and our study needs to be replicated with other reptile species to determine its generalizability.
Nevertheless, MA affected the painted turtle’s behaviour, suggesting that it could be used as an aversive stimulus in the context of behavioural testing (Werner & Avery, 2017). While this study did not aim to train or teach the turtles in any way, we showed that MA is aversive to turtles and could potentially be used in studies of learning. In learning tasks, an ideal aversive stimulus has significant, immediate effects, cannot be easily avoided by the test subject, and is controllable (Capaldi & Capaldi, 1972). Electric shocks are historically a common aversive stimulus and ideal learning motivator due to their immediate, painful, and directed impact (Brush, 2014; Berezhnoy et al., 2020). While electric shocks do impact reptiles, they are more invasive and less accessible (e.g., the logistics of running electrical shock in aquatic species and cost and maintenance of the equipment to produce the shock) than a food-based motivator, like MA (Lau et al., 2025). MA’s observable impact makes the association between the task and the aversive stimulus (MA) clear and simple for reptiles to learn (Yau & McNally, 2019). This may speed up the learning and make it more effective relative to other, less salient (to reptiles) aversive stimuli, such as temperature or withholding food.
Having a more effective aversive stimulus, like MA, also makes it more likely for researchers to learn about the test species’ cognitive abilities, as they would be less limited by an association that is difficult to learn. Using MA would allow for the use of aversive stimuli, not just positive stimuli (Szabo et al., 2021) and set up food avoidance/preference scenarios (Clark et al., 2014a; Skotnikova & Zhelankin, 2018). MA as an aversive stimulus even has potential value for use in reptiles in a conservation setting, potentially setting up conditioned taste aversions to help reptiles avoid unsafe situations (Andrewartha et al., 2023). Thus, the possibilities for using MA in a testing setting as an aversive stimulus are vast and variable, and it could help us test reptiles’ cognitive abilities more efficiently, without being limited by ineffective aversive stimuli.
In conclusion, this study was an initial exploration of reptiles responding to MA in a way consistent with having a negative reaction. We propose that MA could potentially be used as an aversive stimulus for reptiles in behavioural and cognitive tasks. Further investigations into the mechanisms behind the reaction to MA, the generalizability of our results to all reptiles, and a more detailed comparison of bird versus reptilian taste mechanisms need to be done to fine-tune and further our results. MA could allow researchers to study and test the behaviour, learning, and cognition of reptiles similarly to how mammals and birds are tested, while keeping the species within their natural gestalts, allowing for more effective testing, higher validity, and greater comparison between taxa. Cognitive test results in reptiles could then be made more definitive, allowing for the clarification of reptilian capacities to perform certain complex cognitive behaviours, and more effective comparisons between taxa can shed additional light on the evolution of cognition, specifically within reptiles.
Corresponding author’s e-mail address: theresagalban52@gmail.com
Acknowledgements
Many thanks to the National Science Foundation, which partially funded this project (Grant NSF-1754297 (TCR)), as well as the Franklin & Marshall College Vivarium and Psychology Department. This study was conducted with permission from the Maryland Department of Natural Resources (SCO No. 51936) and Franklin & Marshall College animal ethics committee (IACUC No. R_73neuzUgQmK0BMJ). We would like to thank Lillian Basom, Richelle Wagner, and all of the volunteers and caretakers in The Vivarium for assisting in the painted turtle husbandry and day-to-day care. Finally, we want to thank Shoshana Frank for her advice and motivation, as well as her endless love and support.
References
Ahmad, S., Saleem, Z., Jabeen, F., Hussain, B., Sultana, T., Sultana, S., Al-Ghanim, K.A., Al-Mulhim, N.M.A. & Mahboob, S. (2018). Potential of natural repellents methylanthranilate and anthraquinone applied on maize seeds and seedlings against house sparrow (Passer domesticus) in captivity. — Braz. J. Biol. 78: 667-672.
Ahmad, S., Saleem, Z., Jabeen, F., Khan, H.A., Yaqub, S., Samiullah, K., Mubarik, S., Irfan, M. & Shaheen, T. (2016). Effectiveness of methylanthranilate and anthraquinone as repellent against house crows (Corvus splendens) on wheat seeds and seedlings in captivity. — Pak. J. Zool. 48: 1591-1594.
Alarcón, D.E. (2022). Motivational state. — In: Encyclopedia of animal cognition and behavior (Shackelford, T. & Vonk, J., eds). Springer, Cham, p. 4437-4443.
Andrewartha, T., Evans, M., Blencowe, A., Brewer, K., Gordon, I.J. & Manning, A.D. (2023). Landscapes of nausea: successful conditioned taste aversion in a wild red fox population. — Conserv. Sci. Pract. 5: e12984.
Anselme, P. & Güntürkün, O. (2019). How foraging works: uncertainty magnifies food-seeking motivation. — Behav. Brain Sci. 42: e35.
Avery, M.L., Humphrey, J.S. & Decker, D.G. (1997). Feeding deterrence of anthraquinone, anthracene, and anthrone to rice-eating birds. — J. Wildl. Manage.: 1359-1365.
Berezhnoy, D.S., Zamorina, T.A. & Inozemtsev, A.N. (2020). Appetitive conditioning task in a shuttle box and its comparison with the active avoidance paradigm. — Learn. Behav. 48: 364-372.
Bernich, A., Maute, K., Contador-Kelsall, I.C., Story, P.G., Hose, G.C. & French, K. (2022). Space use and daily movement patterns in an arid zone agamid lizard. — Wildl. Res. 49: 557-570.
Bodensteiner, B.L., Agudelo-Cantero, G.A., Arietta, A.A., Gunderson, A.R., Muñoz, M.M., Refsnider, J.M. & Gangloff, E.J. (2021). Thermal adaptation revisited: how conserved are thermal traits of reptiles and amphibians? — J. Exp. Zool. Part A: Ecol. Integr. Physiol. 335: 173-194.
Brejcha, J., Bataller, J.V., Bosáková, Z., Geryk, J., Havlíková, M., Kleisner, K., Maršík, P. & Font, E. (2019). Body coloration and mechanisms of colour production in Archelosauria: the case of deirocheline turtles. — Roy. Soc. Open Sci. 6: 190319.
Brush, F.R. (ed.) (2014). Aversive conditioning and learning. — Academic Press, San Diego, CA.
Burghardt, G.M. (1977). Learning processes in reptiles. — Herpetologica 2: 175-196.
Burghardt, G.M. (2020). Box B9: The learning repertoire of reptiles. — In: Zoo animal learning and training (Dorey, N., Melfi, V. & Ward, S., eds). Wiley, Hoboken, NJ, p. 227-230.
Capaldi, E.J. & Capaldi, E.D. (1972). Aversive learning situations: apparatus and procedures. — In: Methods in psychobiology (Myers, R.D., ed.). Academic Press, San Diego, CA, p. 59-81.
Chamorro, S., Cimiano Fernández, T., Arija, I., Viveros, A. & Pascual-Teresa, S.D. (2023). Development of an animal model for testing the effect of diet on memory (poster presentation). — In: 4th International Food Bioactives & Health Conference, Prague, Czech Republic.
Clark, B.F., Amiel, J.J., Shine, R., Noble, D.W. & Whiting, M.J. (2014b). Colour discrimination and associative learning in hatchling lizards incubated at ‘hot’and ‘cold’temperatures. — Behav. Ecol. Sociobiol. 68: 239-247.
Clark, L., Bryant, B. & Mezine, I. (2000). Bird aversive properties of methyl anthranilate, yucca, xanthoxylum, and their mixtures. — J. Chem. Ecol. 26: 1219-1234.
Clark, L., Hagelin, J. & Werner, S. (2014b). The chemical senses in birds. — In: Sturkie’s avian physiology, 6th edn. (Scanes, C., ed.). Academic Press, San Diego, CA, p. 89-111.
Crowe-Riddell, J.M. & Lillywhite, H.B. (2023). Sensory systems. — In: Health and welfare of captive reptiles (Arena, P., Burghardt, G. & Warwick, C., eds). Springer, Cham, p. 45-91.
Davis, K.M. & Burghardt, G.M. (2011). Turtles (Pseudemys nelsoni) learn about visual cues indicating food from experienced turtles. — J. Comp. Psychol. 125: 404.
Dayan, P. & Balleine, B.W. (2002). Reward, motivation, and reinforcement learning. — Neuron 36: 285-298.
De Meester, G. & Baeckens, S. (2021). Reinstating reptiles: from clueless creatures to esteemed models of cognitive biology. — Behaviour 158: 1057-1076.
DeLiberto, S.T. & Werner, S.J. (2024). Applications of chemical bird repellents for crop and resource protection: a review and synthesis. — Wildl. Res. 51: WR23062.
DeNardo, D.F. (2021). Behavioral biology of reptiles. — In: Behavioral biology of laboratory animals (Coleman, K. & Shapiro, S., eds). CRC Press, Boca Raton, FL, p. 361-375.
Fischer, M.J., Ciotu, C.I. & Szallasi, A. (2020). The mysteries of capsaicin-sensitive afferents. — Front. Physiol. 11: 554195.
Fischer, E.K., Nowicki, J.P. & O’Connell, L.A. (2019). Evolution of affiliation: pattern of convergence from genomes to behaviour. — Philos. Trans. Roy. Soc. Lond. B.: Biol Sci. 374: 20180242.
Gharnit, E., Dammhahn, M., Garant, D. & Réale, D. (2022). Resource availability, sex, and individual differences in exploration drive individual diet specialization. — Am. Nat. 200: 1-16.
Giacometti, D., Yagi, K.T., Abney, C.R., Jung, M.P. & Tattersall, G.J. (2021). Staying warm is not always the norm: behavioural differences in thermoregulation of two snake species. — Can. J. Zool. 99: 974-983.
Griffin, D.K., Larkin, D.M., O’Connor, R.E. & Romanov, M.N. (2022). Dinosaurs: comparative cytogenomics of their reptile cousins and avian descendants. — Animals 13: 106.
Huey, R.B., Garland Jr, T. & Turelli, M. (2019). Revisiting a key innovation in evolutionary biology: Felsenstein’s “phylogenies and the comparative method”. — Am. Nat. 193: 755-772.
Kaur, R., Singla, N., Bansal, N. & Pathak, D. (2016). Development of an integrated rodent pest management approach using capsicum oleoresin as an aversive agent against rodents. — Nat. Res. Manage.: Ecol. Perspect. 13: 835-836.
Krishnatreyya, H., Hazarika, H., Saha, A. & Chattopadhyay, P. (2018). Capsaicin, the primary constituent of pepper sprays and its pharmacological effects on mammalian ocular tissues. — Eur. J. Pharmacol. 819: 114-121.
Krochmal, A.R. & Bakken, G.S. (2003). Thermoregulation is the pits: use of thermal radiation for retreat site selection by rattlesnakes. — J. Exp. Biol. 206: 2539-2545.
Krochmal, A.R., Place, A.J., LaDuc, T.J. & Roth II, T.C. (2018). Phylogenetic patterns in learning and decision making in pit vipers (Viperidae: Crotalinae). — Anim. Behav. 145: 117-123.
Kuczaj, S.A. & Xitco Jr, M.J. (2002). It takes more than fish: the psychology of marine mammal training. — Int. J. Comp. Psychol. 15: 186-200.
Kundey, S.M. (2024). Advancing our understanding of cognition by including amphibians and reptiles in comparative cognition research. — Comp. Cogn. Behav. Rev. 19: 29-32.
Lau, C., Hu, C., Koprowski, J., Lee, Z., Narula, A., Sankaran, S., Welk, J. & Powers, A.S. (2025). Classical fear conditioning in turtles (Chrysemys picta). — Int. J. Comp. Psychol. 38: ijcp.39691.
Legendre, L.J. & Davesne, D. (2020). The evolution of mechanisms involved in vertebrate endothermy. — Philos. Trans. Roy. Soc. Lond. B: Biol. Sci. 375: 20190136.
López, J.C., Rodríguez, F., Gómez, Y., Vargas, J.P., Broglio, C. & Salas, C. (2000). Place and cue learning in turtles. — Anim. Learn. Behav. 28: 360-372.
López, J.C., Vargas, J.P., Gómez, Y. & Salas, C. (2003). Spatial and non-spatial learning in turtles: the role of medial cortex. — Behav. Brain Res. 143: 109-120.
Lunceford, B.E. & Kubanek, J. (2015). Reception of aversive taste. — Integr. Compar. Biol. 55: 507-517.
Luo, Z.W., Cho, J.S. & Lee, S.Y. (2019). Microbial production of methyl anthranilate, a grape flavor compound. — Proc. Natl. Acad. Sci. USA 116: 10749-10756.
Marples, N.M. & Roper, T.J. (1997). Response of domestic chicks to methyl anthranilate odour. — Anim. Behav. 53: 1263-1270.
Matsubara, S., Deeming, D.C. & Wilkinson, A. (2017). Cold-blooded cognition: new directions in reptile cognition. — Curr. Opin. Behav. Sci. 16: 126-130.
McNab, B.K. (2019). What determines the basal rate of metabolism? — J. Exp. Biol. 222: jeb205591.
Mednikova, Y.S. & Kalabushev, S.N. (2023). Thermally dependent behaviour of cold-blooded animals: overcoming two temperature barriers on the way to warm-blooded. — J. Behav. Brain Sci. 13: 95-112.
Michael, J. (1975). Positive and negative reinforcement, a distinction that is no longer necessary; or a better way to talk about bad things. — Behaviorism 3: 33-44.
Mikiciuk, G., Chełpiński, P., Mikiciuk, M., Możdżer, E. & Telesiński, A. (2021). The effect of methyl anthranilate-based repellent on chemical composition and selected physiological parameters of sweet cherry (Prunus avium L.). — Agronomy 11: 256.
Miles, D.B. & Dunham, A.E. (1993). Historical perspectives in ecology and evolutionary biology: the use of phylogenetic comparative analyses. — Annu. Rev. Ecol. Syst. 24: 587-619.
Moldowan, P.D., Keevil, M.G., Mills, P.B., Brooks, R.J. & Litzgus, J.D. (2015). Diet and feeding behaviour of snapping turtles (Chelydra serpentina) and midland painted turtles (Chrysemys picta marginata) in Algonquin Provincial Park, Ontario. — Can. Field-Nat. 129: 403-408.
Mota-Rojas, D., Titto, C.G., Orihuela, A., Martínez-Burnes, J., Gómez-Prado, J., Torres-Bernal, F., Flores-Padilla, K., Carvajal-de la Fuente, V. & Wang, D. (2021). Physiological and behavioral mechanisms of thermoregulation in mammals. — Animals 11: 1733.
Nolden, A.A., McGeary, J.E. & Hayes, J.E. (2016). Differential bitterness in capsaicin, piperine, and ethanol associates with polymorphisms in multiple bitter taste receptor genes. — Physiol. Behav. 156: 117-127.
Padmala, S. & Pessoa, L. (2010). Interactions between cognition and motivation during response inhibition. — Neuropsychology 48: 558-565.
Pettersen, A.K. (2020). Countergradient variation in reptiles: thermal sensitivity of developmental and metabolic rates across locally adapted populations. — Front. Physiol. 11: 547.
Powers, A.S., Hogue, P., Lynch, C., Gattuso, B., Lissek, S. & Nayal, C. (2009). Role of acetylcholine in negative patterning in turtles (Chrysemys picta). — Behav. Neurosci. 123: 804.
Prassack, S.L., Bagatto, B. & Henry, R.P. (2001). Effects of temperature and aquatic PO2 on the physiology and behaviour of Apalone ferox and Chrysemys picta. — J. Exp. Biol. 204: 2185-2195.
Roth, A.D., Krochmal, A.R. & Roth, T.C. (2021). Context-specific cue use in the eastern painted turtle (Chrysemys picta) and its effects on decision making. — Behaviour 158: 1101-1120.
Roth, T.C. & Krochmal, A.R. (2016). Pharmacological evidence is consistent with a prominent role of spatial memory in complex navigation. — Proc. Roy Soc. Lond. B: Biol. Sci. 283: 20152548.
Roth, T.C. & Krochmal, A.R. (2024). Reptilian cognition. — Curr. Biol. 34: R129-R130.
Roth, T.C., Krochmal, A.R. & LaDage, L.D. (2019). Reptilian cognition: a more complex picture via integration of neurological mechanisms, behavioral constraints, and evolutionary context. — BioEssays 41: 1900033.
Roth, T.C., Rosier, M., Krochmal, A.R. & Clark, L. (2020). A multi-trait, field-based examination of personality in a semi-aquatic turtle. — Ethology 126: 851-857.
Rowland, H.M., Rockwell Parker, M., Jiang, P., Reed, D.R. & Beauchamp, G.K. (2015). Comparative taste biology with special focus on birds and reptiles. — In: Handbook of olfaction and gustation (Doty, R.L., ed.). Wiley, Hoboken, NJ, p. 957-982.
Saito, S. & Saito, C.T. (2025). Evolution of temperature receptors and their roles in sensory diversification and adaptation. — Zool. Sci. 42: zs240060.
Saito, S. & Tominaga, M. (2017). Evolutionary tuning of TRPA1 and TRPV1 thermal and chemical sensitivity in vertebrates. — Temperature 4: 141-152.
Saito, S., Nakatsuka, K., Takahashi, K., Fukuta, N., Imagawa, T., Ohta, T. & Tominaga, M. (2012). Analysis of transient receptor potential ankyrin 1 (TRPA1) in frogs and lizards illuminates both nociceptive heat and chemical sensitivities and coexpression with TRP vanilloid 1 (TRPV1) in ancestral vertebrates. — J. Biol. Chem. 287: 30743-30754.
Shanbhag, B., Ammanna, V. & Saidapur, S. (2010). Associative learning in hatchlings of the lizard Calotes versicolor: taste and colour discrimination. — Amphibia-Reptilia 31: 475-481.
Shaw, R.C. & Schmelz, M. (2017). Cognitive test batteries in animal cognition research: evaluating the past, present and future of comparative psychometrics. — Anim. Cogn. 20: 1003-1018.
Shuba, Y.M. (2021). Beyond neuronal heat sensing: diversity of TRPV1 heat-capsaicin receptor-channel functions. — Front. Cell. Neurol. 14: 612480.
Skotnikova, I.G. & Zhelankin, R.V. (2018). Reptiles’ decision making strategies in a choice between response alternatives (color discrimination task) and possible similarities with human behavior. — Fechner Day 148.
Southwood, A., Fritsches, K., Brill, R. & Swimmer, Y. (2008). Sound, chemical, and light detection in sea turtles and pelagic fishes: sensory-based approaches to bycatch reduction in longline fisheries. — Endanger. Species Res. 5: 225-238.
Swimmer, Y., McNaughton, L., Southwood, A. & Brill, R. (2007). Tests of repellent bait to reduce turtle bycatch in longline fisheries. — Bycatch Community Network Newsletter 5: 18-20.
Szabo, B., Noble, D.W. & Whiting, M.J. (2021). Learning in non-avian reptiles 40 years on: advances and promising new directions. — Biol. Rev. 96: 331-356.
Szabo, B., Valencia-Aguilar, A., Damas-Moreira, I. & Ringler, E. (2022). Wild cognition–linking form and function of cognitive abilities within a natural context. — Curr. Opin. Behav. Sci. 44: 101115.
Szolcsányi, J. (2014). Capsaicin and sensory neurons: a historical perspective. — In: Capsaicin as a therapeutic molecule (Abdel-Salam, O., ed.). Springer, Cham, p. 1-37.
Talavera, K., Startek, J.B., Alvarez-Collazo, J., Boonen, B., Alpizar, Y.A., Sanchez, A., Naert, R. & Nilius, B. (2020). Mammalian transient receptor potential TRPA1 channels: from structure to disease. — Physiol. Rev. 100: 725-803.
Tattersall, G.J. (2016). Reptile thermogenesis and the origins of endothermy. — Zoology 119: 403-405.
Terčič, D., Pančur, M., Jordan, D. & Zupan Šemrov, M. (2020). Effects of dimethyl anthranilate-based repellents on behavior, plumage condition, egg quality, and performance in laying hens. — Front. Vet. Sci. 7: 533.
Toates, F. (1995). Animal motivation and cognition. — In: Comparative approaches to cognitive science (Meyer, J. & Roitblat, H., eds). Bradford Books, Cambridge, MA, p. 435-464.
Tomonaga, M., Haraguchi, D. & Wilkinson, A. (2023). Slowly walking down to the more food: relative quantity discrimination in African spurred tortoises (Centrochelys sulcata). — Anim. Cogn. 26: 1675-1683.
Valenzuela, N. (2009). The painted turtle, Chrysemys picta: a model system for vertebrate evolution, ecology, and human health. — Cold Spring Harbor Protocols 7: pdb-emo124.
van der Schaaf, M.E., Schmidt, K., Kaur, J., Gamer, M., Wiech, K., Forkmann, K. & Bingel, U. (2022). Acquisition learning is stronger for aversive than appetitive events. — Commun. Biol. 5: 302.
van Horik, J.O. & Madden, J.R. (2016). A problem with problem solving: motivational traits, but not cognition, predict success on novel operant foraging tasks. — Anim. Behav. 114: 189-198.
Varholick, J.A. (2022). Umwelt. — In: Encyclopedia of animal cognition and behavior (Shackelford, T. & Vonk, J., eds). Springer, Cham, p. 7116-7118.
Waters, R.M., Bowers, B.B. & Burghardt, G.M. (2017). Personality and individuality in reptile behavior. — In: Personality in nonhuman animals (Kuczaj, S., Vonk, J. & Weiss, A., eds). Springer, Cham, p. 153-184.
Wergård, E.M., Temrin, H., Forkman, B., Spångberg, M., Fredlund, H. & Westlund, K. (2015). Training pair-housed rhesus macaques (Macaca mulatta) using a combination of negative and positive reinforcement. — Behav. Process. 113: 51-59.
Werner, S.J. & Avery, M.L. (2017). Chemical repellents. — In: Ecology and management of blackbirds (Icteridae) in North America (Avery, M., Dolbeer, R. & Linz, G., eds). CRC Press, Boca Raton, FL, p. 135-158.
Wilkinson, A. & Glass, E. (2018). Tortoises–cold-blooded cognition: how to get a tortoise out of its shell. — In: Field and laboratory methods in animal cognition: a comparative guide (Amici, F. & Bueno-Guerra, N., eds). Cambridge University Press, Cambridge, p. 401.
Yau, J.O.Y. & McNally, G.P. (2019). Rules for aversive learning and decision-making. — Curr. Opin. Behav. Sci. 26: 1-8.
Yee, D.M. & Braver, T.S. (2018). Interactions of motivation and cognitive control. — Curr. Opin. Behav. Sci. 19: 83-90.
Zhang, H., Wang, C., Zhang, K., Kamau, P.M., Luo, A., Tian, L. & Lai, R. (2022). The role of TRPA1 channels in thermosensation. — Cell Insight 1: 100059.
Zhong, H., Shang, S., Wu, X., Chen, J., Zhu, W., Yan, J., Li, H. & Zhang, H. (2017). Genomic evidence of bitter taste in snakes and phylogenetic analysis of bitter taste receptor genes in reptiles. — PeerJ. 5: e3708.
Zhong, H., Shang, S., Zhang, H., Chen, J., Wu, X. & Zhang, H. (2019). Characterization and phylogeny of bitter taste receptor genes (Tas2r) in squamata. — Genetica 147: 131-139.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}